
Effects of Olive (Olea europaea L.) Leaves with Antioxidant and Antimicrobial Activities on In Vitro Ruminal Fermentation and Methane Emission

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. In Vitro Batch Fermentation
2.3. Total Polyphenols, Total Flavonoid Contents, and Antioxidant Activity
2.4. Assessment of Antimicrobial Assay
2.5. Analysis of Metabolites in Gas Chromatography-Mass Spectrometry (GC-MS)
2.6. Analysis of In Vitro Fermentation
2.7. Total DNA Extraction and Quantitative Real-Time Polymerase Chain Reaction
2.8. Statistical Analysis
3. Results
3.1. Chemical Composition of OLs and Their Total Phenolic and Flavonoid Contents
3.2. Carbohydrate Composition in OLs
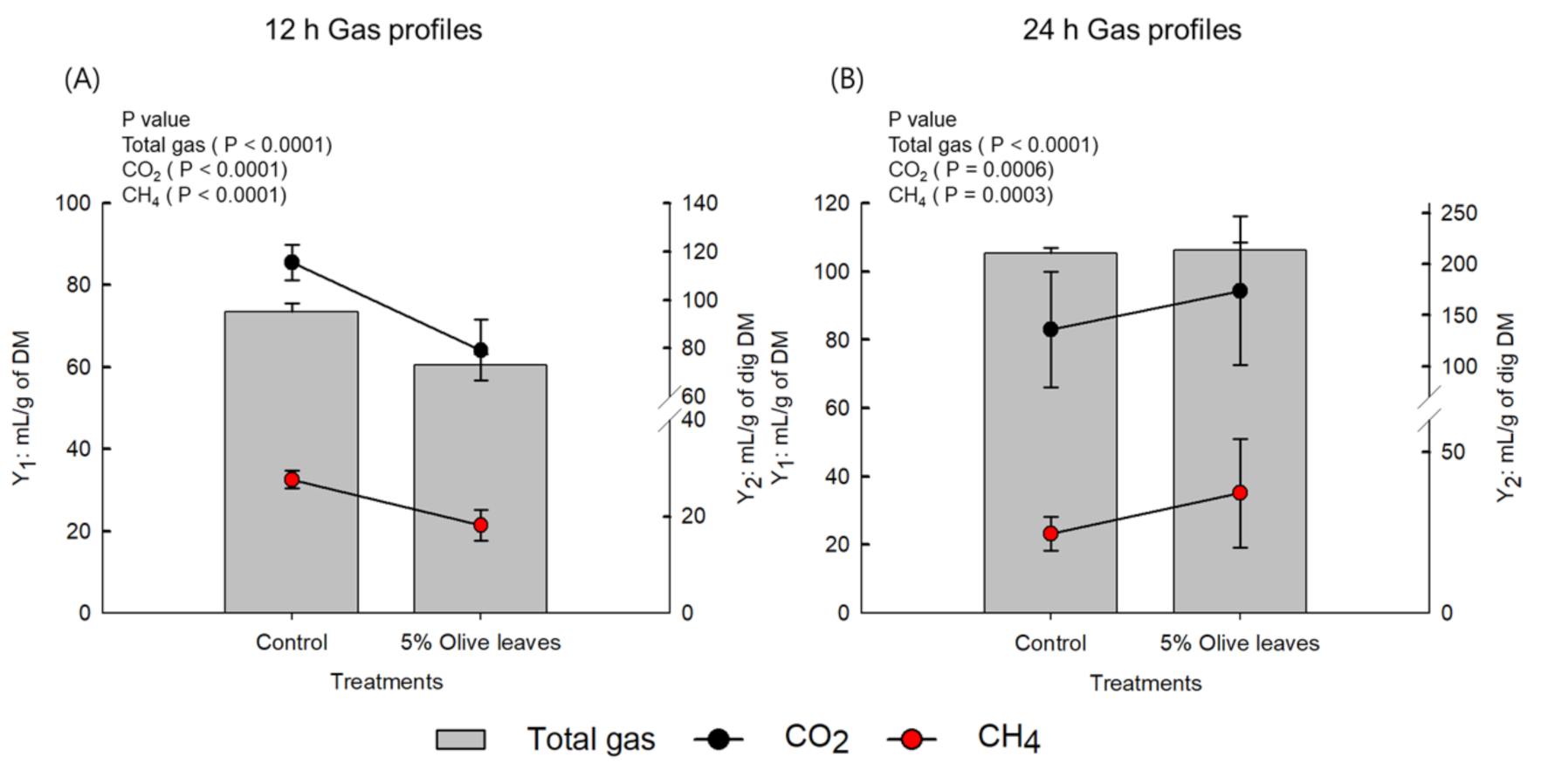
3.3. In Vitro Batch Fermentation
3.4. Antibacterial Activity
3.5. Effect of OLs on the Microbial Composition on In Vitro Batch Fermentation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Tabera, J.; Guinda, A.; Ruiz-Rodríguez, A.; Señoráns, F.J.; Ibáñez, E.; Albi, T.; Reglero, G. Countercurrent supercritical fluid extraction and fractionation of high-added-value compounds from a hexane extract of olive leaves. J. Agric. Food Chem. 2004, 52, 4774–4779. [Google Scholar] [CrossRef] [PubMed]
- Vogel, P.; Machado, I.K.; Garavaglia, J.; Zani, V.T.; de Souza, D.; Morelo dal Bosco, S. Polyphenols benefits of olive leaf (Olea europaea L.) to human health. Nutr. Hosp. 2015, 31, 1427–1433. [Google Scholar]
- Nilüfer, A.T.; Ağagündüz, D. Olive leaf (Olea europaea L. folium): Potential effects on glycemia and lipidemia. Ann. Nutr. Metab. 2020, 76, 10–15. [Google Scholar]
- Rosell’o-Soto, E.; Koubaa, M.; Moubarik, A.; Lopes, R.P.; Saraiva, J.A.; Boussetta, N.; Grimi, N.; Barba, F.J. Emerging opportunities for the effective valorization of wastes and by-products generated during olive oil production process: Nonconventional methods for the recovery of high-added value compounds. Trends Food Sci. Technol. 2015, 45, 296–310. [Google Scholar] [CrossRef]
- Moudache, M.; Colon, M.; Nerín, C.; Zaidi, F. Phenolic content and antioxidant activity of olive by-products and antioxidant film containing olive leaf extract. Food Chem. 2016, 212, 521–527. [Google Scholar] [CrossRef]
- Brahmi, F.; Mechri, B.; Dabbou, S.; Dhibi, M.; Hammami, M. Efficacy of phenolics compounds with different polarities as antioxidants from olive leaves depending on seasonal variations. Ind. Crops Prod. 2012, 38, 146–152. [Google Scholar] [CrossRef]
