
Physiological Correlation between Hypothalamic–Pituitary–Adrenal Axis, Leptin, UCP1 and Lipid Panel in Mares during Late Pregnancy and Early Postpartum Period

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Experimental Design
2.2. Data Collection and Laboratory Analysis
2.3. Statistical Analysis
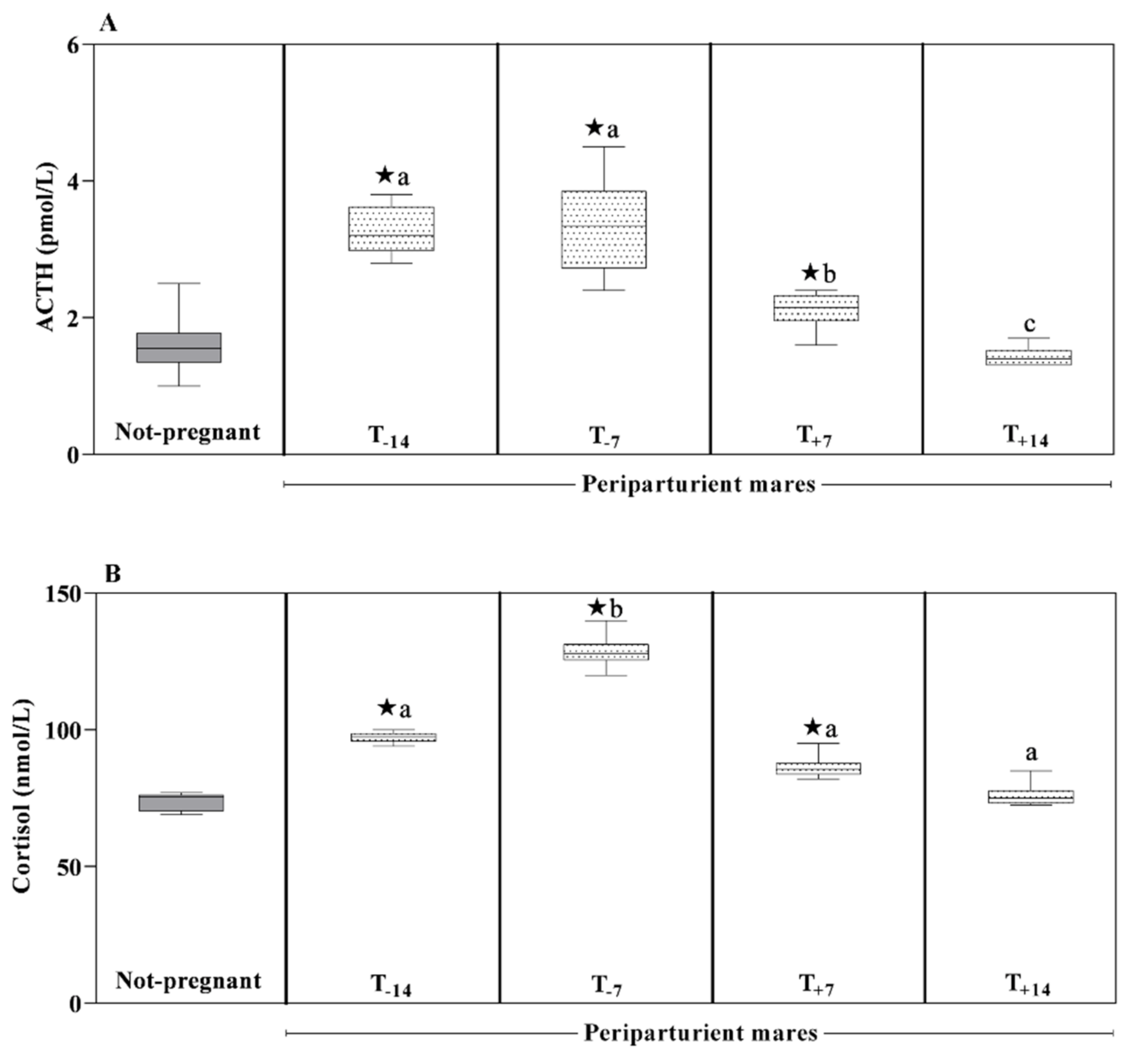
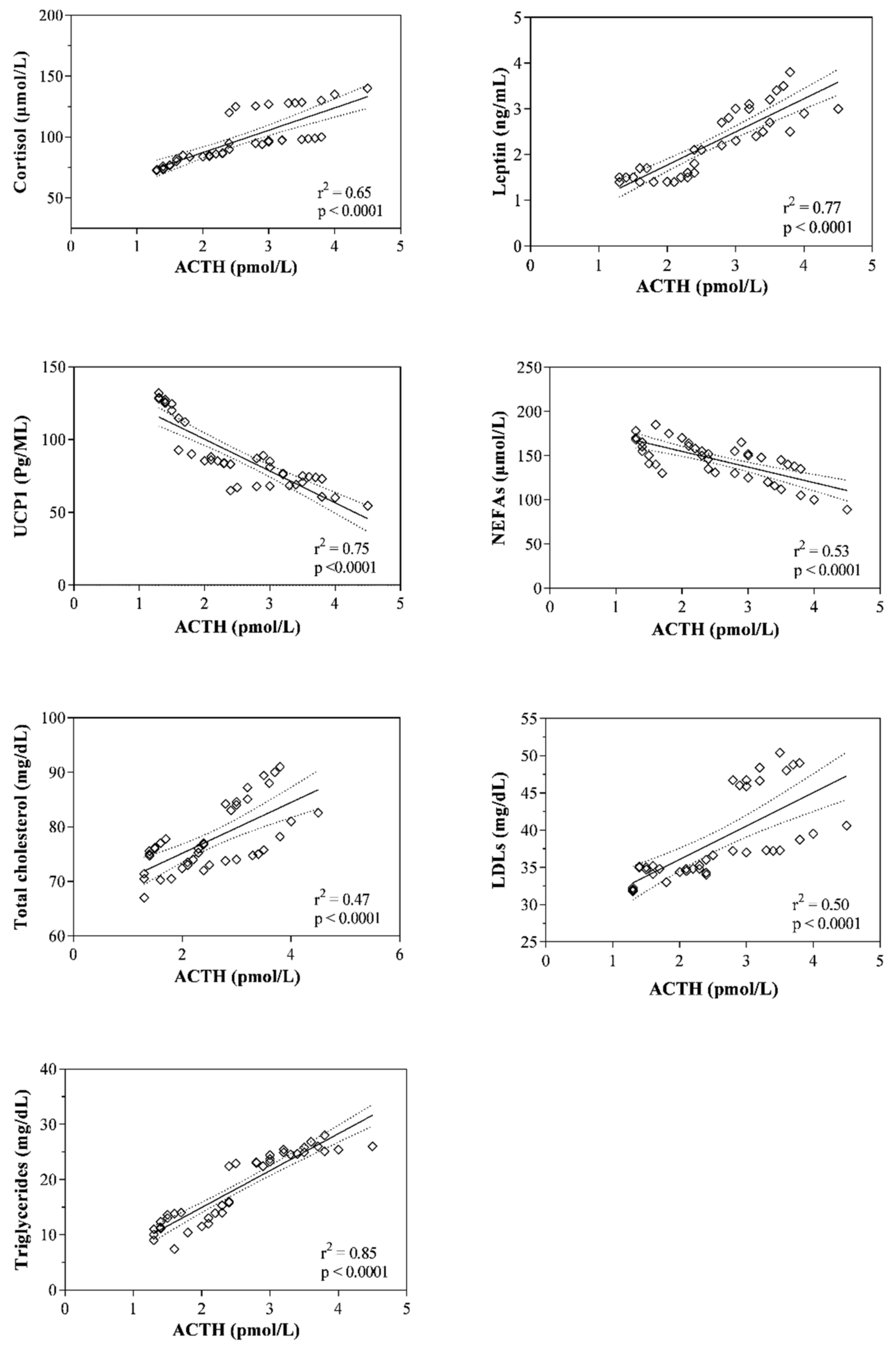
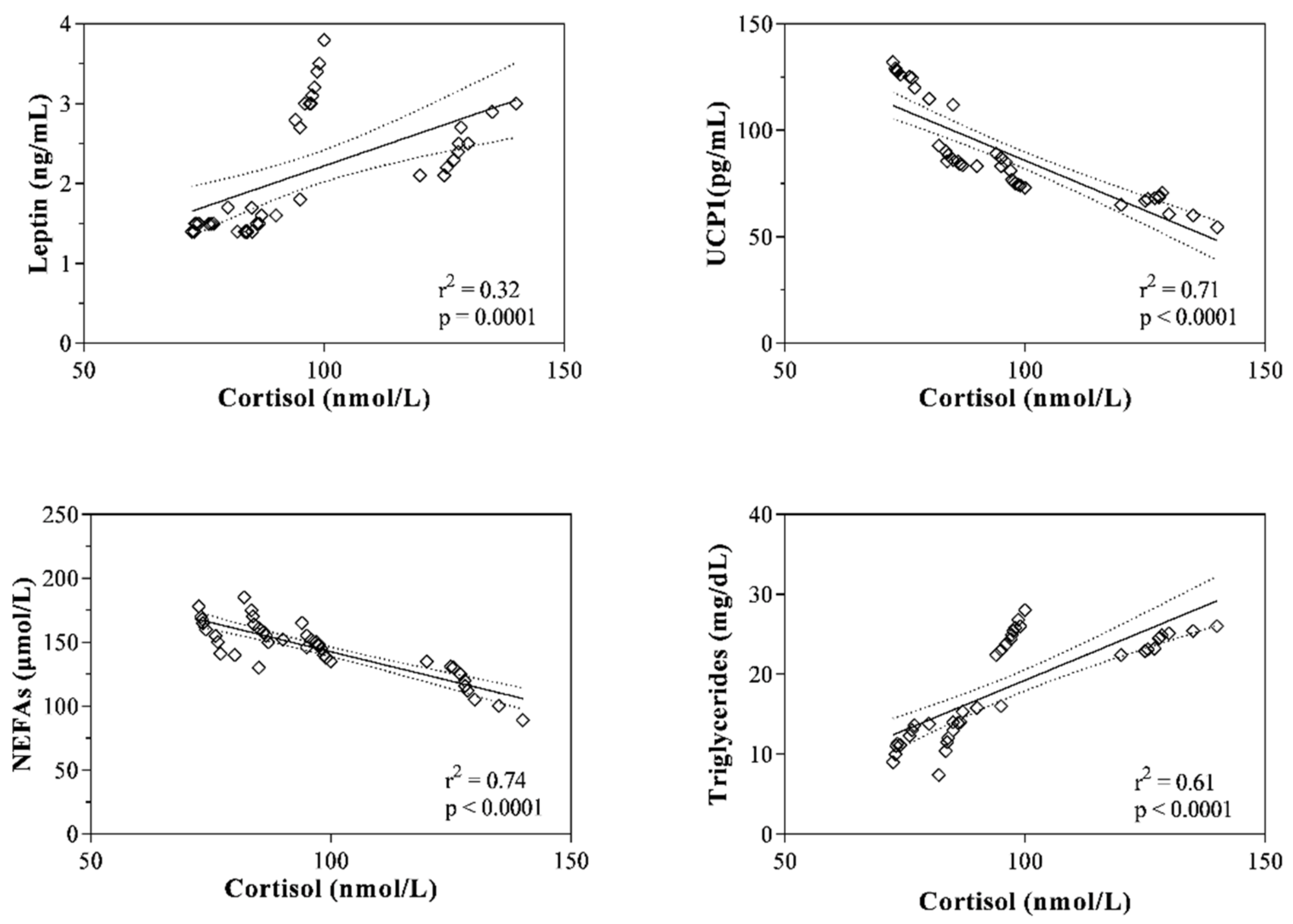
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Knottenbelt, D.C.; Holdstock, N.; Madigan, J.E. Equine Neonatology, Medicine and Surgery, 1st ed.; Saunders: Edinburgh, UK, 2004. [Google Scholar]
- Aoki, T.; Ishii, M. Hematologic and biochemical profiles in peripartum mares and neonatal foals (Heavy Draft Horse). J. Equine. Vet. Sci. 2012, 32, 170–176. [Google Scholar] [CrossRef]
- Özçelik, E.; Uslu, S.; Kebapc, N.; Kara, M.; Dokumacıoğlu, A.; Musmul, A. Interrelations of serum leptin levels with adrenocorticotropic hormone, basal cortisol and dehydroepiandrosterone sulphate levels in patients with metabolic syndrome. Diabetes Metab. Syndr. 2010, 4, 13–17. [Google Scholar] [CrossRef]
- Holubová, A.; Štofková, A.; Jurčovičová, J.; Šlamberová, R. The effect of neonatal maternal stress on plasma levels of adrenocorticotropic hormone, corticosterone, leptin, and ghrelin in adult male rats exposed to acute heterotypic stressor. Physiol. Res. 2016, 65, S557–S566. [Google Scholar] [CrossRef] [PubMed]
- Salzmann, C.; Otis, M.; Long, H.; Roberge, C.; Gallo-Payet, N.; Walker, C.D. Inhibition of steroidogenic response to adrenocorticotropin by leptin: Implications for the adrenal response to maternal separation in neonatal rats. Endocrinology 2004, 145, 1810–1822. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.V.; Levine, S.; Alam, S.; Harbich, D.; Sterlemann, V.; Ganea, K.; De Kloet, E.R.; Holsboer, F.; Muller, M.B. Metabolic signals modulate hypothalamic-pituitary-adrenal axis activation during maternal separation of the neonatal mouse. J. Neuroendocr. 2006, 18, 865–874. [Google Scholar] [CrossRef]
