
Conservation of Imprinting and Methylation of MKRN3, MAGEL2 and NDN Genes in Cattle

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Tissue and Placental Sample Collection
2.2. DNA Extraction and PCR Amplification for Identification of SNPs
2.3. Allelic Expression Analysis by RT-PCR
2.4. DNA Methylation Analysis by Bisulfite Sequencing PCR (BSP)
3. Results
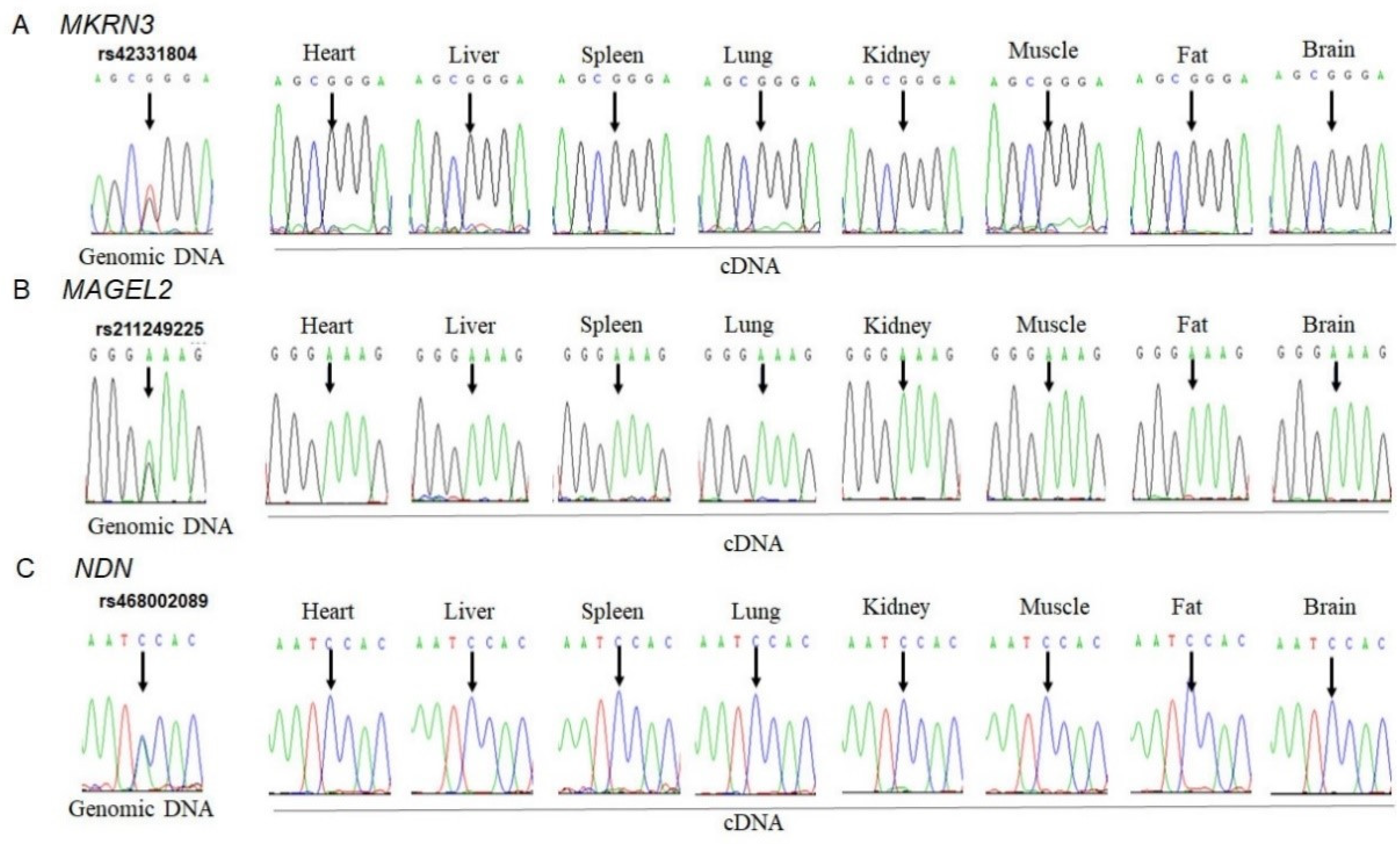
3.1. Monoallelic Expression of MKRN3, MAGEL2 and NDN in Bovine Tissues
3.2. Paternally Expressed MKRN3, MAGEL2 and NDN in Bovine Placenta
3.3. Identification of Bovine PWS-IC, MKRN3 and NDN DMR
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AT | annealing temperature |
| DMRs | differentially methylated regions |
| SNP | single-nucleotide polymorphism |
| 5’UTR | 5’ untranslated region |
| NDN | necdin |
| MAGEL2 | melanoma antigen-like gene 2 |
| MKRN3 | makorin ring finger protein 3 |
| PWS | Prader-Willi syndrome |
| AS | Angelman syndrome |
| snoRNA | small nucleolar RNA |
| ICRs | imprinting control regions |
| gDMRs | germline DMRs |
| sDMRs | secondary DMRs |
| PWS-IC | PWS imprinting center |
| CGIs | CpG islands |
| GTEx | Genotype-Tissue Expression |
References
- Edwards, C.A.; Ferguson-Smith, A.C. Mechanisms regulating imprinted genes in clusters. Curr. Opin. Cell. Biol. 2007, 19, 281–289. [Google Scholar] [CrossRef]
- Soellner, L.; Begemann, M.; Mackay, D.J.; Grønskov, K.; Tümer, Z.; Maher, E.R.; Temple, I.K.; Monk, D.; Riccio, A.; Linglart, A.; et al. Recent advances in imprinting disorders. Clin. Genet. 2017, 91, 3–13. [Google Scholar] [CrossRef] [Green Version]
- Uribe-Lewis, S.; Woodfine, K.; Stojic, L.; Murrell, A. Molecular mechanisms of genomic imprinting and clinical implications for cancer. Expert. Rev. Mol. Med. 2011, 13, e2. [Google Scholar] [CrossRef]
- Nicholls, R.D.; Saitoh, S.; Horsthemke, B. Imprinting in Prader-Willi and Angelman syndromes. Trends Genet. 1998, 14, 194–200. [Google Scholar] [CrossRef]
- Cassidy, S.B.; Schwartz, S.; Miller, J.L.; Driscoll, D.J. Prader-Willi syndrome. Genet. Med. 2012, 14, 10–26. [Google Scholar] [CrossRef] [Green Version]
- Chamberlain, S.J. RNAs of the human chromosome 15q11-q13 imprinted region. Wiley Interdiscip. Rev. RNA 2013, 4, 155–166. [Google Scholar] [CrossRef]
- Rougeulle, C.; Glatt, H.; Lalande, M. The Angelman syndrome candidate gene, UBE3A/E6-AP, is imprinted in brain. Nat. Genet. 1997, 17, 14–15. [Google Scholar] [CrossRef]
- Barlow, D.P.; Bartolomei, M.S. Genomic imprinting in mammals. Cold Spring Harb. Perspect. Biol. 2014, 6, a018382. [Google Scholar] [CrossRef] [Green Version]
- Pólvora-Brandão, D.; Joaquim, M.; Godinho, I.; Aprile, D.; Álvaro, A.R.; Onofre, I.; Raposo, A.C.; de Almeida, L.P.; Duarte, S.T.; da Rocha, S.T. Loss of hierarchical imprinting regulation at the Prader-Willi/Angelman syndrome locus in human iPSCs. Hum. Mol. Genet. 2018, 27, 3999–4011. [Google Scholar] [CrossRef]
