
Fat Deposition in the Muscle of Female and Male Yak and the Correlation of Yak Meat Quality with Fat

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples Collection
2.2. Meat Quality Measurements
2.3. Determination of Fat Content and Fatty Acid (FA) Profiles
2.4. Transcriptome Analyses
2.5. Protein Preparation and Digestion
2.6. Reversed-Phase Liquid Chromatography (RPLC) Separation and Mass Spectrum (MS) Analysis
2.7. Statistical Analysis
3. Results
3.1. Meat Quality
3.2. Fat Content and FA Profiles
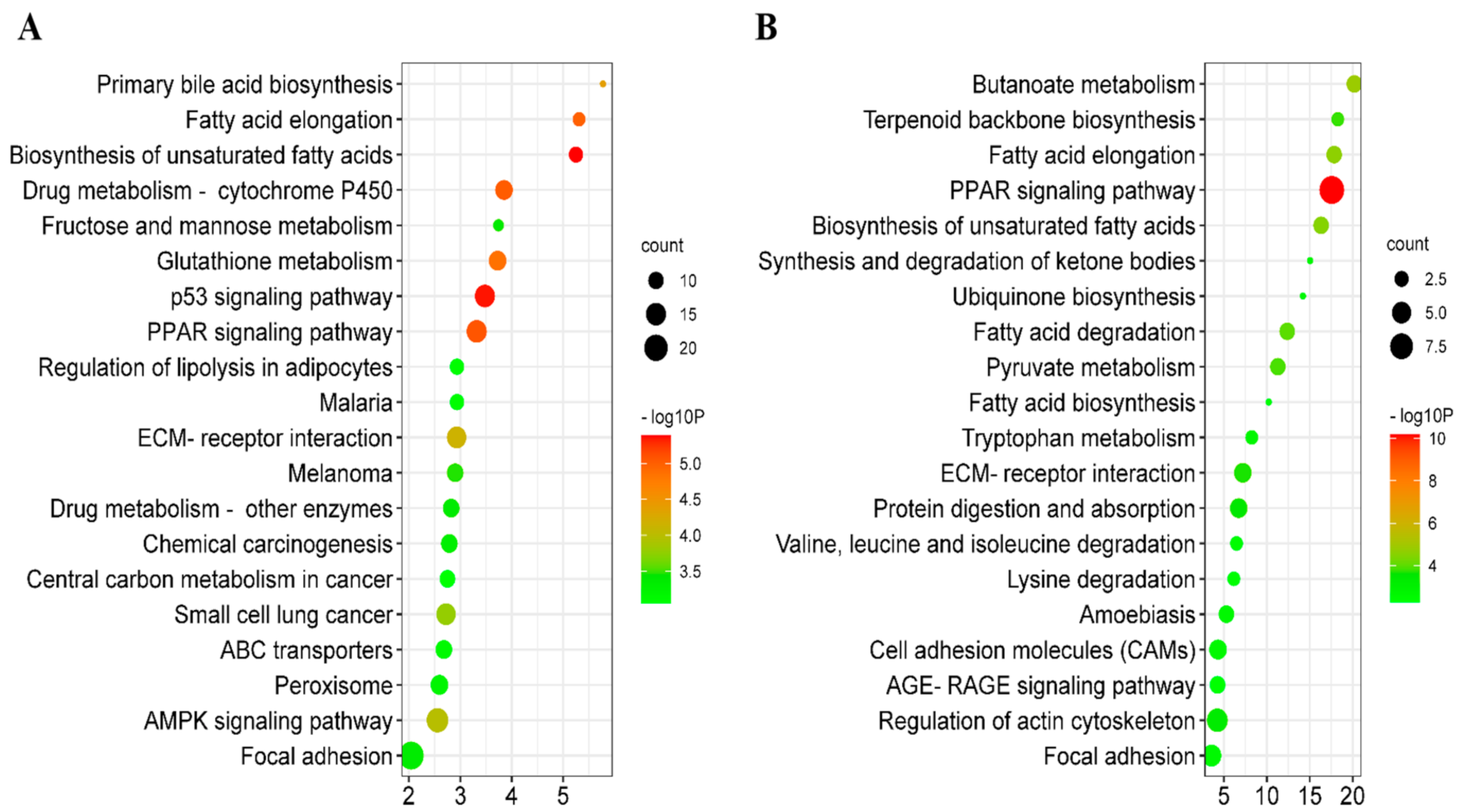
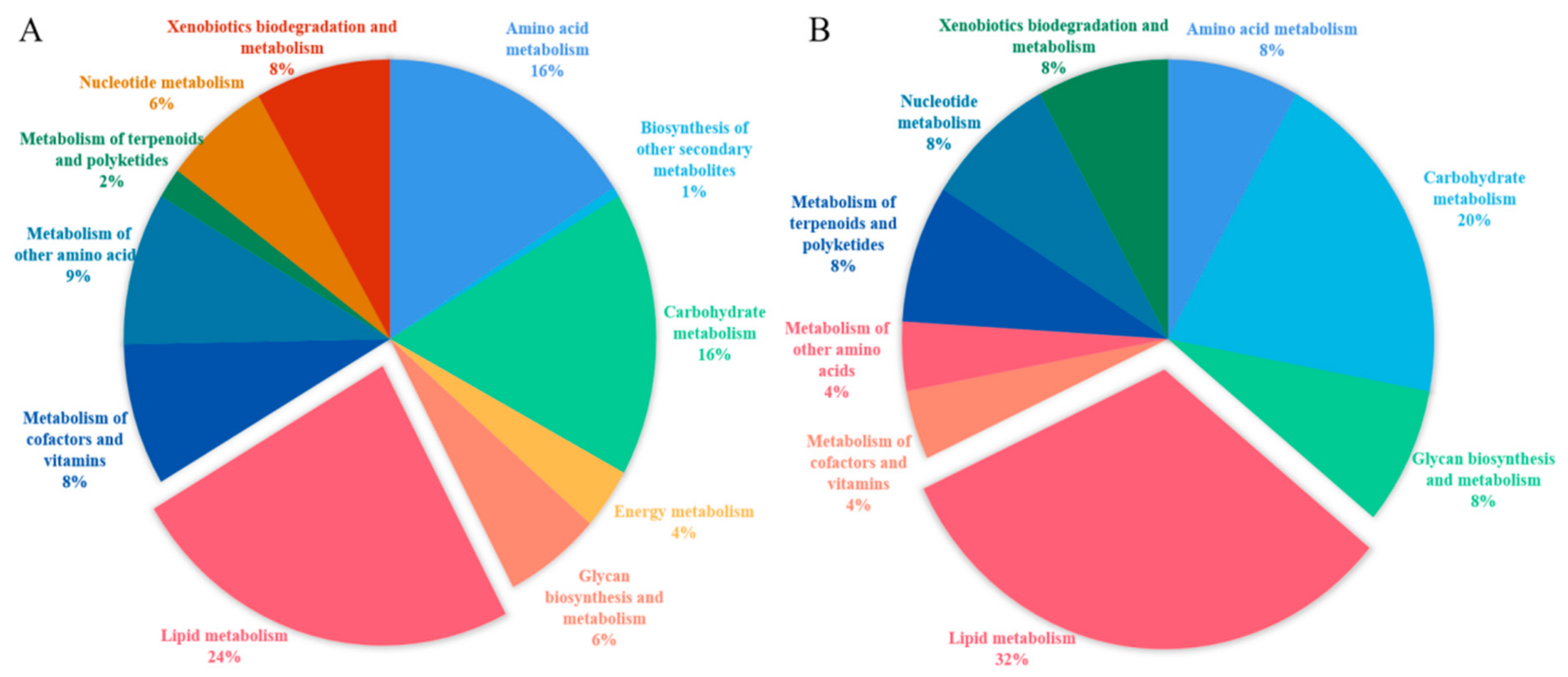
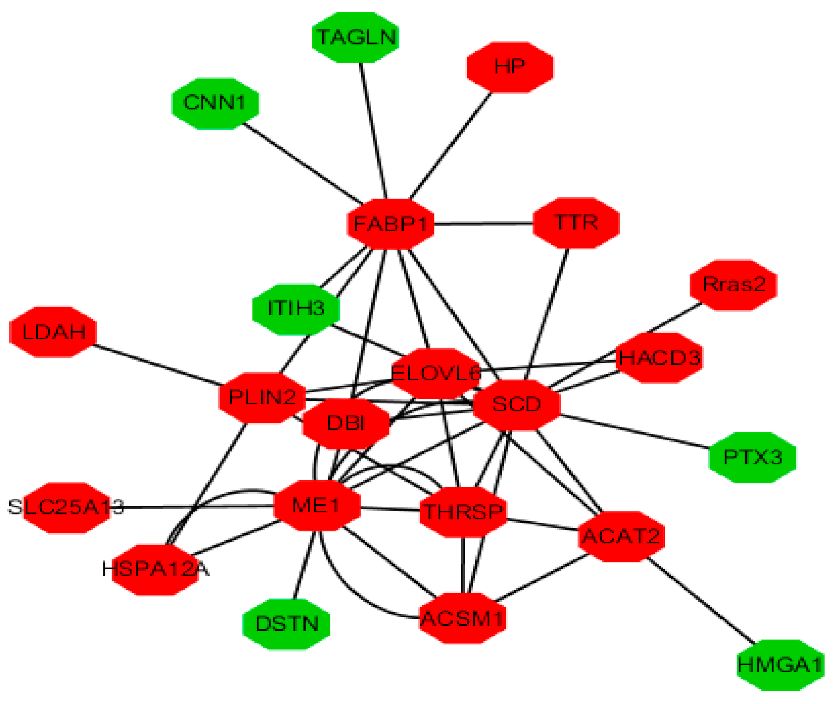
3.3. Function Enrichment Analysis for Differentially Expressed Genes (DEGs) and Differentially Expressed Proteins (DEPs)
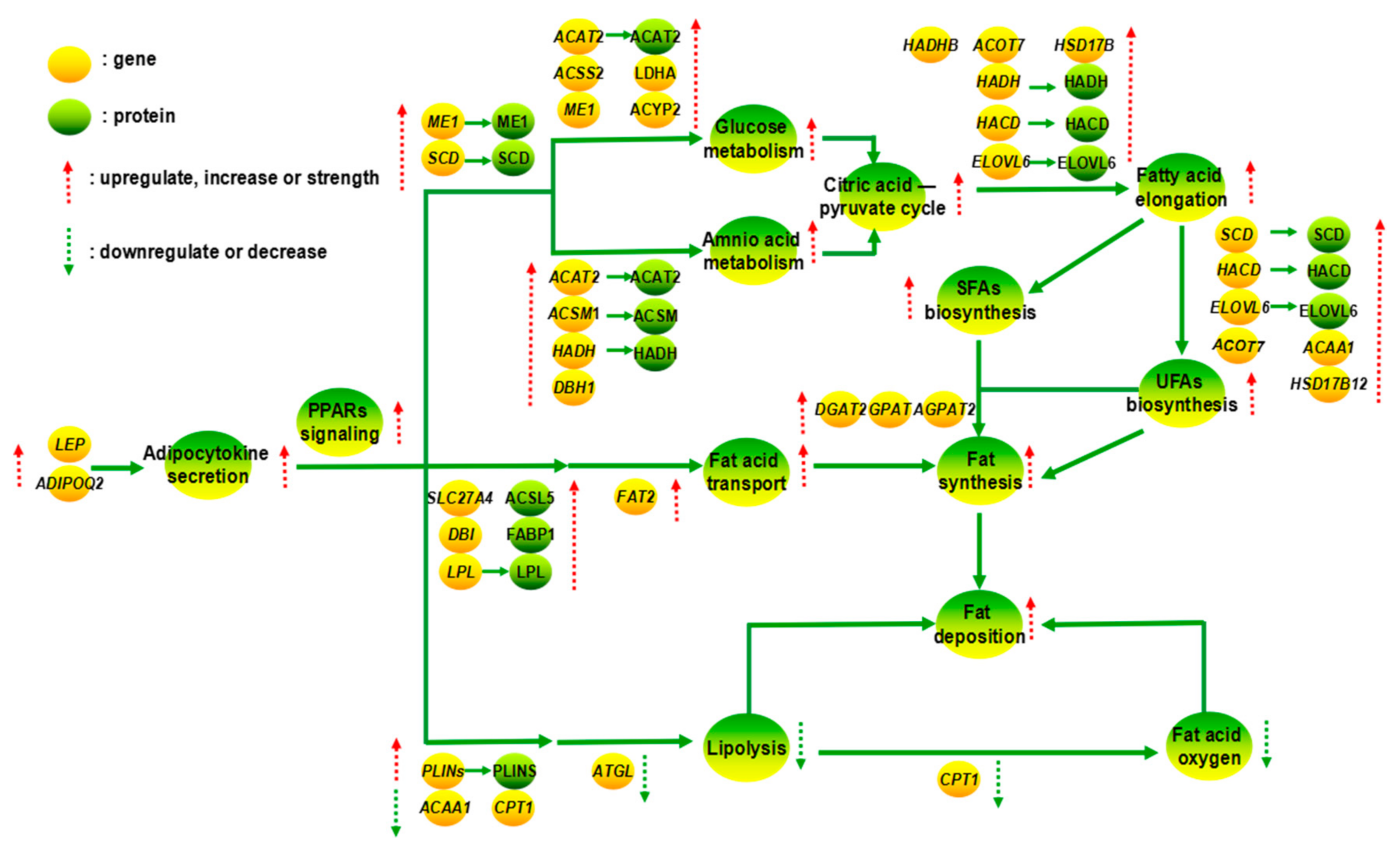
4. Discussion
4.1. Correlation of Fat with Meat Quality
4.2. Effect of Gender to Fat Deposition in Yak
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gagaoua, M.; Bonnet, M.; Picard, B. Protein array-based approach to evaluate biomarkers of beef tenderness and marbling in cows: Understanding of the underlying mechanisms and prediction. Foods 2020, 9, 1180. [Google Scholar] [CrossRef] [PubMed]
- Pewan, S.B.; Otto, J.R.; Huerlimann, R.; Budd, A.M.; Mwangi, F.W.; Edmunds, R.C.; Holman, B.W.B.; Henry, M.L.E.; Kinobe, R.; Adegboye, O.A.; et al. Genetics of omega-3 long-chain polyunsaturated fatty acid metabolism and meat eating quality in Tattykeel Australian white lambs. Genes 2020, 11, 587. [Google Scholar] [CrossRef] [PubMed]
- Ekine-Dzivenu, C.; Vinsky, M.; Basarab, J.A.; Aalhus, J.L.; Dugan, M.E.R.; Li, C. Phenotypic and genetic correlations of fatty acid composition in subcutaneous adipose tissue with carcass merit and meat tenderness traits in Canadian beef cattle1. J. Anim. Sci. 2017, 95, 5184–5196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wen, Y.; Liu, H.; Liu, K.; Cao, H.; Mao, H.; Dong, X.; Yin, Z. Analysis of the physical meat quality in partridge (Alectoris chukar) and its relationship with intramuscular fat. Poult. Sci. 2020, 99, 1225–1231. [Google Scholar] [CrossRef]
- Douny, C.; El Khoury, R.; Delmelle, J.; Brose, F.; Degand, G.; Moula, N.; Farnir, F.; Clinquart, A.; Maghuin-Rogister, G.; Scippo, M. Effect of storage and cooking on the fatty acid profile of omega-3 enriched eggs and pork meat marketed in Belgium. Food Sci. Nutr. 2014, 3, 140–152. [Google Scholar] [CrossRef]
- Liu, S.; Huang, J.; Wang, X.; Ma, Y. Transcription factors regulate adipocyte differentiation in beef cattle. Anim. Genet. 2020, 51, 351–357. [Google Scholar] [CrossRef]
- Guo, Q.; Kong, X.F.; Hu, C.J.; Zhou, B.; Wang, C.T.; Shen, Q.W. Fatty acid content, flavor compounds, and sensory quality of pork loin as affected by dietary supplementation with larginine and glutamic acid. J. Food Sci. 2019, 84, 3445–3453. [Google Scholar] [CrossRef]