- Baltaci, A.K.; Vurucu, N.; Uzun, A.; Mogulkoc, R.; Kilic, M. The effect of acute swimming exercise on plasma leptin in rats. Bratisl. Lek. Listy. 2012, 113, 592–594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zareian, P.; Karimi, M.V.; Dorneyani, G. The comparison of the effects of acute swimming stress on plasma corticosterone and leptin concentration in male and female rats. Acta Med. Iran. 2011, 49, 284–287. [Google Scholar]
- Arfuso, F.; Giannetto, C.; Rizzo, M.; Fazio, F.; Giudice, E.; Piccione, G. Serum levels of mitochondrial uncoupling protein 1, leptin, and lipids during late pregnancy and the early postpartum period in mares. Theriogenology 2016, 86, 1156–1164. [Google Scholar] [CrossRef]
- Murakami, T.; Yamashita, T.; Iida, M.; Kuwajima, M.; Shima, K. A short form of leptin receptor performs signal transduction. Biochem. Biophys. Res. Commun. 1997, 231, 26–29. [Google Scholar] [CrossRef]
- Spinedi, E.; Gaillard, R.C. A regulatory loop between the hypothalamo-pituitary-adrenal (HPA) axis and circulating leptin: A physiological role of ACTH. Endocrinology 1998, 139, 4016–4020. [Google Scholar] [CrossRef]
- Giacobino, J.P. Uncoupling proteins, leptin, and obesity: An updated review. Ann. N. Y. Acad. Sci. 2002, 967, 398–402. [Google Scholar] [CrossRef]
- Brondani, L.A.; Assmann, T.S.; Coutinho, G.; Duarte, K.; Gross, J.L.; Canani, L.H.; Crispim, D. The role of the uncoupling protein 1 (UCP1) on the development of obesity and type 2 diabetes mellitus. Arq. Bras. Endocrinol. Metabol. 2012, 56, 215–225. [Google Scholar] [CrossRef] [Green Version]
- Bazzano, M.; Giannetto, C.; Fazio, F.; Arfuso, F.; Giudice, E.; Piccione, G. Metabolic profile of broodmares during late pregnancy and early post-partum. Reprod. Domest. Anim. 2014, 49, 947–953. [Google Scholar] [CrossRef]
- Bazzano, M.; Giannetto, C.; Fazio, F.; Marafioti, S.; Giudice, E.; Piccione, G. Hemostatic profile during late pregnancy and early postpartum period in mares. Theriogenology 2014, 81, 639–943. [Google Scholar] [CrossRef]