- Xie, W.; Barr, C.L.; Kim, A.; Yue, F.; Lee, A.Y.; Eubanks, J.; Dempster, E.L.; Ren, B. Base-resolution analyses of sequence and parent-of-origin dependent DNA methylation in the mouse genome. Cell 2012, 148, 816–831. [Google Scholar] [CrossRef] [Green Version]
- Smith, E.Y.; Futtner, C.R.; Chamberlain, S.J.; Johnstone, K.A.; Resnick, J.L. Transcription is required to establish maternal imprinting at the Prader–Willi syndrome and Angelman syndrome locus. PLoS Genet. 2011, 7, e1002422. [Google Scholar] [CrossRef] [Green Version]
- Jay, P.; Rougeulle, C.; Massacrier, A.; Moncla, A.; Mattei, M.G.; Malzac, P.; Roëckel, N.; Taviaux, S.; Lefranc, J.L.; Cau, P.; et al. The human necdin gene, NDN, is maternally imprintedand located in the Prader-Willi syndrome chromosomal region. Nat. Genet. 1997, 17, 357–361. [Google Scholar] [CrossRef] [PubMed]
- Jong, M.T.C.; Gray, T.A.; Ji, Y.; Glenn, C.C.; Saitoh, S.; Driscoll, D.J.; Nicholls, R.D. A novel imprinted gene, encoding a RING zinc-finger protein, and overlapping antisense transcript in the Prader-Willi syndrome critical region. Hum. Mol. Genet. 1999, 8, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Bielinska, B.; Blaydes, S.M.; Buiting, K.; Yang, T.; Krajewska-Walasek, M.; Horsthemke, B.; Brannan, C.I. De novo deletions of SNRPN exon 1 in early human and mouse embryos result in a paternal to maternal imprint switch. Nat. Genet. 2000, 25, 74–78. [Google Scholar] [CrossRef]
- Ohta, T.; Gray, T.; Rogan, P.; Buiting, K.; Gabriel, J.; Saitoh, S.; Muralidhar, B.; Bilienska, B.; Krajewska-Walasek, M.; Driscoll, D.; et al. Imprinting-mutation mechanisms in Prader–Willi syndrome. Am. J. Hum. Genet. 1999, 64, 397–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Donato, M.; Hussain, T.; Rodulfo, H.; Peters, S.O.; Imumorin, I.G.; Thomas, B.N. Conservation of Repeats at the Mammalian KCNQ1OT1-CDKN1C Region Suggests a Role in Genomic Imprinting. Evol. Bioinform. Online 2017, 13, 1176934317715238. [Google Scholar] [CrossRef] [PubMed]
- Surani, M.A.; Barton, S.C.; Norris, M.L. Development of reconstituted mouse eggs suggests imprinting of the genome during gametogenesis. Nature 1984, 308, 548–550. [Google Scholar] [CrossRef]
- Barlow, D.P. Gametic imprinting in mammals. Science 1995, 270, 1610–1613. [Google Scholar] [CrossRef]
- Kuehn, C.; Edel, C.; Weikard, R.; Thaller, G. Dominance and parent-of-origin effects of coding and non-coding alleles at the acylCoA-diacylglycerol-acyltransferase (DGAT1) gene on milk production traits in German Holstein cows. BMC Genet. 2007, 8, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pausch, H.; Flisikowski, K.; Jung, S.; Emmerling, R.; Edel, C.; Götz, K.U.; Fries, R. Genome-wide association study identifies two major loci affecting calving ease and growth-related traits in cattle. Genetics 2011, 187, 289–297. [Google Scholar] [CrossRef] [Green Version]
- Van Laere, A.-S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.; Buys, N.; Tally, M.; et al. A regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [CrossRef] [PubMed]
