
Impact of Estrogens Present in Environment on Health and Welfare of Animals

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Invertebrates
3. Fish
4. Amphibians
5. Reptiles
6. Birds
7. Mammals
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Hammes, S.R.; Levin, E.R. Impact of estrogens in males and androgens in females. J. Clin. Investig. 2019, 129, 1818–1826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boonyaratanakornkit, V.; Edwards, D.P. Receptor Mechanisms Mediating Non-Genomic Actions of Sex Steroids. In Seminars in Reproductive Medicine; Thieme Medical Publishers: New York, NY, USA, 2007; Volume 25, pp. 139–153. [Google Scholar]
- Rechsteiner, D.; Schrade, S.; Zähner, M.; Müller, M.; Hollender, J.; Bucheli, T.D. Occurrence and Fate of Natural Estrogens in Swiss Cattle and Pig Slurry. J. Agric. Food Chem. 2020, 68, 5545–5554. [Google Scholar] [CrossRef] [PubMed]
- Yang, S.; Yu, W.; Yang, L.; Du, B.; Chen, S.; Sun, W.; Jiang, H.; Xie, M.; Tang, J. Occurrence and Fate of Steroid Estrogens in a Chinese Typical Concentrated Dairy Farm and Slurry Irrigated Soil. J. Agric. Food Chem. 2021, 69, 67–77. [Google Scholar] [CrossRef]
- Adeel, M.; Song, X.; Wang, Y.; Francis, D.; Yang, Y. Environmental impact of estrogens on human, animal and plant life: A critical review. Environ. Int. 2017, 99, 107–119. [Google Scholar] [CrossRef]
- Huang, Y.; Li, W.; Qin, L.; Xie, X.; Gao, B.; Sun, J.; Li, A. Distribution of endocrine-disrupting chemicals in colloidal and soluble phases in municipal secondary effluents and their removal by different advanced treatment processes. Chemosphere 2019, 219, 730–739. [Google Scholar] [CrossRef] [PubMed]
- Tran, N.H.; Reinhard, M.; Gin, K.Y.-H. Occurrence and fate of emerging contaminants in municipal wastewater treatment plants from different geographical regions—A review. Water Res. 2018, 133, 182–207. [Google Scholar] [CrossRef]
- Silva, L.G.R.; Costa, E.P.; Starling, M.C.V.M.; dos Santos Azevedo, T.; Bottrel, S.E.C.; Pereira, R.O.; Sanson, A.L.; Afonso, R.J.C.F.; Amorim, C.C. LED irradiated photo-Fenton for the removal of estrogenic activity and endocrine disruptors from wastewater treatment plant effluent. Environ. Sci. Pollut. Res. 2021, 28, 24067–24078. [Google Scholar] [CrossRef]
- Aris, A.Z.; Shamsuddin, A.S.; Praveena, S.M. Occurrence of 17α-ethynylestradiol (EE2) in the environment and effect on ex-posed biota: A review. Environ. Int. 2014, 69, 104–119. [Google Scholar] [CrossRef]
- Shore, L.S.; Shemesh, M. Estrogen as an Environmental Pollutant. Bull. Environ. Contam. Toxicol. 2016, 97, 447–448. [Google Scholar] [CrossRef] [Green Version]
- Müller, A.-K.; Markert, N.; Leser, K.; Kämpfer, D.; Crawford, S.E.; Schäffer, A.; Segner, H.; Hollert, H. Assessing endocrine disruption in freshwater fish species from a “hotspot” for estrogenic activity in sediment. Environ. Pollut. 2020, 257, 113636. [Google Scholar] [CrossRef]
- Abdellah, Y.A.Y.; Zang, H.; Li, C. Steroidal Estrogens During Composting of Animal Manure: Persistence, Degradation, and Fate, a Review. Water Air Soil Pollut. 2020, 231, 1–19. [Google Scholar] [CrossRef]
- Pratush, A.; Ye, X.; Yang, Q.; Kan, J.; Peng, T.; Wang, H.; Huang, T.; Xiong, G.; Hu, Z. Biotransformation strategies for steroid estrogen and androgen pollution. Appl. Microbiol. Biotechnol. 2020, 104, 2385–2409. [Google Scholar] [CrossRef]