- El, S.N.; Karakaya, S. Olive tree (Olea europaea) leaves: Potential beneficial effects on human health. Nutr. Rev. 2009, 67, 632–638. [Google Scholar] [CrossRef] [PubMed]
- Bouaziz, M.; Fki, I.; Jemai, H.; Ayadi, M.; Sayadi, S. Effect of storage on refined and husk olive oils composition: Stabilization by addition of natural antioxidants from Chemlali olive leaves. Food Chem. 2008, 108, 253–262. [Google Scholar] [CrossRef]
- Khayyal, M.T.; el-Ghazaly, M.A.; Abdallah, D.M.; Nassar, N.N.; Okpanyi, S.N.; Kreuter, M.H. Blood pressure lowering effect of an olive leaf extract (Olea europaea) in L-NAME induced hypertension in rats. Arzneimittelforschung 2002, 52, 797–802. [Google Scholar]
- Martínez-Martos, J.M.; Mayas, M.D.; Carrera, P. Phenolic compounds oleuropein and hydroxytyrosol exert differential effects on glioma development via antioxidant defense systems. J. Funct. Foods 2014, 11, 221–234. [Google Scholar] [CrossRef]
- Puel, C.; Mathey, J.; Agalias, A. Dose-response study of effect of oleuropein, an olive oil polyphenol, in an ovariectomy/inflammation experimental model of bone loss in the rat. Clin. Nutr. 2006, 25, 859–868. [Google Scholar] [CrossRef] [PubMed]
- Molina-Alcaide, E.; Yáñez Ruiz, D.R. Potential use of olive by-products in ruminant feeding: A review. Anim. Feed. Sci. Technol. 2008, 147, 247–264. [Google Scholar] [CrossRef]
- Shakeri, P.; Durmic, Z.; Vadhanabhuti, J.; Vercoe, P.E. Products derived from olive leaves and fruits can alter in vitro ruminal fermentation and methane production. J. Sci. Food Agric. 2017, 97, 1367–1372. [Google Scholar] [CrossRef]
- Escalona, B.; Rocha, R.; García, J.; Carabano, R.; de-Blas, C. Characterization of in situ fibre digestion of several fibrous foods. Anim. Sci. 1999, 68, 217–221. [Google Scholar] [CrossRef]
- Yáñez Ruiz, D.R.; Martín García, A.I.; Moumen, A.; Molina Alcaide, E. Ruminal fermentation and degradation patterns, protozoa population and urinary purine derivatives excretion in goats and wethers fed diets based on olive leaves. J. Anim. Sci. 2004, 82, 3006–3014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martín García, A.I.; Moumen, A.; Yáñez Ruiz, D.R.; Molina Alcaide, E. Chemical composition and nutrients availability for goats and sheep of two-stage olive cake and olive leaves. Anim. Feed Sci. Technol. 2003, 107, 61–74. [Google Scholar] [CrossRef]