- Boysen, T.J.; Tetens, J.; Thaller, G. Detection of a quantitative trait locus for ham weight with polar over dominance near the ortholog of the callipyge locus in an experimental pig F2 population. J. Anim. Sci. 2010, 88, 3167–3172. [Google Scholar] [CrossRef] [Green Version]
- Cockett, N.E.; Jackson, S.P.; Shay, T.L.; Farnir, F.; Berghmans, S.; Snowder, G.D.; Nielsen, D.M.; Georges, M. Polar over dominance at the ovine callipyge locus. Science 1996, 273, 236–238. [Google Scholar] [CrossRef] [PubMed]
- Matika, O.; Sechi, S.; Pong-Wong, R.; Houston, R.D.; Clop, A.; Woolliams, J.A.; Bishop, S.C. Characterization of OAR1 and OAR18 QTL associated with muscle depth in British commercial terminal sire sheep. Anim. Genet. 2010, 42, 172–180. [Google Scholar] [CrossRef]
- Hansen, P.J. Current and future assisted reproductive technologies for mammalian farm animals. In Current and Future Reproductive Technologies and World Food Production; Cliff lamb, G., DiLorenzo, N., Eds.; Advances in Experimental Medicine and Biology; Springer: New York, NY, USA, 2014; Volume 752, pp. 1–22. [Google Scholar]
- Bourneuf, E.; Otz, P.; Pausch, H.; Jagannathan, V.; Michot, P.; Grohs, C.; Piton, G.; Ammermüller, S.; Deloche, M.C.; Fritz, S.; et al. Rapid Discovery of De Novo Deleterious Mutations in Cattle Enhances the Value of Livestock as Model Species. Sci. Rep. 2017, 7, 11466. [Google Scholar] [CrossRef] [Green Version]
- Zaitoun, I.; Khatib, H. Comparative genomic imprinting and expression analysis of six cattle genes. J. Anim. Sci. 2008, 86, 25–32. [Google Scholar] [CrossRef]
- Zaitoun, I.; Khatib, H. Assessment of genomic imprinting of SLC38A4, NNAT, NAP1L5, and H19 in cattle. BMC Genet. 2006, 7, 49. [Google Scholar] [CrossRef] [Green Version]
- Kaneda, M.; Takahashi, M.; Yamanaka, K.I.; Saito, K.; Taniguchi, M.; Akagi, S.; Watanabe, S.; Nagai, T. Epigenetic analysis of bovine parthenogenetic embryonic fibroblasts. J. Reprod. Dev. 2017, 63, 365–375. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, X.; Clark, A.G. Using next-generation RNA sequencing to identify imprinted genes. Heredity 2014, 113, 156–166. [Google Scholar] [CrossRef] [Green Version]
- Babak, T.; Deveale, B.; Armour, C.; Raymond, C.; Cleary, M.A.; van der Kooy, D.; Johnson, J.M.; Lim, L.P. Global survey of genomic imprinting by transcriptome sequencing. Curr. Biol. 2008, 18, 1735–1741. [Google Scholar] [CrossRef] [Green Version]
- DeVeale, B.; van der Kooy, D.; Babak, T. Critical evaluation of imprinted gene expression by RNA-Seq: A new perspective. PLoS Genet. 2012, 8, e1002600. [Google Scholar] [CrossRef]
- Baran, Y.; Subramaniam, M.; Biton, A.; Tukiainen, T.; Tsang, E.K.; Rivas, M.A.; Pirinen, M.; Gutierrez-Arcelus, M.; Smith, K.S.; Kukurba, K.R.; et al. The landscape of genomic imprinting across diverse adult human tissues. Genome Res. 2015, 25, 927–936. [Google Scholar] [CrossRef] [Green Version]