- Cawthorn, D.M.; Hoffman, L.C. Controversial cuisine: A global account of the demand, supply and acceptance of “unconventional” and “exotic” meats. Meat Sci. 2016, 120, 19–36. [Google Scholar] [CrossRef]
- Maggiolino, A.; Lorenzo, J.M.; Centoducati, G.; Domínguez, R.; Dinardo, F.R.; Marino, R.; Malva, A.D.; Bragaglio, A.; Palo, P.D. How volatile compounds, oxidative profile and sensory evaluation can change with vacuum aging in donkey meat. Animals 2020, 10, 2126. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Munekata, P.E.S.; Campagnol, P.C.B.; Zhu, Z.Z.; Alpas, H.; Barba, F.J.; Tomasevic, I. Technological aspects of horse meat products-a review. Food Res. Int. 2017, 102, 176–183. [Google Scholar] [CrossRef]
- Popoola, I.O.; Soladoye, P.O.; Gaudette, N.J.; Wismer, W.V. A Review of Sensory and Consumer-related Factors Influencing the Acceptance of Red Meats from Alternative Animal Species. Food Rev. Int. 2020, 36, 1–20. [Google Scholar] [CrossRef]
- Luo, J.; Huang, Z.Y.; Liu, H.N.; Zhang, Y.; Ren, F.Z. Yak milk fat globules from the Qinghai-Tibetan Plateau: Membrane lipid composition and morphological properties. Food Chem. 2018, 245, 731–737. [Google Scholar] [CrossRef] [PubMed]
- Leslie, D.M.; Schaller, G.B. Bos grunniens and Bos mutus (Artiodactyla: Bovidae). Mamm. Spec. 2009, 836, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Wen, W.; Luo, X.; Xia, B.; Guan, J.; Nie, Y.; Li, L.; Duan, J.; Suman, S.; Sun, Q. Post-mortem oxidative stability of three yak (Bos grunniens) muscles as influenced by animal age. Meat Sci. 2015, 105, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Long, R.; Kreuzer, M.; Ding, L.; Shang, Z.; Zhang, Y.; Yang, Y.; Cui, G. Importance of Functional Ingredients in Yak Milk-Derived Food on Health of Tibetan Nomads Living Under High-Altitude Stress: A Review. Crit. Rev. Food Sci. Nutr. 2013, 54, 292–302. [Google Scholar] [CrossRef]
- Wang, L.-L.; Han, L.; Ma, X.-L.; Yu, Q.-L.; Zhao, S.-N. Effect of mitochondrial apoptotic activation through the mitochondrial membrane permeability transition pore on yak meat tenderness during postmortem aging. Food Chem. 2017, 234, 323–331. [Google Scholar] [CrossRef]
- Brzozowska, A.M.; Lukaszewicz, M.; Oprzadek, J.M. Energy-protein supplementation and lactation affect fatty acid profile of liver and adipose tissue of dairy cows. Molecules 2018, 23, 618. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; He, Y.; Li, H.; Wu, F.; Qiu, Q.; Niu, W.; Gao, Z.; Su, H.; Cao, B. Rumen fermentation, intramuscular fat fatty acid profiles and related rumen bacterial populations of Holstein bulls fed diets with different energy levels. Appl. Microbiol. Biotechnol. 2019, 103, 4931–4942. [Google Scholar] [CrossRef]