- AOAC. Official Methods of Analysis, 15th ed.; Association of Official Analytical Chemists: Arlington, VA, USA, 1990. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Choi, Y.Y.; Lee, S.J.; Lee, Y.J.; Kim, H.S.; Eom, J.S.; Kim, S.C.; Kim, E.T.; Lee, S.S. New challenges for efficient usage of Sargassum fusiforme for ruminant production. Sci. Rep. 2020, 10, 19655. [Google Scholar] [CrossRef]
- McDougall, E.I. Studies on ruminant saliva. 1. The composition and output of sheep’s saliva. Biochem. J. 1948, 43, 99–109. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Singleton, V.L.; Rossi, J.R. Colorimetry of total phenolics with phosphomolybdic-phosphotungstic acid reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar]
- Meda, A.; Lamien, C.E.; Romito, M.; Millogo, J.; Nacoulma, O.G. Determination of the total phenolic, flavonoid and proline contents in Burkina Fasan honey, as well as their radical scavenging activity. Food Chem. 2005, 91, 571–577. [Google Scholar] [CrossRef]
- Brand-Williams, W.; Cuvelier, M.E.; Berset, C. Use of a free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Re, R.; Pellegrini, N.; Proteggente, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Elizabeth, K.; Rao, M.N.A. Oxygen radical scavenging activity of curcumin. Int. J. Pharm. 1990, 58, 237–240. [Google Scholar]
- Lee, H.S. Antioxidative activity of browning reaction products isolated from storage-aged orange juice. J. Agric. Food Chem. 1992, 40, 550–552. [Google Scholar] [CrossRef]
- Kalemba, D.; Kunicka, A. Antibacterial and antifungal properties of essential oils. Curr. Med. Chem. 2003, 10, 813–829. [Google Scholar]
- European Medicines Agency. Harmonisation of European Breakpoints Set by MEA/CHMP and EUCAST; EMEA: London, UK, 2005. [Google Scholar]
- Li, Y.; Sun, M.; Li, Y.; Cheng, Y.; Zhu, W. Co-cultured methanogen improved the metabolism in the hydrogenosome of anaerobic fungus as revealed by gas chromatography-mass spectrometry analysis. Asian Australas. J. Anim. Sci. 2020, 33, 1948–1956. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.H.; Lee, S.J.; Oh, D.S.; Lee, I.D.; Eom, J.S.; Park, H.Y.; Choi, S.H.; Lee, S.S. In vitro evaluation of Rhus succedanea extracts for ruminants. Asian Australas. J. Anim. Sci. 2018, 31, 1635–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.; Morrison, M. Improved extraction of PCR-quality community DNA from digesta and fecal samples. Biotechniques 2004, 36, 808–812. [Google Scholar] [CrossRef] [PubMed]