- Fazio, E.; Medica, P.; Cravana, C.; Bruschetta, G.; Ferlazzo, A. Seasonal thyroid and lipid profiles in Thoroughbred pregnant and nonpregnant mares (Equus caballus). Theriogenology 2016, 85, 1582–1589. [Google Scholar] [CrossRef] [PubMed]
- Cartmill, J.A.; Thompson, D.L., Jr.; Storer, W.A.; Gentry, L.R.; Huff, N.K. Endocrine responses in mares and geldings with high body condition scores grouped by high vs. low resting leptin concentrations. J. Anim. Sci. 2003, 81, 2311–2321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henneke, D.R.; Potter, G.D.; Kreider, J.L.; Yeates, B.F. Relationship between condition score, physical measurement and body fat percentage in mares. Equine. Vet. J. 1983, 15, 371–372. [Google Scholar] [CrossRef] [PubMed]
- Giannetto, C.; Fazio, F.; Alberghina, D.; Assenza, A.; Panzera, M.; Piccione, G. Different daily patterns of serum cortisol and locomotor activity rhythm in horses under natural photoperiod. J. Vet. Behav. 2015, 10, 118–121. [Google Scholar] [CrossRef]
- Giannetto, C.; Fazio, F.; Alberghina, D.; Panzera, M.; Piccione, G. Constant darkness disrupt daily rhythm of adrenocorticotrophin in horses. J. Appl. Biomed. 2013, 11, 41–45. [Google Scholar] [CrossRef] [Green Version]
- Friedewald, W.T.; Levy, R.I.; Fredrickson, D.S. Estimation of the concentration of low–density lipoprotein cholesterol in plasma, without the use of the preparative ultracentrifuge. Clin. Chem. 1972, 18, 499–502. [Google Scholar] [CrossRef] [PubMed]
- Maniam, J.; Antoniadis, C.; Morris, M.J. Early-life stress, HPA axis adaptation, and mechanisms contributing to later health outcomes. Front. Endocrinol. 2014, 5, 73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pralong, F.P.; Gaillard, R.C. Neuroendocrine effects of leptin. Pituitary 2001, 4, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, B.; Gentz, F.; Failing, K. Investigations into the course of progesterone-, oestrogen-and eCG- concentrations during normal and impaired pregnancy in the mare. Reprod. Domest. Anim. 1996, 31, 717–723. [Google Scholar] [CrossRef]