- Chen, Z.; Hagen, D.E.; Wang, J.; Elsik, C.G.; Ji, T.; Siqueira, L.G.; Hansen, P.J.; Rivera, R.M. Global assessment of imprinted gene expression in the bovine conceptus by next generation sequencing. Epigenetics 2016, 11, 501–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Imumorin, I.G.; Kim, E.H.; Lee, Y.M.; De Koning, D.J.; van Arendonk, J.A.; De Donato, M.; Taylor, J.F.; Kim, J.J. Genome Scan for Parent-of-Origin QTL Effects on Bovine Growth and Carcass Traits. Front. Genet. 2011, 2, 44. [Google Scholar] [CrossRef] [Green Version]
- Boccaccio, I.; Glatt-Deeley, H.; Watrin, F.; Roeckel, N.; Lalande, M.; Muscatelli, F. The human MAGEL2 gene and its mouse homologue are paternally expressed and mapped to the Prader-Willi region. Hum. Mol. Genet. 1999, 8, 2497–2505. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sutcliffe, J.S.; Han, M.; Christian, S.L.; Ledbetter, D.H. Neuronally-expressed necdin gene: An imprinted candidate gene in Prader-Willi syndrome. Lancet 1997, 350, 1520–1521. [Google Scholar] [CrossRef]
- Abreu, A.P.; Dauber, A.; Macedo, D.B.; Noel, S.D.; Brito, V.N.; Gill, J.C.; Cukier, P.; Thompson, I.R.; Navarro, V.M.; Gagliardi, P.C.; et al. Central precocious puberty caused by mutations in the imprinted gene MKRN3. N. Engl. J. Med. 2013, 368, 2467–2475. [Google Scholar] [CrossRef] [Green Version]
- Abreu, A.P.; Macedo, D.B.; Brito, V.N.; Kaiser, U.B.; Latronico, A.C. A new pathway in the control of the initiation of puberty: The MKRN3 gene. J. Mol. Endocrinol. 2015, 54, R131–R139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simsek, E.; Demiral, M.; Ceylaner, S.; Kırel, B. Two Frameshift Mutations in MKRN3 in Turkish Patients with Familial Central Precocious Puberty. Horm. Res. Paediatr. 2017, 87, 405–411. [Google Scholar] [CrossRef] [PubMed]
- Fanis, P.; Skordis, N.; Toumba, M.; Papaioannou, N.; Makris, A.; Kyriakou, A.; Neocleous, V.; Phylactou, L.A. Central Precocious Puberty Caused by Novel Mutations in the Promoter and 5′-UTR Region of the Imprinted MKRN3 Gene. Front. Endocrinol. 2019, 10, 677. [Google Scholar] [CrossRef]
- Macedo, D.B.; França, M.M.; Montenegro, L.R.; Cunha-Silva, M.; Bessa, D.S.; Abreu, A.P.; Kaiser, U.B.; Mendonca, B.; Jorge, A.A.; Brito, V.N.; et al. Central Precocious Puberty Caused by a Heterozygous Deletion in the MKRN3 Promoter Region. Neuroendocrinology 2018, 107, 127–132. [Google Scholar] [CrossRef]
- Lee, S.; Kozlov, S.; Hernandez, L.; Chamberlain, S.J.; Brannan, C.I.; Stewart, C.L.; Waverick, R. Expression and imprinting of MAGEL2 suggesta role in Prader-Willi syndrome and the homologous murine imprinting phenotype. Hum. Mol. Genet. 2000, 9, 1813–1819. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MacDonald, H.R.; Wevrick, R. The necdin gene is deleted in Prader-Willi syndrome and is imprinted in human and mouse. Hum. Mol. Genet. 1997, 6, 1873–1878. [Google Scholar] [CrossRef] [Green Version]