- Almeida, Â.; Silva, M.G.; Soares, A.M.V.M.; Freitas, R. Concentrations levels and effects of 17alpha-Ethinylestradiol in fresh-water and marine waters and bivalves: A review. Environ. Res. 2020, 185, 109316. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Hain, E.; Timm, A.; Blaney, L. Bioaccumulation of estrogenic hormones and UV-filters in red swamp crayfish (Pro-cambarus clarkii). Sci. Total Environ. 2021, 764, 142871. [Google Scholar] [CrossRef]
- Amenyogbe, E.; Chen, G.; Wang, Z.; Lu, X.; Lin, M.; Lin, A.Y. A Review on Sex Steroid Hormone Estrogen Receptors in Mammals and Fish. Int. J. Endocrinol. 2020, 2020, 1–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Henderson, B.E.; Feigelson, H.S. Hormonal carcinogenesis. Carcinogenesis 2000, 21, 427–433. [Google Scholar] [CrossRef] [PubMed]
- Bohra, A.; Bhateja, S. Carcinogenesis and sex hormones: A review. Endocrinol. Metab. Syndr. 2015, 4, 1–14. [Google Scholar]
- Ford, A.T.; Leblanc, G.A. Endocrine Disruption in Invertebrates: A Survey of Research Progress. Environ. Sci. Technol. 2020, 54, 13365–13369. [Google Scholar] [CrossRef]
- Castro, L.F.C.; Santos, M.M. To bind or not to bind: The taxonomic scope of nuclear receptor mediated endocrine disruption in invertebrate phyla. Environ. Sci. Technol. 2014, 48, 5361–5363. [Google Scholar] [CrossRef]
- Bovier, T.F.; Rossi, S.; Mita, D.G.; Digilio, F.A. Effects of the synthetic estrogen 17-α-ethinylestradiol on Drosophila melanogaster: Dose and gender dependence. Ecotoxicol. Environ. Saf. 2018, 162, 625–632. [Google Scholar] [CrossRef]
- Segner, H.; Caroll, K.; Fenske, M.; Janssen, C.; Maack, G.; Pascoe, D.; Schäfers, C.; Vandenbergh, G.; Watts, M.; Wenzel, A. Identification of endocrine-disrupting effects in aquatic vertebrates and invertebrates: Report from the European IDEA project. Ecotoxicol. Environ. Saf. 2003, 54, 302–314. [Google Scholar] [CrossRef]
- Islam, R.; Yu, R.M.K.; O’Connor, W.A.; Tran, T.K.A.; Andrew-Priestley, M.; Leusch, F.D.; MacFarlane, G.R. Parental exposure to the synthetic estrogen 17α-ethinylestradiol (EE2) affects offspring development in the Sydney rock oyster, Saccostrea glomerata. Environ. Pollut. 2020, 266, 114994. [Google Scholar] [CrossRef]
- Ciocan, C.; Leon, E.C.; Puinean, A.M.; Hill, E.M.; Minier, C.; Osada, M.; Fenlon, K.; Rotchell, J.M. Effects of estrogen exposure in mussels, Mytilus edulis, at different stages of gametogenesis. Environ. Pollut. 2010, 158, 2977–2984. [Google Scholar] [CrossRef]
- Fodor, I.; Urbán, P.; Scott, A.P.; Pirger, Z. A critical evaluation of some of the recent so-called ‘evidence’ for the involvement of vertebrate-type sex steroids in the reproduction of mollusks. Mol. Cell. Endocrinol. 2020, 516, 110949. [Google Scholar] [CrossRef] [PubMed]
- Clubbs, R.L.; Brooks, B.W. Daphnia magna responses to a vertebrate estrogen receptor agonist and an antagonist: A multi-generational study. Ecotoxicol. Environ. Saf. 2007, 67, 385–398. [Google Scholar] [CrossRef] [PubMed]
- Fernández-González, L.E.; Diz, A.P.; Grueiro, N.G.; Muniategui-Lorenzo, S.; Beiras, R.; Sánchez-Marín, P. No evidence that vitellogenin protein expression is induced in marine mussels after exposure to an estrogenic chemical. Sci. Total Environ. 2020, 721, 137638. [Google Scholar] [CrossRef]