- Mastorakos, G.; Ilias, I. Maternal and fetal Hypothalamic–Pituitary–Adrenal Axes during pregnancy and postpartum. Ann. N. Y. Acad. Sci. 2003, 997, 136–149. [Google Scholar] [CrossRef] [PubMed]
- Ousey, J.C. Peripartal endocrinology in the mare and foetus. Reprod. Domest. Anim. 2004, 39, 222–231. [Google Scholar] [CrossRef] [PubMed]
- Torner, L.; Toschi, N.; Nava, G.; Clapp, C.; Neumann, I.D. Increased hypothalamic expression of prolactin in lactation: Involvement in behavioral and neuroendocrine stress responses. Eur. J. Neurosci. 2002, 15, 1381–1389. [Google Scholar] [CrossRef] [PubMed]
- Pi, X.J.; Voogt, J.L. Effect of suckling on prolactin receptor immunoreactivity in the hypothalamus of the rat. Neuroendocrinology 2000, 71, 308–317. [Google Scholar] [CrossRef]
- Bosch, O.J.; Musch, W.; Bredewold, R.; Slattery, D.A.; Neumann, I.G. Prenatal stress increases HPA axis activity and impairs maternal care in lactating female offspring: Implication for postpartum mood disorder. Psychoneuroendocrinology 2007, 32, 267–278. [Google Scholar] [CrossRef]
- Watson, T.D.; Burns, L.; Packard, C.J.; Shepherd, J. Effects of pregnancy and lactation on plasma lipid and lipoprotein concentrations, lipoprotein composition and post-heparin lipase activities in Shetland pony mares. J. Reprod. Fertil. 1993, 97, 563–568. [Google Scholar] [CrossRef] [Green Version]
- Schwartz, M.W.; Seeley, R.J. The new biology of body weight regulation. J. Am. Diet. Assoc. 1997, 97, 54–58. [Google Scholar] [CrossRef]
- Dallman, M.F.; Akana, S.F.; Strack, A.M.; Hanson, E.S.; Sebastian, R.J. The neural network that regulates energy balance is responsive to glucocorticoids and insulin and also regulates HPA axis responsivity at a site proximal to CRF neurons. Ann. N. Y. Acad. Sci. 1995, 771, 730–742. [Google Scholar] [CrossRef] [PubMed]