- Stergiadis, S.; Berlitz, C.B.; Hunt, B.; Garg, S.; Givens, D.I.; Kliem, K.E. An update to the fatty acid profiles of bovine retail milk in the United Kingdom: Implications for nutrition in different age and gender groups. Food Chem. 2019, 276, 218–230. [Google Scholar] [CrossRef]
- Oliveira, P.P.A.; Corte, R.; Silva, S.; Rodriguez, P.; Sakamoto, L.; Pedroso, A.; Tullio, R.; Berndt, A. The effect of grazing system intensification on the growth and meat quality of beef cattle in the Brazilian Atlantic Forest biome. Meat Sci. 2018, 139, 157–161. [Google Scholar] [CrossRef]
- Bergamaschi, M.; Cipolat-Gotet, C.; Cecchinato, A.; Schiavon, S.; Giovanni Bittante, G. Chemometric authentication of farming systems of origin of food (milk andripened cheese) using infrared spectra, fatty acid profiles, flavor, fingerprints, and sensory descriptions. Food Chem. 2020, 305, 125480. [Google Scholar] [CrossRef]
- Hedrington, M.S.; Davis, S.N. Sexual dimorphism in glucose andlipid metabolism during fasting, hypoglycemia, and exercise. Front. Endocrinol. 2015, 6, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aaslyng, M.D.; Jensen, H.; Karlsson, A.H. The gender background of texture attributes of pork loin. Meat Sci. 2018, 136, 79–84. [Google Scholar] [CrossRef]
- Liu, Y.N.; Albrecht, E.; Schering, L.; Kuehn, C.; Yang, R.J.; Zhao, Z.H.; Maak, S. Agouti signaling protein and itsreceptors as potential molecularmarkers for intramuscular and body fat deposition in cattle. Front. Physiol. 2018, 9, 172. [Google Scholar] [CrossRef] [PubMed]
- Cesar, A.S.M.; Regitano, L.C.A.; Poleti, M.D.; Andrade, S.C.S.; Tizioto, P.C.; Oliveira, P.S.N.; Felício, A.M.; Nascimento, M.L.D.; Chaves, A.S.; Lanna, D.P.D.; et al. Differences in the skeletal muscle transcriptome profile associated with extreme values of fatty acids content. BMC Genom. 2016, 17, 961. [Google Scholar] [CrossRef] [PubMed]
- Jiang, R.; Li, H.; Huang, Y.; Lan, X.; Lei, C.; Chen, H. Transcriptome profiling of lncRNA related to fat tissues of Qinchuan cattle. Gene 2020, 742, 144587. [Google Scholar] [CrossRef]
- Xu, K.; Ji, M.; Huang, X.; Peng, Y.; Wu, W.; Zhang, J. Differential Regulatory Roles of MicroRNAs in Porcine Intramuscular and Subcutaneous Adipocytes. J. Agric. Food Chem. 2020, 68, 3954–3962. [Google Scholar] [CrossRef] [PubMed]
- Piórkowska, K.; Żukowski, K.; Ropka-Molik, K.; Tyra, M.; Gurgul, A. A comprehensive transcriptome analysis of skeletal muscles in two Polish pig breeds differing in fat and meat quality traits. Genet. Mol. Biol. 2018, 41, 125–136. [Google Scholar] [CrossRef] [Green Version]
- Bazile, J.; Picard, B.; Chambon, C.; Valais, A.; Bonnet, M. Pathways and biomarkers of marbling and carcass fat deposition in bovine revealed by a combination of gel-based and gel-free proteomic analyses. Meat Sci. 2019, 156, 146–155. [Google Scholar] [CrossRef]