- Denman, S.E.; McSweeney, C.S. Development of a real-time PCR assay for monitoring anaerobic fungal and cellulolytic bacterial populations within the rumen. FEMS Microbiol. Ecol. 2006, 58, 572–582. [Google Scholar] [CrossRef]
- Sylvester, J.T.; Karnati, S.K.; Yu, Z.; Morrison, M.; Firkins, J.L. Development of an assay to quantify rumen ciliate protozoal biomass in cows using real-time PCR. J. Nutr. 2004, 134, 3378–3384. [Google Scholar] [CrossRef] [Green Version]
- Rong-Fu, W.; Wei-Wen, C.; Cerniglia, C.E. PCR detection of Ruminococcus spp. in human and animal faecal samples. Mol. Cell. Probes 1997, 11, 259–265. [Google Scholar]
- Stevenson, D.M.; Weimer, P.J. Dominance of Prevotella and low abundance of classical ruminal bacterial species in the bovine rumen revealed by relative quantification real-time PCR. Appl. Microbiol. Biotechnol. 2007, 75, 165–174. [Google Scholar] [CrossRef]
- Paillard, D.; McKain, N.; Chaudhary, L.C.; Walker, N.D.; Pizette, F.; Koppova, I.; McEwan, N.R.; Kopecný, J.; Vercoe, P.E.; Louis, P.; et al. Relation between phylogenetic position, lipid metabolism and butyrate production by different Butyrivibrio-like bacteria from the rumen. Antonie Van Leeuwenhoek 2007, 91, 417–422. [Google Scholar] [CrossRef]
- Tajima, K.; Aminov, R.I.; Nagamine, T.; Matsui, H.; Nakamura, M.; Benno, Y. Diet-dependent shifts in the bacterial population of the rumen revealed with real-time PCR. Appl. Environ. Microbiol. 2016, 7, 2766–2774. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Whelan, J.A.; Russell, N.B.; Whelan, M.A. A method for the absolute quantification of cDNA using real-time PCR. J. Immunol. Methods 2003, 278, 261–269. [Google Scholar] [CrossRef]
- SAS Institute Inc. SAS/STAT User’s Guide: Version 9.2; SAS Institute Inc.: Cary, NC, USA, 2002. [Google Scholar]
- Lee, O.H.; Lee, B.Y. Antioxidant and antimicrobial activities of individual and combined phenolics in Olea europaea leaf extract. Bioresour. Technol. 2010, 101, 3751–3754. [Google Scholar] [CrossRef] [PubMed]
- Brahmi, F.; Flamini, G.; Issaoui, M.; Dhibi, M.; Dabbou, S.; Mastouri, M.; Hammamoi, M. Chemical composition and biological activities of volatile fractions from three Tunisian cultivars of olive leaves. Med. Chem. Res. 2012, 21, 2863–2872. [Google Scholar] [CrossRef]
- Hatice, Z.; Ayse, H.B. Antibacterial and antioxidant activity of essential oil terpenes against pathogenic and spoilage-forming bacteria and cell structure-activity relationships evaluated by SEM microscopy. Molecules 2014, 19, 17773–17798. [Google Scholar]