- Souza, M.S.; Hallgren, P.; Balseiro, E.; Hansson, L.-A. Low concentrations, potential ecological consequences: Synthetic estro-gens alter life-history and demographic structures of aquatic invertebrates. Environ. Pollut. 2013, 178, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Marcial, H.S.; Hagiwara, A.; Snell, T.W. Estrogenic compounds affect development of harpacticoid copepod Tigriopus japonicus. Environ. Toxicol. Chem. 2003, 22, 3025–3030. [Google Scholar] [CrossRef]
- Fernandez, M.A. Populations Collapses in Marine Invertebrates Due to Endocrine Disruption: A Cause for Concern? Front. Endocrinol. 2019, 10, 721. [Google Scholar] [CrossRef] [PubMed]
- Katsiadaki, I. Are marine invertebrates really at risk from endocrine-disrupting chemicals? Curr. Opin. Environ. Sci. Health 2019, 11, 37–42. [Google Scholar] [CrossRef]
- Zubizarreta, L.; Silva, A.C.; Quintana, L. The estrogenic pathway modulates non-breeding female aggression in a teleost fish. Physiol. Behav. 2020, 220, 112883. [Google Scholar] [CrossRef] [PubMed]
- Cabas, I.; Chaves-Pozo, E.; Mulero, V.; García-Ayala, A. Role of estrogens in fish immunity with special emphasis on GPER1. Dev. Comp. Immunol. 2018, 89, 102–110. [Google Scholar] [CrossRef] [PubMed]
- Szwejser, E.; Kemenade, B.L.V.-V.; Maciuszek, M.; Chadzinska, M. Estrogen-dependent seasonal adaptations in the immune response of fish. Horm. Behav. 2017, 88, 15–24. [Google Scholar] [CrossRef]
- Müller, A.-K.; Markert, N.; Leser, K.; Kämpfer, D.; Schiwy, S.; Riegraf, C.; Buchinger, S.; Gan, L.; Abdallah, A.T.; Denecke, B.; et al. Bioavailability and impacts of estrogenic compounds from suspended sediment on rainbow trout (Oncorhynchus mykiss). Aquat. Toxicol. 2021, 231, 105719. [Google Scholar] [CrossRef] [PubMed]
- Thrupp, T.J.; Runnalls, T.J.; Scholze, M.; Kugathas, S.; Kortenkamp, A.; Sumpter, J.P. The consequences of exposure to mixtures of chemicals: Something from ‘nothing’ and ‘a lot from a little’ when fish are exposed to steroid hormones. Sci. Total Environ. 2018, 619, 1482–1492. [Google Scholar] [CrossRef] [PubMed]
- Filby, A.L.; Thorpe, K.L.; Maack, G.; Tyler, C.R. Gene expression profiles revealing the mechanisms of anti-androgen-and estrogen-induced feminization in fish. Aquat. Toxicol. 2017, 81, 219–231. [Google Scholar] [CrossRef]
- Golshan, M.; Alavi, S.M.H. Androgen signaling in male fishes: Examples of anti-androgenic chemicals that cause reproductive disorders. Theriogenology 2019, 139, 58–71. [Google Scholar] [CrossRef] [PubMed]
- Kinnberg, K.; Toft, G. Effects of estrogenic and antiandrogenic compounds on the testis structure of the adult guppy (Poecilia reticulata). Ecotoxicol. Environ. Saf. 2003, 54, 16–24. [Google Scholar] [CrossRef]
- Green, C.; Brian, J.; Kanda, R.; Scholze, M.; Williams, R.; Jobling, S. Environmental concentrations of anti-androgenic pharmaceuticals do not impact sexual disruption in fish alone or in combination with steroid oestrogens. Aquat. Toxicol. 2015, 160, 117–127. [Google Scholar] [CrossRef]
- Cox, M.; Peterson, K.; Tan, D.; Novak, P.; Schoenfuss, H.; Ward, J. Temperature modulates estrone degradation and biological effects of exposure in fathead minnows. Sci. Total Environ. 2018, 621, 1591–1600. [Google Scholar] [CrossRef] [Green Version]
- Korsgaard, B.; Mommsen, T.P.; Saunders, R.L. The effect of temperature on the vitellogenic response in Atlantic salmon post-smolts (Salmo salar). Gen. Comp. Endocrinol. 1986, 62, 193–201. [Google Scholar] [CrossRef]