- Kanber, D.; Giltay, J.; Wieczorek, D.; Zogel, C.; Hochstenbach, R.; Caliebe, A.; Kuechler, A.; Horsthemke, B.; Buiting, K. A paternal deletion of MKRN3, MAGEL2 and NDN does not result in Prader–Willi syndrome. Eur. J. Hum. Genet. 2009, 17, 582–590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peters, S.O.; De Donato, M.; Hussain, T.; Rodulfo, H.; Babar, M.E.; Imumorin, I.G. Sequence variation of necdin gene in Bovidae. J. Anim. Sci. Technol. 2018, 60, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Watrin, F.; Roëckel, N.; Lacroix, L.; Mignon, C.; Mattei, M.-G.; Disteche, C.; Muscatelli, F. The mouse Necdin gene is expressed from the paternal allele only and lies in the 7C region of the mouse chromosome 7, a region of conserved synteny to the human Prader–Willi syndrome region. Eur. J. Hum. Genet. 1997, 5, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Hershko, A.; Razin, A.; Shemer, R. Imprinted methylation and its effect on expression of the mouse Zfp127 gene. Gene 1999, 234, 323–327. [Google Scholar] [CrossRef]
- Buettner, V.L.; Walker, A.M.; Singer-Sam, J. Novel paternally expressed intergenic transcripts at the mouse Prader-Willi/Angelman Syndrome locus. Mamm. Genome 2005, 16, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Lai, L.; Duan, F.; Chen, M.; Deng, J.; Li, Z. Conservation of imprinting of MKRN3 and NAP1L5 in rabbits. Anim. Gen. 2016, 47, 507–509. [Google Scholar] [CrossRef]
- Hanna, C.W. Placental imprinting: Emerging mechanisms and functions. PLoS Genet. 2020, 16, e1008709. [Google Scholar] [CrossRef]
- John, R.M. Imprinted genes and the regulation of placental endocrine function: Pregnancy and beyond. Placenta 2017, 56, 86–90. [Google Scholar] [CrossRef] [PubMed]
- Monk, D. Genomic imprinting in the human placenta. Am. J. Obstet. Gynecol. 2015, 213, 152–162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noguer-Dance, M.; Abu-Amero, S.; Al-Khtib, M.; Lefèvre, A.; Coullin, P.; Moore, G.E.; Cavaillé, J. The primate-specific microRNA gene cluster (C19MC) is imprinted in the placenta. Hum. Mol. Genet. 2010, 19, 3566–3582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okae, H.; Hiura, H.; Nishida, Y.; Funayama, R.; Tanaka, S.; Chiba, H.; Yaegashi, N.; Nakayama, K.; Sasaki, H.; Arima, T. Re-investigation and RNA sequencing-based identification of genes with placenta-specific imprinted expression. Hum. Mol. Genet. 2012, 21, 548–558. [Google Scholar] [CrossRef] [Green Version]
- Pilvar, D.; Reiman, M.; Pilvar, A.; Laan, M. Parent-of-origin-specific allelic expression in the human placenta is limited to established imprinted loci and it is stably maintained across pregnancy. Clin. Epigenetics 2019, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Barlow, D.P. Genomic imprinting: A mammalian epigenetic discovery model. Annu. Rev. Genet. 2011, 45, 379–403. [Google Scholar] [CrossRef]
- White, C.R.; MacDonald, W.A.; Mann, M.R. Conservation of DNA Methylation Programming Between Mouse and Human Gametes and Preimplantation Embryos. Biol. Reprod. 2016, 95, 61. [Google Scholar] [CrossRef] [Green Version]