- Sudjana, A.N.; d’Orazio, C.; Ryan, V.; Rasool, N.; Ng, J.; Islam, N.; Riley, T.V.; Hammer, K.A. Antimicrobial activity of commercial Olea europaea (olive) leaf extract. Int. J. Antimicrob. Agents 2009, 33, 461–463. [Google Scholar] [CrossRef]
- Techathuvanan, C.; Reyes, F.; David, J.R.; Davidson, P.M. Efficacy of commercial natural antimicrobials alone and in combinations against pathogenic and spoilage microorganisms. J. Food Prot. 2014, 77, 269–275. [Google Scholar] [CrossRef]
- Mothana, R.A.; Lindequist, U. Antimicrobial activity of some medicinal plants of the island Soqotra. J. Ethnopharmacol. 2005, 96, 177–181. [Google Scholar] [CrossRef]
- Benchaar, C.; McAllister, T.A.; Chouinard, P.Y. Digestion, ruminal fermentation, ciliate protozoal populations, and milk production from dairy cows fed cinnamaldehyde, quebracho condensed tannin, or yucca schidigera saponin extracts. J. Dairy Sci. 2008, 91, 4765–4777. [Google Scholar] [CrossRef]
- Lee, O.H.; Lee, H.B.; Son, J.Y. Antimicrobial activities and nitrite-scavenging ability of olive leaf fractions. Korean J. Soc. Food Cook. Sci. 2004, 20, 86–92. [Google Scholar]
- Davis, W.W.; Stout, T.R. Disc plate method of microbiological antibiotic assay: Ⅰ. Factors influencing variability and error. Appl. Microbiol. 1971, 22, 659–665. [Google Scholar] [CrossRef]
- Lee-Huang, S.; Zhang, L.; Huang, P.L.; Chang, Y.T.; Huang, P.L. Anti-HIV activity of olive leaf extract (OLE) and modulation of host cell gene expression by HIV-1 infection and OLE treatment. Biochem. Biophys. Res. Commun. Actions 2003, 307, 1029–1037. [Google Scholar] [CrossRef]
- Eom, J.S.; Lee, S.J.; Lee, Y.; Kim, H.S.; Choi, Y.Y.; Kim, H.S.; Kim, D.H.; Lee, S.S. Effects of supplementation levels of Allium fistulosum L. extract on in vitro ruminal fermentation characteristics and methane emisson. Peer J. 2020, 8, e9651. [Google Scholar] [CrossRef] [PubMed]
- Ernesto Vargas, J.; Andrés, S.; López-Ferreras, L.; Snelling, T.J.; Yáñez-Ruíz, D.R.; García-Estrada, C.; López, S. Dietary supplemental plant oils reduce methanogenesis from anaerobic microbial fermentation in the rumen. Sci. Rep. 2020, 10, 1613. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tapio, I.; Snelling, T.J.; Strozzi, F.; Wallace, R.J. The ruminal microbiome associated with methane emissions from ruminant livestock. J. Anim. Sci. Biotechnol. 2017, 8, 7. [Google Scholar] [CrossRef]