- Jackson, L.; Klerks, P. Effects of the synthetic estrogen 17α-ethinylestradiol on Heterandria formosa populations: Does matrotrophy circumvent population collapse? Aquat. Toxicol. 2020, 229, 105659. [Google Scholar] [CrossRef]
- Karki, N.P.; Colombo, R.E.; Gaines, K.F.; Maia, A. Exposure to 17β estradiol causes erosion of sexual dimorphism in Bluegill (Lepomis macrochirus). Environ. Sci. Pollut. Res. 2021, 28, 6450–6458. [Google Scholar] [CrossRef]
- Sumpter, J.P. Feminized responses in fish to environmental estrogens. Toxicol. Lett. 1995, 82–83, 737–742. [Google Scholar] [CrossRef]
- Martinez-Bengochea, A.; Doretto, L.; Rosa, I.; de Oliveira, M.A.; Silva, C.; Silva, D.; Santos, G.; Santos, J.; Avelar, M.; Silva, L.; et al. Effects of 17β-estradiol on early gonadal development and expression of genes implicated in sexual differentiation of a South American teleost, Astyanax altiparanae. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2020, 248–249, 110467. [Google Scholar] [CrossRef]
- Filby, A.L.; Shears, J.A.; Drage, B.E.; Churchley, J.H.; Tyler, C.R. Effects of Advanced Treatments of Wastewater Effluents on Estrogenic and Reproductive Health Impacts in Fish. Environ. Sci. Technol. 2010, 44, 4348–4354. [Google Scholar] [CrossRef]
- Dang, Z.; Kienzler, A. Changes in fish sex ratio as a basis for regulating endocrine disruptors. Environ. Int. 2019, 130, 104928. [Google Scholar] [CrossRef] [PubMed]
- Hill, R.L., Jr.; Janz, D.M. Developmental estrogenic exposure in zebrafish (Danio rerio): I Effects on sex ratio and breeding success. Aquat. Toxicol. 2003, 63, 417–429. [Google Scholar] [CrossRef]
- Vajda, A.M.; Barber, L.B.; Gray, J.L.; Lopez, E.M.; Woodling, J.D.; Norris, D.O. Reproductive disruption in fish downstream from an estrogenic wastewater effluent. Environ. Sci. Technol. 2008, 42, 3407–3414. [Google Scholar] [CrossRef]
- Teta, C.; Holbech, B.; Norrgren, L.; Naik, Y.; Teta, C.; Holbech, B.; Norrgren, L.; Naik, Y. Occurrence of oestrogenic pollutants and widespread feminisation of male tilapia in peri-urban dams in Bulawayo, Zimbabwe. Afr. J. Aquat. Sci. 2018, 43, 17–26. [Google Scholar] [CrossRef]
- Thorpe, K.; Maack, G.; Benstead, R.; Tyler, C.R. Estrogenic Wastewater Treatment Works Effluents Reduce Egg Production in Fish. Environ. Sci. Technol. 2009, 43, 2976–2982. [Google Scholar] [CrossRef]
- Gutjahr-Gobell, R.E.; Zaroogian, G.E.; Horowitz, D.J.B.; Gleason, T.R.; Mills, L.J. Individual effects of estrogens on a marine fish, Cunner (Tautogolabrus adspersus), extrapolated to the population level. Ecotoxicol. Environ. Saf. 2006, 63, 244–252. [Google Scholar] [CrossRef]
- Brian, J.V.; Harris, C.A.; Scholze, M.; Kortenkamp, A.; Booy, P.; Lamoree, M.; Pojana, G.; Jonkers, N.; Marcomini, A.A.; Sumpter, J.P. Evidence of Estrogenic Mixture Effects on the Reproductive Performance of Fish. Environ. Sci. Technol. 2007, 41, 337–344. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schäfers, C.; Teigeler, M.; Wenzel, A.; Maack, G.; Fenske, M.; Segner, H. Concentration—And Time—Dependent Effects of the Synthetic Estrogen, 17α-ethinylestradiol, on Reproductive Capabilities of the Zebrafish, Danio rerio. J. Toxicol. Environ. Health Part A 2007, 70, 768–779. [Google Scholar] [CrossRef]
- Vajda, A.M.; Barber, L.B.; Gray, J.L.; Lopez, E.M.; Bolden, A.M.; Schoenfuss, H.; Norris, D.O. Demasculinization of male fish by wastewater treatment plant effluent. Aquat. Toxicol. 2011, 103, 213–221. [Google Scholar] [CrossRef] [PubMed]