- Ferguson-Smith, A.C. Genomic imprinting: The emergence of an epigenetic paradigm. Nat. Rev. Genet. 2011, 12, 565–575. [Google Scholar] [CrossRef]
- Macdonald, W.A.; Mann, M.R.W. Epigenetic regulation of genomic imprinting from germ line to preimplantation. Mol. Reprod. Dev. 2014, 81, 126–140. [Google Scholar] [CrossRef]
- Geuns, E.; De Rycke, M.; Van Steirteghem, A.; Liebaers, I. Methylation imprints of the imprint control region of the SNRPN-gene in human gametes and preimplantation embryos. Hum. Mol. Genet. 2003, 12, 2873–2879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Maarri, O.; Buiting, K.; Peery, E.G.; Kroisel, P.M.; Balaban, B.; Wagner, K.; Urman, B.; Heyd, J.; Lich, C.; Brannan, C.I.; et al. Maternal methylation imprints on human chromosome 15 are established during or after fertilization. Nat. Genet. 2001, 27, 341–344. [Google Scholar] [CrossRef] [PubMed]
- El-Maarri, O.; Seoud, M.; Coullin, P.; Herbiniaux, U.; Oldenburg, J.; Rouleau, G.; Slim, R. Maternal alleles acquiring paternal methylation patterns in biparental complete hydatidiform moles. Hum. Mol. Genet. 2003, 12, 1405–1413. [Google Scholar] [CrossRef] [Green Version]
- Dubose, A.J.; Smith, E.Y.; Yang, T.P.; Johnstone, K.A.; Resnick, J.L. A new deletion refines the boundaries of the murine Prader-Willi syndrome imprinting center. Hum. Mol. Genet. 2011, 20, 3461–3466. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Horsthemke, B.; Wagstaff, J. Mechanisms of imprinting of the Prader-Willi/Angelman region. Am. J. Med. Genet. A 2008, 146, 2041–2052. [Google Scholar] [CrossRef]
- Rabinovitz, S.; Kaufman, Y.; Ludwig, G.; Razin, A.; Shemer, R. Mechanisms of activation of the paternally expressed genes by the Prader-Willi imprinting center in the Prader-Willi/Angelman syndromes domains. Proc. Natl. Acad. Sci. USA 2012, 109, 7403–7408. [Google Scholar] [CrossRef] [Green Version]
- Bressler, J.; Tsai, T.-F.; Wu, M.-Y.; Tsai, S.-F.; Ramirez, M.A.; Armstrong, D.; Beaudet, A.L. The SNRPN promoter is not required for genomic imprinting of the Prader-Willi/Angelman domain in mice. Nat. Genet. 2001, 28, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Sharp, A.J.; Migliavacca, E.; Dupré, Y.; Stathaki, E.; Sailani, M.R.; Baumer, A.; Schinzel, A.; Mackay, D.J.; Robinson, D.O.; Cobellis, G.; et al. Methylation profiling in individuals with uniparental disomy identifies novel differentially methylated regions on chromosome 15. Genome Res. 2010, 20, 1271–1278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chai, J.H.; Locke, D.P.; Ohta, T.; Greally, J.M.; Nicholls, R.D. Retrotransposed genes such as Frat3 in the mouse Chromosome 7C Prader-Willi syndrome region acquire the imprinted status of their insertion site. Mamm. Genome 2001, 12, 813–821. [Google Scholar] [CrossRef]
- Robinson, W.; Knoblauch, H.; Buiting, K.; Schmidt, K.; Gillessen-Kaesbach, G.; Horsthemke, B. Molecular diagnosis of the Prader-Willi and Angelman syndromes by detection of parent-of-origin specific DNA methylation in 15q11-13. Hum. Genet. 1992, 90, 313–315. [Google Scholar]