- Denis, O.K.; Stuart, E.D.; Roderick, I.M.; Mark, M.; Ann, L.R.; Graeme, T.A.; Christopher, S.M. Opportunities to improve fiber degradation in the rumen: Microbiology, ecology, and genomics. FEMS Microbiol. Rev. 2003, 27, 663–693. [Google Scholar]


{kind=link}
| Target Species | Primer | Sequence (5′ → 3′) | Size (bp) | Reference |
|---|---|---|---|---|
| General bacteria | F | CGGCAACGAGCGCAACCC | 130 | [32] |
| R | CCATTGTAGCACGTGTGTAGCC | |||
| Ciliate protozoa | F | GCTTTCGWTGGTAGTGTATT | 223 | [33] |
| R | CTTGCCCTCYAATCGTWCT | |||
| Fungi | F | GAGGAAGTAAAAGTCGTAACAAGGTTTC | 120 | [32] |
| R | CAAATTCACAAAGGGTAGGATGATT | |||
| Methanogenic archaea | F | GAGGAAGGAGTGGACGACGGTA | 232 | [33] |
| R | ACGGGCGGTGTGTGCAAG | |||
| Fibrobacter succinogenes | F | GTTCGGAATTACTGGGCGTAAA | 121 | [32] |
| R | CGCCTGCCCCTGAACTATC | |||
| Ruminococcus albus | F | CCCTAAAAGCAGTCTTAGTTCG | 176 | [34] |
| R | CCTCCTTGCGGTTAGAACA | |||
| Ruminococcus flavefaciens | F | CGAACGGAGATAATTTGAGTTTACTTAGG | 132 | [32] |
| R | CGGTCTCTGTATGTTATGAGGTATTACC | |||
| Prevotella ruminicola | F | GCGAAAGTCGGATTAATGCTCTATG | 78 | [33] |
| R | CCCATCCTATAGCGGTAAACCTTTG | |||
| Butyrivibrio fibrisolvens | F | ACCGCATAAGCGCACGGA | 65 | [35] |
| R | CGGGTCCATCTTGTACCGATAAAT | |||
| Butyrivibrio proteoclasticus | F | TCCGGTGGTATGAGATGGGC | 185 | [36] |
| R | GTCGCTGCATCAGAGTTTCCT | |||
| Anearovibrio lipolytica | F | TGGGTGTTAGAAATGGATTC | 597 | [37] |
| R | CTCTCCTGCACTCAAGAATT |
| Items | Olive leaves (Mean ± SEM) |
|---|---|
| Chemical composition | |
| Dry matter (DM) | 94.59 ± 0.03 |
| Crude protein | 10.87 ± 0.49 |
| Ether extract | 8.80 ± 0.30 |
| Crude ash | 8.28 ± 0.17 |
| Neutral detergent fiber | 38.82 ± 0.81 |
| Acid detergent fiber | 27.35 ± 0.11 |
| Antioxidant activity | |
| Total polyphenol (mg catechin/g extract) | 34.79 ± 2.72 |
| Total flavonoid (mg quercetin/g extract) | 5.91 ± 0.24 |
| IC50 for DPPH (μg/mL) 1 | 78.14 |
| IC50 for ABTS (μg/mL) 2 | 33.21 |
| Content 1 | Plant Concentration (µg/mL, Mean ± SEM) | SEM 2 | p-Value | Contrast 3 | |||||
|---|---|---|---|---|---|---|---|---|---|
| 10 | 50 | 100 | 200 | L | Q | C | |||
| DPPH | 33.18 ± 1.52 c | 47.37 ± 1.15 b | 53.21 ± 0.81 a | 54.25 ± 0.43 a | 1.06 | <0.0001 | <0.0001 | <0.0001 | 0.0434 |
| ABTS | 82.40 ± 0.94 d | 87.84 ± 0.24 c | 90.80 ± 0.09 b | 92.36 ± 0.11 a | 0.38 | <0.0001 | <0.0001 | <0.0001 | 0.0859 |
| HO | 63.62 ± 0.47 d | 104.15 ± 0.12 c | 265.78 ± 0.04 b | 596.4 ± 0.06 a | 6.18 | <0.0001 | <0.0001 | <0.0001 | 0.0004 |