- Jones, P.; De Coen, W.M.; Tremblay, L.; Giesy, J.P. Vitellogenin as a biomarker for environmental estrogens. Water Sci. Technol. 2000, 42, 1–14. [Google Scholar] [CrossRef]
- Jobling, S.; Casey, D.; Rodgers-Gray, T.; Oehlmann, J.; Schulte-Oehlmann, U.; Pawlowski, S.; Baunbeck, T.; Turner, A.; Tyler, C. Comparative responses of molluscs and fish to environmental estrogens and an estrogenic effluent. Aquat. Toxicol. 2003, 65, 205–220. [Google Scholar] [CrossRef]
- Song, J.; Nagae, M.; Takao, Y.; Soyano, K. Field survey of environmental estrogen pollution in the coastal area of Tokyo Bay and Nagasaki City using the Japanese common goby Acanthogobius flavimanus. Environ. Pollut. 2020, 258, 113673. [Google Scholar] [CrossRef]
- Ward, J.L.; Korn, V.; Auxier, A.N.; Schoenfuss, H.L. Temperature and Estrogen Alter Predator—Prey Interactions between Fish Species. Integr. Org. Biol. 2020, 2, obaa008. [Google Scholar] [CrossRef] [Green Version]
- Wu, C.-C.; Shields, J.N.; Akemann, C.; Meyer, D.N.; Connell, M.; Baker, B.B.; Pitts, D.K.; Baker, T.R. The phenotypic and transcriptomic effects of developmental exposure to nanomolar levels of estrone and bisphenol A in zebrafish. Sci. Total Environ. 2021, 757, 143736. [Google Scholar] [CrossRef]
- Petersen, K.; Tollefsen, K.E. Assessing combined toxicity of estrogen receptor agonists in a primary culture of rainbow trout (Oncorhynchus mykiss) hepatocytes. Aquat. Toxicol. 2011, 101, 186–195. [Google Scholar] [CrossRef]
- Kidd, K.A.; Blanchfield, P.J.; Mills, K.H.; Palace, V.P.; Evans, R.E.; Lazorchak, J.M.; Flick, R.W. Collapse of a fish population after exposure to a synthetic estrogen. Proc. Natl. Acad. Sci. USA 2007, 104, 8897–8901. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jobling, S.; Williams, R.; Johnson, A.; Taylor, A.; Gross-Sorokin, M.; Nolan, M.; Tyler, C.R.; van Aerle, R.; Santos, E.; Brighty, G. Predicted Exposures to Steroid Estrogens in U.K. Rivers Correlate with Widespread Sexual Disruption in Wild Fish Populations. Environ. Health Perspect. 2006, 114, 32–39. [Google Scholar] [CrossRef] [Green Version]
- Hicks, K.A.; Fuzzen, M.L.M.; McCann, E.K.; Arlos, M.J.; Bragg, L.M.; Kleywegt, S.; Tetreault, G.R.; McMaster, M.E.; Servos, M.R. Reduction of Intersex in a Wild Fish Population in Response to Major Municipal Wastewater Treatment Plant Upgrades. Environ. Sci. Technol. 2017, 51, 1811–1819. [Google Scholar] [CrossRef]
- Wang, C.; An, L.; Wu, S.; Jia, A.; Sun, J.; Huang, C.; Mu, D.; Hu, J. Potential Link between Equol Pollution and Field-Observed Intersex in Wild So-iuy Mullets (Mugil soiuy). Environ. Sci. Technol. 2020, 54, 12393–12401. [Google Scholar] [CrossRef] [PubMed]
- Munakata, A. Migratory Behaviors in Masu Salmon (Oncorhynchus masou) and the Influence of Endocrinological Factors. Aqua-BioSci. Monogr. 2012, 5, 29–65. [Google Scholar] [CrossRef] [Green Version]
- Munakata, A.; Amano, M.; Ikuta, K.; Kitamura, S.; Aida, K. Involvement of sex steroids and thyroid hormones in upstream and downstream behaviors in masu salmon, Oncorhynchus masou. Aquaculture 2012, 362–363, 158–166. [Google Scholar] [CrossRef]
- Madsen, S.S.; Skovbølling, S.; Nielsen, C.; Korsgaard, B. 17-β Estradiol and 4-nonylphenol delay smolt development and downstream migration in Atlantic salmon, Salmo salar. Aquat. Toxicol. 2004, 68, 109–120. [Google Scholar] [CrossRef]