- Ma, C.; Wang, W.; Wang, Y.; Sun, Y.; Kang, L.; Zhang, Q.; Jiang, Y. TMT-labeled quantitative proteomic analyses on the longissimus dorsi to identify the proteins underlying intramuscular fat content in pigs. J. Proteom. 2020, 213, 103630. [Google Scholar] [CrossRef]
- Ministry of National Defense of the People’s Republic of China, Beijing, China. Technical Code of Practice for Pork Quality Assessment, China National Standard: NYT821-2019; Ministry of National Defense of the People’s Republic of China: Beijing, China, 2019. [Google Scholar]
- Song, S.Z.; Wu, J.P.; Zhao, S.G.; Casper, D.P.; He, B.; Liu, T.; Lang, X.; Gong, X.Y.; Liu, L.S. The effect of energy restriction on fatty acid profiles of longissimus dorsi and tissue adipose depots in sheep. J. Anim. Sci. 2017, 95, 3940–3948. [Google Scholar] [CrossRef]
- Popova, T.; Tejeda, L.; Peñarrieta, J.M.; Smith, M.A.; Bush, R.D.; Hopkins, D.L. Meat of South American camelids—Sensory quality and nutritional composition. Meat Sci. 2021, 171, 108285. [Google Scholar] [CrossRef]
- Zhang, H.; Wang, Q.; Fan, E. Stability Profile of Fatty Acids in Yak (Bos grunniens) Kidney Fat During the Initial Stages of Autoxidation. J. Am. Oil Chem. Soc. 2009, 86, 1057–1063. [Google Scholar] [CrossRef]
- Grundy, S.; Denke, M. Dietary influences on serum lipids and lipoproteins. J. Lipid Res. 1990, 31, 1149–1172. [Google Scholar] [CrossRef]
- Reddy, B.V.; Sivakumar, A.S.; Jeong, D.W.; Woo, Y.-B.; Park, S.-J.; Lee, S.-Y.; Byun, J.-Y.; Kim, C.-H.; Cho, S.-H.; Hwang, I. Beef quality traits of heifer in comparison with steer, bull and cow at various feeding environments. Anim. Sci. J. 2014, 86, 1–16. [Google Scholar] [CrossRef]
- Van Leeuwen, K.A.; Camin, F.; Jerónimo, E.; Vasta, V.; Prenzler, P.D.; Ryan, D.; Bessa, R.J.B. Dietary Effects on Stable Carbon Isotope Composition of Fatty Acids in Polar and Neutral Fractions of Intramuscular Fat of Lambs. J. Agric. Food Chem. 2017, 65, 9404–9411. [Google Scholar] [CrossRef]
- Wu, K.; Zhao, T.; Hogstrand, C.; Xu, Y.-C.; Ling, S.-C.; Chen, G.-H.; Luo, Z. FXR-mediated inhibition of autophagy contributes to FA-induced TG accumulation and accordingly reduces FA-induced lipotoxicity. Cell Commun. Signal. 2020, 18, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gómez, I.; Sarriés, M.V.; Ibañez, F.C.; Beriain, M.J. Quality Characteristics of a Low-Fat Beef Patty Enriched by Polyunsaturated Fatty Acids and Vitamin D3. J. Food Sci. 2018, 83, 454–463. [Google Scholar] [CrossRef] [PubMed]
- Enser, M.; Richardson, R.; Wood, J.; Gill, B.; Sheard, P. Feeding linseed to increase the n-3 PUFA of pork: Fatty acid composition of muscle, adipose tissue, liver and sausages. Meat Sci. 2000, 55, 201–212. [Google Scholar] [CrossRef]