- Guillaume-Gentil, C.; Assimacopoulos-Jeannet, F.; Jeanrenaud, B. Involvement of non-esterified fatty acid oxidation in glucocorticoid-induced peripheral insulin resistance in vivo in rats. Diabetologia 1993, 36, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Glasow, A.; Bornstein, S.R. Leptin and the adrenal gland. Eur. J. Clin. Investig. 2000, 3, 39–45. [Google Scholar] [CrossRef]
- Heiman, M.L.; Ahima, R.S.; Craft, L.S.; Schoner, B.; Stephens, T.W.; Flier, J.S. Leptin inhibition of the hypothalamic-pituitary-adrenal axis in response to stress. Endocrinology 1997, 138, 3859–3863. [Google Scholar] [CrossRef] [PubMed]
- Bornstein, S.R.; Uhlmann, K.; Haidan, A.; Ehrhart-Bornstein, M.; Scherbaum, W.A. Evidence for a novel peripheral action of leptin as a metabolic signal to the adrenal gland: Leptin inhibits cortisol release directly. Diabetes 1997, 46, 1235–1238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wabitsch, M.; Jensen, P.B.; Blum, W.F.; Christoffersen, C.T.; Englaro, P.; Heinze, E.; Rascher, W.; Teller, W.; Tornqvist, H.; Hauner, H. Insulin and cortisol promote leptin production in cultured human fat cells. Diabetes 1996, 45, 1435–1438. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papaspyrou-Rao, S.; Schneider, S.H.; Petersen, R.N.; Fried, S.K. Dexamethasone increases leptin expression in humans in vivo. J. Clin. Endocrinol. Metab. 1997, 82, 1635–1637. [Google Scholar] [CrossRef]
- Friedman, M.; Rosenman, R.H.; Carroll, V. Changes in the serum cholesterol and blood clotting time in men subjected to cyclic variation of occupational stress. Circulation 1958, 17, 852–861. [Google Scholar] [CrossRef] [Green Version]
- Buff, P.R.; Dodds, A.C.; Morrison, C.D.; Whitley, N.C.; McFadin, E.L.; Daniel, J.A.; Djiane, J.; Keisler, D.H. Leptin in horses: Tissue localization and relationship between peripheral concentrations in leptin and body condition. J. Anim. Sci. 2002, 80, 2942–2948. [Google Scholar] [CrossRef] [Green Version]
- Carter, R.A.; McCutcheon, L.J.; George, L.A.; Smith, T.L.; Frank, N.; Geor, R.J. Effects of diet-induced gain on insulin sensitivity and plasma hormone and lipid concentrations in horses. Am. J. Vet. Res. 2009, 70, 1250–1258. [Google Scholar] [CrossRef]