- Bangsgaard, K.; Madsen, S.S.; Korsgaard, B. Effect of waterborne exposure to 4-tert-octylphenol and 17β-estradiol on smolti-fication and downstream migration in Atlantic salmon, Salmo salar. Aquat. Toxicol. 2006, 80, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Williams, R.J.; Keller, V.D.; Johnson, A.C.; Young, A.R.; Holmes, M.G.; Wells, C.; Gross-Sorokin, M.; Benstead, R. A national risk assessment for intersex in fish arising from steroid estrogens. Environ. Toxicol. Chem. 2009, 28, 220–230. [Google Scholar] [CrossRef]
- Johnson, A.C.; Chen, Y. Does exposure to domestic wastewater effluent (including steroid estrogens) harm fish populations in the UK? Sci. Total Environ. 2017, 589, 89–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hoffmann, F.; Kloas, W. Estrogens Can Disrupt Amphibian Mating Behavior. PLoS ONE 2012, 7, e32097. [Google Scholar] [CrossRef] [Green Version]
- Lambert, M.R.; Giller, G.S.J.; Barber, L.B.; Fitzgerald, K.C.; Skelly, D.K. Suburbanization, estrogen contamination, and sex ratio in wild amphibian populations. Proc. Natl. Acad. Sci. USA 2015, 112, 11881–11886. [Google Scholar] [CrossRef] [Green Version]
- Park, B.J.; Kidd, K. Effects of the synthetic estrogen ethinylestradiol on early life stages of mink frogs and green frogs in the wild and in situ. Environ. Toxicol. Chem. 2005, 24, 2027–2036. [Google Scholar] [CrossRef]
- Hayes, T.B.; Case, P.; Chui, S.; Chung, D.; Haeffele, C.; Haston, K.; Lee, M.; Mai, V.P.; Marjuoa, Y.; Parker, J.; et al. Pesticide Mixtures, Endocrine Disruption, and Amphibian Declines: Are We Underestimating the Impact? Environ. Health Perspect. 2006, 114, 40–50. [Google Scholar] [CrossRef]
- Ghali, K.; Leuenberger, J.; Ansermet, M.; Perrin, N.; Dufresnes, C. Toxic effects of estradiol E2 on development in the European tree frog (Hyla arborea). Herpetol. Notes 2016, 9, 249–253. [Google Scholar]
- Wilczynski, W.; Lynch, K.S. Female sexual arousal in amphibians. Horm. Behav. 2011, 59, 630–636. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kikuyama, S.; Yamamoto, K.; Iwata, T.; Toyoda, F. Peptide and protein pheromones in amphibians. Comp. Biochem. Part B Biochem. Mol. Biol. 2002, 132, 69–74. [Google Scholar] [CrossRef]
- Kloas, W.; Lutz, I.; Einspanier, R. Amphibians as a model to study endocrine disruptors: II Estrogenic activity of environmental chemicals in vitro and in vivo. Sci. Total Environ. 1999, 225, 59–68. [Google Scholar] [CrossRef]
- Mosconi, G.; Carnevali, O.; Franzoni, M.; Cottone, E.; Lutz, I.; Kloas, W.; Yamamoto, K.; Kikuyama, S.; Polzonetti-Magni, A. Environmental Estrogens and Reproductive Biology in Amphibians. Gen. Comp. Endocrinol. 2002, 126, 125–129. [Google Scholar] [CrossRef] [PubMed]
- Fenske, M.; Maack, G.; Schäfers, C.; Segner, H. An environmentally relevant concentration of estrogen induces arrest of male gonad development in zebrafish, Danio rerio. Environ. Toxicol. Chem. 2005, 24, 1088–1098. [Google Scholar] [CrossRef]
- Qin, G.; Zhang, Y.; Zhang, B.; Zhang, Y.; Liu, Y.; Lin, Q. Environmental estrogens and progestins disturb testis and brood pouch development with modifying transcriptomes in male-pregnancy lined seahorse Hippocampus erectus. Sci. Total Environ. 2020, 715, 136840. [Google Scholar] [CrossRef] [PubMed]