- Li, P. Fatty acid composition assay and function evaluation of white yak meat. Food Sci. 2008, 29, 106–108. [Google Scholar]
- Jeong, J.; Kwon, E.G.; Im, S.K.; Seo, K.S.; Baik, M. Expression of fat deposition and fat removal genes is associated with intramuscular fat content in longissimus dorsi muscle of Korean cattle steers. J. Anim. Sci. 2012, 90, 2044–2053. [Google Scholar] [CrossRef]
- Dumbrepatil, A.B.; Zegalia, K.A.; Sajja, K.; Kennedy, R.T.; Marsh, E.N.G. Targeting viperin to the mitochondrion inhibits the thiolase activity of the trifunctional enzyme complex. J. Biol. Chem. 2020, 295, 2839–2849. [Google Scholar] [CrossRef]
- Jung, Y.H.; Bu, S.Y. Suppression of long chain acyl-CoA synthetase blocks intracellular fatty acid flux and glucose uptake in skeletal myotubes. BBA-Mol. Cell Biol. 2020, 1865, 158678. [Google Scholar] [CrossRef] [PubMed]
- Hiltunen, J.K.; Kastaniotis, A.J.; Autio, K.J.; Jiang, G.Y.; Chen, Z.J.; Glumoff, T. 17B-hydroxysteroid dehydrogenases as acyl thioester metabolizing enzymes. Mol. Cell. Endocrinol. 2019, 489, 107–118. [Google Scholar] [CrossRef]
- Sawai, M.; Uchida, Y.; Ohno, Y.; Miyamoto, M.; Nishioka, C.; Itohara, S.; Sassa, T.; Kihara, A. The 3-hydroxyacyl-CoA dehydratases HACD1 and HACD2 exhibit functional redundancy and are active in a wide range of fatty acid elongation pathways. J. Biol. Chem. 2017, 292, 15538–15551. [Google Scholar] [CrossRef] [Green Version]
- Krause, K.; Weiner, J.; Hönes, S.; Klöting, N.; Rijntjes, E.; Heiker, J.T.; Gebhardt, C.; Köhrle, J.; Führer, D.; Steinhoff, K.; et al. The Effects of Thyroid Hormones on Gene Expression of Acyl-Coenzyme A Thioesterases in Adipose Tissue and Liver of Mice. Eur. Thyroid. J. 2015, 4 (Suppl. 1), 59–66. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oliveira, D.M.; Chalfun-Junior, A.; Chizzotti, M.L.; Barreto, H.G.; Coelho, T.C.; Paiva, L.V.; Teixeira, P.D.; Schoonmaker, J.P.; Ladeira, M.M. Expression of genes involved in lipid metabolism in the muscle of beef cattle fed soybean or rumen-protected fat, with or without monensin supplementation. J. Anim. Sci. 2014, 92, 5426–5436. [Google Scholar] [CrossRef] [Green Version]
- Ko, M.H.; Puglielli, L. The sterol carrier protein SCP-x/pro-SCP-2 gene has transcriptional activity and regulates the alzheimer disease gamma-secretase. J. Biol. Chem. 2007, 282, 19742–19752. [Google Scholar] [CrossRef] [Green Version]
- Li, G.; Fu, S.; Chen, Y.; Jin, W.; Zhai, B.; Li, Y.; Sun, G.; Han, R.; Wang, Y.; Tian, Y.; et al. MicroRNA-15a Regulates the Differentiation of Intramuscular Preadipocytes by Targeting ACAA1, ACOX1 and SCP2 in Chickens. Int. J. Mol. Sci. 2019, 20, 4063. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anton, I.; Kovács, K.; Holló, G.; Farkas, V.; Lehel, L.; Hajda, Z.; Zsolnai, A. Effect of leptin, DGAT1 and TG gene polymorphisms on the intramuscular fat of Angus cattle in Hungary. Livest. Sci. 2011, 135, 300–303. [Google Scholar] [CrossRef]