- Amico, J.A.; Thomas, A.; Crowley, R.S.; Burmeister, L.A. Concentrations of leptin in the serum of pregnant, lactating, and cycling rats and of leptin messenger ribonucleic acid in rat placental tissue. Life Sci. 1998, 63, 1387–1395. [Google Scholar] [CrossRef]
- Ehrhardt, R.A.; Slepetis, R.M.; Bell, A.W.; Boisclair, Y.R. Maternal leptin is elevated during pregnancy in sheep. Domest. Anim. Endocrinol. 2001, 21, 85–96. [Google Scholar] [CrossRef]
- Kędzierski, W.; Kapica, M. Plasma concentration of leptin and ghrelin in Standardbred foals as related to the age, sex, exercise and training. Animal 2008, 2, 582–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kędzierski, W.; Kusy, R.; Kowalik, S. Plasma leptin level in hyperlipidemic mares and their newborn foals. Reprod. Domest. Anim. 2011, 46, 275–280. [Google Scholar] [CrossRef] [PubMed]
- Berg, E.L.; McNamara, D.L.; Keisler, D.H. Endocrine profiles of periparturient mares and their foals. J. Anim. Sci. 2007, 85, 1660–1668. [Google Scholar] [CrossRef]
- Heidler, B.; Parvizi, N.; Sauerwein, H.; Bruckmaier, R.M.; Heintges, U.; Aurich, J.E.; Aurich, C. Effects of lactation on metabolic and reproductive hormones in Lipizzaner mares. Domest. Anim. Endocrinol. 2003, 25, 47–59. [Google Scholar] [CrossRef]
- Thomas, L.; Wallace, J.M.; Aitken, R.P.; Mercer, J.G.; Trayhurn, P.; Hoggard, N. Circulating leptin during ovine pregnancy in relation to maternal nutrition, body composition, and pregnancy outcome. J. Endocrinol. 2001, 169, 465–476. [Google Scholar] [CrossRef] [Green Version]
- Jakimiuk, A.J.; Skalba, P.; Huterski, D.; Tarkowski, R.; Haczynski, J.; Magoffin, D.A. Leptin messenger ribonucleic acid (mRNA) content in the human placenta at term: Relationship to levels of leptin in cord blood and placental weight. Gynecol. Endocrinol. 2003, 17, 311–316. [Google Scholar] [CrossRef]