| NO | 15.29 ± 0.75 d | 19.75 ± 1.07 c | 23.85 ± 0.33 b | 27.94 ± 0.46 a | 0.71 | <0.0001 | <0.0001 | 0.0087 | 0.9259 |
| RT (min) | Compound | Formula | Area (%) | MW(g/mol) 1 | Class 2 |
|---|---|---|---|---|---|
| 5.471 | Lactic acid | C3H6O3 | 0.38 | 90.080 | OA |
| 8.894 | Glycerol | C3H8O3 | 0.72 | 92.094 | FA |
| 16.497 | Fructose | C6H12O6 | 1.44 | 180.160 | CHO |
| 16.599 | Fructose | C6H12O6 | 1.10 | 180.160 | CHO |
| 16.704 | Galactose | C6H12O6 | 0.47 | 180.156 | CHO |
| 16.77 | Glucose | C6H12O6 | 10.42 | 180.156 | CHO |
| 16.981 | Glucose | C6H12O6 | 2.06 | 180.156 | CHO |
| 17.12 | Sorbitol | C6H14O6 | 26.30 | 182.170 | CHO |
| 17.189 | Ethyl-alpha-glucopyranoside | C8H16O6 | 3.57 | 208.210 | CHO |
| 18.411 | Palmitic acid | C16H32O2 | 6.84 | 256.400 | OA |
| 18.778 | Myo-inositol | C6H12O6 | 0.95 | 180.160 | CA |
| 20.309 | Stearic acid | C18H36O2 | 6.04 | 284.480 | OA |
| 23.552 | Sucrose | C12H22O11 | 39.71 | 342.300 | CHO |
| Parameters 1 | Treatments | SEM 2 | p Value | |
|---|---|---|---|---|
| Control | 5% OLs | |||
| 12 h | ||||
| pH | 7.11 b | 7.28 a | 0.02 | 0.0020 |
| DMD (%) | 38.00 a | 33.67 b | 1.07 | 0.0459 |
| MGR (OD 550 nm) | 0.30 | 0.26 | 0.01 | 0.0843 |
| Ammonia (mg∙dL−1) | 9.80 b | 13.49 a | 0.25 | <0.0001 |
| Total VFA (mM) | 48.73 a | 45.54 b | 0.45 | 0.0023 |
| Acetate (mM) | 31.54 a | 29.28 b | 0.19 | 0.0002 |
| Propionate (mM) | 9.72 | 9.59 | 0.11 | 0.4291 |
| Butyrate (mM) | 7.47 a | 6.67 b | 0.19 | 0.0238 |
| A/P ratio | 3.24 a | 3.05 b | 0.03 | 0.0022 |
| 24 h | ||||
| pH | 6.61 | 6.66 | 0.02 | 0.3034 |
| DMD (%) | 51.92 a | 49.17 b | 0.69 | 0.0311 |
| MGR (OD 550 nm) | 0.35 | 0.27 | 0.03 | 0.1557 |
| Ammonia (mg∙dL−1) | 11.69 b | 13.02 a | 0.19 | 0.0011 |
| Total VFA (mM) | 63.26 | 62.85 | 0.16 | 0.1153 |
| Acetate (mM) | 41.75 | 41.64 | 0.20 | 0.6941 |
| Propionate (mM) | 13.22 a | 12.58 b | 0.05 | 0.0002 |
| Butyrate (mM) | 8.28 b | 8.63 a | 0.10 | 0.0469 |
| A/P ratio | 3.16 b | 3.31 a | 0.02 | 0.0029 |
| Paper Disk | Microorganisms Tested | |||
|---|---|---|---|---|
| Staphylococcus aureus ATCC 6538 | Escherichia Coli ATCC 8739 | Klebsiella Pneumoniae ATCC 4352 | Pseudomonas aeruginosa ATCC 10145 | |
| Conc. (μg/disc) | 50 | 50 | 50 | 50 |
| Clear Zone (mm) | 10 | 8 | 8.5 | 8 |
| Items | Fermentation Time (h) | Control | 5% Olive leaves | SEM 1 | p-Value |
|---|---|---|---|---|---|
| Absolute abundance 2 | |||||
| Total bacteria | 12 | 3.17 a | 2.38 b | 0.15 | 0.0212 |
| 24 | 4.65 a | 2.70 b | 0.38 | 0.0216 | |