- Mazzucco, J.P.; Goszczynski, D.; Ripoli, M.; Melucci, L.; Pardo, A.; Colatto, E.; Rogberg-Muñoz, A.; Mezzadra, C.; Depetris, G.; Giovambattista, G.; et al. Growth, carcass and meat quality traits in beef from Angus, Hereford and cross-breed grazing steers, and their association with SNPs in genes related to fat deposition metabolism. Meat Sci. 2016, 114, 121–129. [Google Scholar] [CrossRef] [PubMed]
- Gallego, S.F.; Sprenger, R.R.; Neess, D.; Pauling, J.K.; Færgeman, N.J.; Ejsing, C.S. Quantitative lipidomics reveals age-dependent perturbations of whole-body lipid metabolism in ACBP deficient mice. BBA-Mol. Cell Biol. L. 2017, 1862, 145–155. [Google Scholar] [CrossRef]
- Bowman, T.A.; O’Keeffe, K.R.; D’Aquila, T.; Yan, Q.W.; Griffin, J.D.; Killion, E.A. Acyl CoA synthetase 5 (ACSL5) ablation in mice increases energy expenditure and insulin sensitivity and delays fat absorption. Mol. Metab. 2016, 5, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Itabe, H.; Yamaguchi, T.; Nimura, S.; Sasabe, N. Perilipins: A diversity of intracellular lipid droplet proteins. Lipids Health Dis. 2017, 16, 83. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | FYs (Mean ± SE) | MYs (Mean ± SE) |
|---|---|---|
| a*45 min | 55.03 ± 0.72 | 53.27 ± 0.47 |
| b*45 min | 23.34 ± 0.84 | 21.31 ± 1.09 |
| L*45 min | 22.77 ± 0.65 a | 19.83 ± 0.74 b |
| a*24 h | 55.78 ± 0.55 | 53.88 ± 0.59 |
| b*24 h | 24.49 ± 0.87 a | 21.41 ± 0.55 b |
| L*24 h | 23.98 ± 1.04 b | 20.82 ± 0.29 a |
| pH45 min | 5.86 ± 0.03 | 5.82 ± 0.03 |
| pH24 h | 5.40 ± 0.05 | 5.55 ± 0.06 |
| Shear force (kg) | 7.93 ± 0.16 A | 9.29 ± 0.27 B |
| Marbling score | 3.25 ± 0.16 a | 2.75 ± 0.10 b |
| Cooking holding percentage (%) | 70.60 ± 0.86 | 71.37 ± 0.36 |
| FAs | Absolute Content (mg/100 g) | Relative Content (%) | ||
|---|---|---|---|---|
| FYs (Mean ± SE) | MYs (Mean ± SE) | FYs (Mean ± SE) | MYs (Mean ± SE) | |
| ΣSFAs | 1002.82 ± 14.94 A | 658.71 ± 15.74 B | 52.28 ± 0.33 A | 46.68 ± 0.43 B |
| C4:0 | 3.46 ± 0.22 A | 2.22 ± 0.09 B | 0.18 ± 0.01 | 0.16 ± 0.01 |
| C6:0 | 0.55 ± 0.04 A | 0.38 ± 0.01 B | 0.02 ± 0.001 | 0.03 ± 0.002 |
| C8:0 | 0.12 ± 0.02 A | 0.55 ± 0.02 B | 0.01 ± 0.001 A | 0.04 ± 0.00 B |
| C10:0 | 0.57 ± 0.03 a | 0.46 ± 0.02 b | 0.03 ± 0.001 | 0.03 ± 0.001 |
| C11:0 | 0.09 ± 0.01 | 0.07 ± 0.003 | 0.004 ± 0.0003 | 0.005 ± 0.0002 |
| C12:0 | 0.50 ± 0.04 | 0.39 ± 0.01 | 0.03 ± 0.002 | 0.03 ± 0.001 |
| C13:0 | 1.25 ±0.13 a | 0.83 ± 0.03 b | 0.06 ± 0.01 | 0.06 ± 0.002 |
| C14:0 | 9.86 ± 0.33 A | 8.03 ± 0.27 B | 0.515 ± 0.02 | 0.57 ± 0.02 |
| C15:0 | 6.92 ± 0.56 A | 9.42 ± 0.30 B | 0.36 ± 0.03 A | 0.67 ± 0.02 B |