- Ramsay, T.G. Porcine leptin inhibits lipogenesis in porcine adipocytes. J. Anim. Sci. 2003, 81, 3008–3017. [Google Scholar] [CrossRef]
- Contreras, C.; Gonzalez, F.; Fernø, J.; Diéguez, C.; Rahmouni, K.; Nogueiras, R.; López, M. The brain and brown fat. Ann. Med. 2015, 47, 150–168. [Google Scholar] [CrossRef] [Green Version]
- Morrison, S.F.; Madden, C.J.; Tupone, D. Central neural regulation of brown adipose tissue thermogenesis and energy expenditure. Cell Metab. 2014, 19, 741–756. [Google Scholar] [CrossRef] [Green Version]
- Martinez de Morentin, P.B.; Lage, R.; González-García, I.; Ruíz-Pino, F.; Martins, L.; Fernández-Mallo, D.; Gallego, R.; Fernø, J.; Señarís, R.; Saha, A.K.; et al. Pregnancy induces resistance to the anorectic effect of hypothalamic malonyl–CoA and the thermogenic effect of hypothalamic AMPK inhibition in female rats. Endocrinology 2015, 156, 947–960. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lopez, M.; Tena–Sempere, M. Estrogens and the control of energy homeostasis: A brain perspective. Trends Endocrinol. Metab. 2015, 26, 411–421. [Google Scholar] [CrossRef] [PubMed]
- Oftedal, O.T.; Hintz, H.F.; Schryver, H.F. Lactation in the horse: Milk composition and intake by foals. J. Nutr. 1983, 113, 2096–2106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harvey, J.W.; Pate, M.G.; Kivipelto, J.; Asquith, R.L. Clinical biochemistry of pregnant and nursing mares. Vet. Clin. Pathol. 2005, 34, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.L.; Lear, S.R.; Forte, T.M.; Ko, W.; Massimi, M.; Erickson, S.K. Effect of pregnancy and lactation on lipoprotein and cholesterol metabolism in the rat. J. Lipid. Res. 1998, 39, 2237–2249. [Google Scholar] [CrossRef]
- Kaneko, J.J.; Harvey, J.W.; Bruss, M.L. Clinical Biochemistry of Domestic Animals, 5th ed.; Academic Press Inc.: San Diego, CA, USA, 1997. [Google Scholar]
- Cupps, P.T. Reproduction in Domestic Animals; Academic Press: San Diego, CA, USA, 1991. [Google Scholar]