| Fungi | 12 | 34.63 | 33.69 | 10.90 | 0.9543 |
| 24 | 5.52 a | 0.66 b | 0.70 | 0.0079 | |
| Ciliate protozoa | 12 | 1.45 | 3.82 | 0.85 | 0.1184 |
| 24 | 0.97 | 1.09 | 0.43 | 0.8569 | |
| Methanogenic archaea | 12 | 10.15 | 8.87 | 0.90 | 0.3712 |
| 24 | 16.60 a | 5.84 b | 0.36 | <0.0001 | |
| Relative proportion, % total bacteria | |||||
| Fibrobacter succinogenes | 12 | 10.54 | 10.98 | 1.44 | 0.8313 |
| 24 | 13.20 a | 0.53 b | 0.38 | <0.0001 | |
| Ruminococcus albus | 12 | 4.27 b | 24.12 a | 0.54 | <0.0001 |
| 24 | 5.29 a | 2.23 b | 0.34 | <0.0001 | |
| Ruminococcus flavefaciens | 12 | 0.86 b | 1.02 a | 0.03 | 0.0012 |
| 24 | 0.66 | 0.63 | 0.02 | 0.2469 | |
| Prevotella ruminicola | 12 | 26.22 b | 31.04 a | 1.48 | 0.0349 |
| 24 | 38.97 a | 28.09 b | 1.07 | <0.0001 | |
| Butyrivibrio fibrisolvens | 12 | 1.99 a | 1.17 b | 0.08 | 0.0012 |
| 24 | 2.78 a | 1.26 b | 0.21 | 0.2469 | |
| Butyrivibrio proteoclasticus | 12 | 0.36 a | 0.26 b | 0.01 | <0.0001 |
| 24 | 0.46 a | 0.32 b | 0.01 | <0.0001 | |
| Anearovibrio lipolytica | 12 | 0.26 b | 1.16 a | 0.08 | <0.0001 |
| 24 | 0.93 b | 4.34 a | 0.27 | <0.0001 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.J.; Kim, H.S.; Eom, J.S.; Choi, Y.Y.; Jo, S.U.; Chu, G.M.; Lee, Y.; Seo, J.; Kim, K.H.; Lee, S.S. Effects of Olive (Olea europaea L.) Leaves with Antioxidant and Antimicrobial Activities on In Vitro Ruminal Fermentation and Methane Emission. Animals 2021, 11, 2008. https://doi.org/10.3390/ani11072008
Lee SJ, Kim HS, Eom JS, Choi YY, Jo SU, Chu GM, Lee Y, Seo J, Kim KH, Lee SS. Effects of Olive (Olea europaea L.) Leaves with Antioxidant and Antimicrobial Activities on In Vitro Ruminal Fermentation and Methane Emission. Animals. 2021; 11(7):2008. https://doi.org/10.3390/ani11072008
Chicago/Turabian StyleLee, Shin Ja, Hyun Sang Kim, Jun Sik Eom, You Young Choi, Seong Uk Jo, Gyo Moon Chu, Yookyung Lee, Jakyeom Seo, Kyoung Hoon Kim, and Sung Sill Lee. 2021. "Effects of Olive (Olea europaea L.) Leaves with Antioxidant and Antimicrobial Activities on In Vitro Ruminal Fermentation and Methane Emission" Animals 11, no. 7: 2008. https://doi.org/10.3390/ani11072008
APA StyleLee, S. J., Kim, H. S., Eom, J. S., Choi, Y. Y., Jo, S. U., Chu, G. M., Lee, Y., Seo, J., Kim, K. H., & Lee, S. S. (2021). Effects of Olive (Olea europaea L.) Leaves with Antioxidant and Antimicrobial Activities on In Vitro Ruminal Fermentation and Methane Emission. Animals, 11(7), 2008. https://doi.org/10.3390/ani11072008