| C16:0 | 271.66 ± 4.70 A | 181.69 ± 3.47 B | 14.18 ± 0.35 a | 12.88 ± 0.14 b |
| C17:0 | 15.52 ± 1.79 a | 23.04 ± 1.44 b | 0.81 ± 0.08 A | 1.63 ± 0.08 B |
| C18:0 | 581.96 ± 12.53 A | 341.49 ± 9.14 B | 30.33 ± 0.40 A | 24.20 ± 0.32 B |
| C20:0 | 2.01 ± 0.25 | 1.60 ± 0.09 | 0.10 ± 0.01 | 0.11 ± 0.01 |
| C21:0 | 6.48 ± 0.46 a | 4.95 ± 0.26 b | 0.34 ± 0.02 | 0.35 ± 0.01 |
| C22:0 | 2.81 ± 0.19 | 2.34 ± 0.21 | 0.15 ± 0.01 | 0.17 ± 0.01 |
| C23:0 | 11.66 ± 1.69 | 9.86 ± 0.28 | 0.61 ± 0.09 | 0.70 ± 0.02 |
| C24:0 | 87.41 ± 4.92 a | 71.41 ± 3.33 b | 4.55 ± 0.21 | 5.06 ± 0.19 |
| ΣMUFAs | 536.21 ± 10.36 A | 423.15 ± 5.61 B | 27.95 ± 0.36 A | 30.02 ± 0.24 B |
| C14:1 | 1.08 ± 0.11 A | 18.29 ± 1.22 B | 0.06 ± 0.01 A | 1.30 ± 0.08 B |
| cis-C15:1 | 2.91 ±0.28 A | 5.10 ± 0.26 B | 0.15 ± 0.01 A | 0.36 ± 0.02 B |
| C16:1 | 36.20 ± 3.59 | 37.01 ± 1.50 | 1.88 ± 0.17 A | 2.62 ± 0.10 B |
| cis-C17:1 | 5.62 ± 0.70 A | 15.23 ±0.89 B | 0.29 ± 0.04 A | 1.08 ± 0.05 B |
| cis-C18:1 | 424.98 ± 7.31 A | 300.62 ± 4.82 B | 22.16 ± 0.32 | 21.33 ± 0.26 |
| trans-C18:1 | 11.24 ± 1.14 A | 6.08 ± 0.55 B | 0.59 ± 0.06 | 0.43 ± 0.04 |
| cis-C20:1 | 4.97 ± 0.44 A | 1.79 ± 0.05 B | 0.26 ± 0.02 A | 0.13 ± 0.004 B |
| C24:1 | 49.23 ± 3.07 a | 39.04 ± 1.04 b | 2.56 ± 0.14 | 2.77 ± 0.09 |
| ΣPUFAs | 379.29 ± 6.90 A | 328.71 ± 7.27 B | 19.77 ± 0.13 A | 23.30 ± 0.32 B |
| cis-C18:2n6 | 274.54 ± 7.37 A | 173.92 ± 4.58 B | 14.31 ± 0.30 | 12.33 ± 0.23 |
| trans-C18:2n6 | 0.53 ± 0.06 | 0.47 ± 0.04 | 0.03 ± 0.01 | 0.03 ± 0.01 |
| cis-C18:3n6 | 5.33 ± 0.52 A | 10.71 ± 0.27 B | 0.28 ± 0.03 A | 0.76 ± 0.02 B |
| C18:3n3 | 16.83 ± 2.48 A | 47.95 ± 2.28 B | 0.87 ± 0.12 A | 3.39 ± 0.11 B |
| cis-C20:2 | 3.17 ± 0.50 A | 6.21 ± 0.32 B | 0.16 ± 0.02 A | 0.44 ± 0.02 B |
| C20:4n6 | 54.56 ± 2.05 A | 43.61 ± 1.62 B | 2.84 ± 0.10 | 3.10 ± 0.13 |
| cis-C20:3n3 | 0.25 ± 0.04 | 0.25 ± 0.01 | 0.01 ± 0.002 | 0.02 ± 0.001 |
| cis-C20:5n3 | 21.57 ± 2.20 A | 43.28 ± 1.09 B | 1.13 ± 0.12 A | 3.07 ± 0.10 B |
| cis-C22:6n3 | 2.51 ± 0.23 | 2.31 ± 0.07 | 0.13 ± 0.01 a | 0.16 ± 0.01 b |
| ΣUFAs | 915.50 ± 15.66 A | 751.86 ± 12.14 B | 47.72 ± 0.33 A | 53.32 ± 0.43 B |
| FAs | FYs (Mean ± SE) | MYs (Mean ± SE) |
|---|---|---|
| Σn-3 PUFAs (%) | 2.15 ± 0.18 A | 6.65 ± 0.07 B |
| Σn-6 PUFAs (%) | 16.22 ± 0.21 a | 17.46 ± 0.34 b |
| Σn-6/Σn-3 PUFAs | 8.53 ±0.86 A | 2.44 ± 0.06 B |
| ΣSFAs/ΣUFAs | 1.10 ± 0.01 A | 0.88 ± 0.02 B |
| ΣMUFAs/ΣPUFAs | 1.41 ± 0.02 A | 1.29 ± 0.02 B |
| ΣPUFAs/ΣSFAs | 0.38 ± 0.01 A | 0.50 ± 0.01 B |
| KEGG Pathway | DEGs | DEPs |
|---|---|---|
| Biosynthesis of unsaturated fatty acids (bom01040) | SCD, ACOT7, HACD2, SCP2, ELOVL6, HSD17B12, ACAA1, HSD17B12 | SCD, ELOVL6, HACD3 |
| Fatty acid elongation (bom00062) | ACOT7, HACD2, THEM4, HADHB, ELOVL6, HADH, HSD17B12 | ELOVL6, HADH, HACD3 |