- Barello, C.; Garoffo, L.P.; Montorfano, G.; Zava, S.; Berra, B.; Conti, A.; Giuffrida, M.G. Analysis of major proteins and fat fractions associated with mare’s milk fat globules. Mol. Nutr. Food. Res. 2008, 52, 1448–1456. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cortisol (nmol/L) | Leptin (ng/mL) | UCP1 (pg/mL) | NEFAs (µmol/L) | Total Cholesterol (mg/dL) | HDLs (mg/dL) | LDLs (mg/dL) | Triglycerides (mg/dL) | |
|---|---|---|---|---|---|---|---|---|
| ACTH (pmol/L) | r = 0.81 p < 0.0001 | r = 0.88 p < 0.0001 | r = −0.87 p < 0.0001 | r = −0.73 p < 0.0001 | r = 0.68 p < 0.0001 | r = 0.07 p = 0.67 | r = 0.71 p < 0.0001 | r = 0.92 p < 0.0001 |
| Cortisol (nmol/L) | r = 0.57 p = 0.0001 | r = −0.86 p < 0.0001 | r= −0.86 p < 0.0001 | r= 0.22 p = 0.19 | r = −0.14 p = 0.37 | r = 0.28 p = 0.08 | r = 0.71 p < 0.0001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Arfuso, F.; Giannetto, C.; Bazzano, M.; Assenza, A.; Piccione, G. Physiological Correlation between Hypothalamic–Pituitary–Adrenal Axis, Leptin, UCP1 and Lipid Panel in Mares during Late Pregnancy and Early Postpartum Period. Animals 2021, 11, 2051. https://doi.org/10.3390/ani11072051
Arfuso F, Giannetto C, Bazzano M, Assenza A, Piccione G. Physiological Correlation between Hypothalamic–Pituitary–Adrenal Axis, Leptin, UCP1 and Lipid Panel in Mares during Late Pregnancy and Early Postpartum Period. Animals. 2021; 11(7):2051. https://doi.org/10.3390/ani11072051
Chicago/Turabian StyleArfuso, Francesca, Claudia Giannetto, Marilena Bazzano, Anna Assenza, and Giuseppe Piccione. 2021. "Physiological Correlation between Hypothalamic–Pituitary–Adrenal Axis, Leptin, UCP1 and Lipid Panel in Mares during Late Pregnancy and Early Postpartum Period" Animals 11, no. 7: 2051. https://doi.org/10.3390/ani11072051
APA StyleArfuso, F., Giannetto, C., Bazzano, M., Assenza, A., & Piccione, G. (2021). Physiological Correlation between Hypothalamic–Pituitary–Adrenal Axis, Leptin, UCP1 and Lipid Panel in Mares during Late Pregnancy and Early Postpartum Period. Animals, 11(7), 2051. https://doi.org/10.3390/ani11072051