- O’Doherty, A.M.; O’Shea, L.C.; Fair, T. Bovine DNA methylation imprints are established in an oocyte size-specific manner, which are coordinated with the expression of the DNMT3 family proteins. Biol. Reprod. 2012, 86, 67. [Google Scholar] [CrossRef]
- Lucifero, D.; Suzuki, J.; Bordignon, V.; Martel, J.; Vigneault, C.; Therrien, J.; Filion, F.; Smith, L.C.; Trasler, J.M. Bovine SNRPN methylation imprint in oocytes and day 17 in vitro-produced and somatic cell nuclear transfer embryos. Biol. Reprod. 2006, 75, 531–558. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer | Sequence (5′-3′) | AT (°C) | Product Size (bp) | Application |
|---|---|---|---|---|
| MKRN3-F MKRN3-R | CGCCCGTCACGTCTGA TACAGCTCTGCCCACGAAAG | 55 | 568 | Identification of SNP rs42331804 and allelic expression analysis ofMKRN3 |
| MAGEL2-F1 MAGEL2-R1 | GAAAAACTTGCCTACCACATC CCACATCCCTGAGCAAGAGTA | 56 | 696 | Identification of SNP rs211249225 and allelic expression analysis ofMAGEL2in tissues |
| MAGEL2-F2 MAGEL2-R2 | CGTAGGCATTCTCTTCTCTC AACCTGTGACTGGATCTGC | 56 | 1045 | Identification of SNP rs110762305 and allelic expression analysis ofMAGEL2in placenta |
| NDN-F NDN-R | AGAAACACTCCACCTTCG CTACCCCAATACACAGCC | 55 | 644 | Identification of SNP rs468002089 and allelic expression analysis of NDN |
| PWS-IC-F1 PWS-IC-R1 | AAGGAAATTGATAGTAAGTATATTAGAGT AACCCAAATCCCCAATAAA | 59 | 791 | PWS-IC DMR identification and analysis |
| PWS-IC-F2 PWS-IC-R2 | GTTATTAGTGGAAAGTTTGAGGAAA ACCACACGACTAACCTAACCC | 59 | 472 | |
| MKRN3 DMR-F1 MKRN3 DMR-R1 | TGTAAGAATTATTAGAAAATAAAGAGTAGA CCCAATCCCTACTTCCTATACCTA | 54 | 541 | MKRN3 DMR identification and analysis |
| MKRN3 DMR-F2 MKRN3 DMR-R2 | TATATAGATATAAAATGAAGTGAATAAAG CCTACTTCCTTCTCTAAACAAA | 52 | 406 | |
| NDN DMR-F1 NDN DMR-R1 | AGTTTTAATAGGACGTTTGGGGAGG AATCTCCCTCTTCGCCTAAAACCTA | 59 | 526 | NDN DMR identification and analysis |
| NDN DMR-F2 NDN DMR-R2 | GGGAGTGATTATTGAGGTTTA ACAAAATTAAAATTACACACATCCTTACT | 59 | 331 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Chen, W.; Li, D.; Gu, S.; Liu, X.; Dong, Y.; Jin, L.; Zhang, C.; Li, S. Conservation of Imprinting and Methylation of MKRN3, MAGEL2 and NDN Genes in Cattle. Animals 2021, 11, 1985. https://doi.org/10.3390/ani11071985
Li J, Chen W, Li D, Gu S, Liu X, Dong Y, Jin L, Zhang C, Li S. Conservation of Imprinting and Methylation of MKRN3, MAGEL2 and NDN Genes in Cattle. Animals. 2021; 11(7):1985. https://doi.org/10.3390/ani11071985
Chicago/Turabian StyleLi, Junliang, Weina Chen, Dongjie Li, Shukai Gu, Xiaoqian Liu, Yanqiu Dong, Lanjie Jin, Cui Zhang, and Shijie Li. 2021. "Conservation of Imprinting and Methylation of MKRN3, MAGEL2 and NDN Genes in Cattle" Animals 11, no. 7: 1985. https://doi.org/10.3390/ani11071985
APA StyleLi, J., Chen, W., Li, D., Gu, S., Liu, X., Dong, Y., Jin, L., Zhang, C., & Li, S. (2021). Conservation of Imprinting and Methylation of MKRN3, MAGEL2 and NDN Genes in Cattle. Animals, 11(7), 1985. https://doi.org/10.3390/ani11071985