| Terpenoid backbone biosynthesis (bom00900) | PCYOX1, ACAT2, FNTB | PCYOX1, ACAT2 |
| Butanoate metabolism (bom00650) | ACSM1, HADH, BDH1, ACAT2 | ACSM1, HADH, ACAT2 |
| Fatty acid degradation (bom00071) | ACADSB, HADHB, HADH, ACAT2, ACAA1, CPT1C | ACSL5, HADH, ACAT2 |
| Tryptophan metabolism (bom00380) | MAOB, AADAT, HADH, ACMSD, ACAT2 | HADH, ACAT2 |
| Synthesis and degradation of ketone bodies (Bom00072) | BDH1, ACAT2 | ACAT2 |
| PPAR signaling pathway (bom03320) | SCD, SCP2, PLIN5, ACOX2, ACAA1, LPL, ME1, CPT1C, SLC27A4, SLC27A6, DBI, SCP2 | ACSL5, SCD, PLIN4, FABP1, PLIN2, LPL, ME1, DBI |
| Pyruvate metabolism (bom00620) | ACAT2, ACSS2, LDHA, ME1, SCP2, ACYP2 | ACAT2, ME1 |
| Gender | Variable | L*45 min | b*24 h | L*24 h | Shear Force | Marbing |
|---|---|---|---|---|---|---|
| MYs | Fat content | 0.41 | 0.79 | 0.89 a,b | −0.84 a,b | 0.85 a,b |
| C18:0 | 0.43 | 0.87 a,b | 0.85 a,b | −0.92 A,B | 0.76 | |
| cis-C18:1 | 0.39 | 0.78 | 0.90 a,b | −0.81 a,b | 0.91 a,b | |
| cis-C18:2 | 0.15 | 0.31 | 0.44 | −0.32 | 0.39 | |
| C16:0 | 0.42 | 0.58 | 0.52 | −0.61 | 0.35 | |
| C20:4n6 | 0.23 | −0.25 | −0.21 | −0.30 | −0.28 | |
| C24:0 | 0.29 | 0.77 | 0.85 a,b | −0.81 a,b | 0.88 a,b | |
| C24:1 | −0.27 | −0.55 | −0.28 | −0.56 | −0.17 | |
| cis-C20:5n3 | 0.56 | −0.06 | 0.01 | −0.12 | −0.07 | |
| FYs | Fat content | 0.94 A,B | 0.88 a,b | 0.93 A,B | −0.84 a,b | 0.66 |
| C18:0 | 0.87 a,b | 0.92 A,B | 0.70 | −0.93 A,B | 0.72 | |
| cis-C18:1 | 0.54 | 0.34 | 0.72 | −0.51 | 0.48 | |
| cis-C18:2 | 0.79 | 0.73 | 0.68 | −0.94 A,B | 0.81 a,b | |
| C16:0 | −0.14 | −0.14 | −0.11 | −0.07 | −0.03 | |
| C20:4n6 | 0.21 | 0.04 | 0.44 | −0.40 | 0.01 | |
| C24:0 | 0.53 | 0.46 | 0.54 | −0.23 | 0.17 | |
| C24:1 | 0.35 | 0.52 | 0.45 | −0.23 | −0.07 | |
| cis-C20:5n3 | −0.57 | −0.71 | −0.53 | −0.76 | −0.38 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xiong, L.; Pei, J.; Chu, M.; Wu, X.; Kalwar, Q.; Yan, P.; Guo, X. Fat Deposition in the Muscle of Female and Male Yak and the Correlation of Yak Meat Quality with Fat. Animals 2021, 11, 2142. https://doi.org/10.3390/ani11072142
Xiong L, Pei J, Chu M, Wu X, Kalwar Q, Yan P, Guo X. Fat Deposition in the Muscle of Female and Male Yak and the Correlation of Yak Meat Quality with Fat. Animals. 2021; 11(7):2142. https://doi.org/10.3390/ani11072142
Chicago/Turabian StyleXiong, Lin, Jie Pei, Min Chu, Xiaoyun Wu, Qudratullah Kalwar, Ping Yan, and Xian Guo. 2021. "Fat Deposition in the Muscle of Female and Male Yak and the Correlation of Yak Meat Quality with Fat" Animals 11, no. 7: 2142. https://doi.org/10.3390/ani11072142
APA StyleXiong, L., Pei, J., Chu, M., Wu, X., Kalwar, Q., Yan, P., & Guo, X. (2021). Fat Deposition in the Muscle of Female and Male Yak and the Correlation of Yak Meat Quality with Fat. Animals, 11(7), 2142. https://doi.org/10.3390/ani11072142