- Miller, D.H.; Jensen, K.M.; Villeneuve, D.L.; Kahl, M.D.; Makynen, E.A.; Durhan, E.J.; Ankley, G.T. Linkage of biochemical responses to population-level effects: A case study with vitellogenin in the fathead minnow (Pimephales promelas). Environ. Toxicol. Chem. 2007, 26, 521–527. [Google Scholar] [CrossRef]
- Falfushynska, H.I.; Gnatyshyna, L.L.; Horyn, O.; Stoliar, O.B. Vulnerability of marsh frog Pelophylax ridibundus to the typical wastewater effluents ibuprofen, triclosan and estrone, detected by multi-biomarker approach. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2017, 202, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Bögi, C.; Levy, G.; Lutz, I.; Kloas, W. Functional genomics and sexual differentiation in amphibians. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 133, 559–570. [Google Scholar] [CrossRef]
- Li, Y.; Li, J.; Shen, Y.; Xiong, Y.; Li, X.; Qin, Z. Identification of estrogen receptor target genes involved in gonadal feminization caused by estrogen in Xenopus laevis. Aquat. Toxicol. 2021, 232, 105760. [Google Scholar] [CrossRef] [PubMed]
- Croteau, M.C.; Hogan, N.; Gibson, J.C.; Lean, D.; Trudeau, V.L. Toxicological threats to amphibians and reptiles in urban environments. Urban Herpetol. 2008, 3, 197–209. [Google Scholar]
- Matthews, J.; Celius, T.; Halgren, R.; Zacharewski, T. Differential estrogen receptor binding of estrogenic substances: A species comparison. J. Steroid Biochem. Mol. Biol. 2000, 74, 223–234. [Google Scholar] [CrossRef]
- Campbell, A.K.; Wann, K.; Matthews, S.B. Lactose causes heart arrhythmia in the water flea Daphnia pulex. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2004, 139, 225–234. [Google Scholar] [CrossRef]
- Palmer, B.D.; Palmer, S.K. Vitellogenin induction by xenobiotic estrogens in the red-eared turtle and African clawed frog. Environ. Health Perspect. 1995, 103, 19–25. [Google Scholar]
- Jones, S.M. Hormonal Regulation of Ovarian Function in Reptiles. In Hormones and Reproduction of Vertebrates; Elsevier: Amsterdam, The Netherlands, 2011; pp. 89–115. [Google Scholar]
- Ho, S.M.; Kleis, S.; McPherson, R.; Heisermann, G.J.; Callard, I.P. Regulation of vitellogenesis in reptiles. Herpetologica 1982, 38, 40–50. [Google Scholar]
- Guillette, L.J., Jr.; Gross, T.S.; Masson, G.R.; Matter, J.M.; Percival, H.F.; Woodward, A.R. Developmental abnormalities of the gonad and abnormal sex hormone concentrations in juvenile alligators from contaminated and control lakes in Florida. Environ. Health Perspect. 1994, 102, 680–688. [Google Scholar] [CrossRef] [PubMed]
- Guillette, L.J.; Iguchi, T. Contaminant-induced endocrine and reproductive alterations in reptiles. Pure Appl. Chem. 2003, 75, 2275–2286. [Google Scholar] [CrossRef]
- Lind, P.M.; Milnes, M.R.; Lundberg, R.; Bermudez, D.; Orberg, J.A.; Guillette Jr, L.J. Abnormal bone composition in female juvenile American alligators from a pesticide-polluted lake (Lake Apopka, Florida). Environ. Health Perspect. 2004, 112, 359–362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, Y.S.; Stumpf, W.E.; Sar, M.; Christine, M.; Vargas, M. Estrogen and Androgen Target Cells in the Brain of Fishes, Reptiles and Birds: Phylogeny and Ontogeny. Am. Zool. 1978, 18, 425–433. [Google Scholar] [CrossRef] [Green Version]
- Niranjan, M.K.; Koiri, R.K.; Srivastava, R. Expression of estrogen receptor alpha in response to stress and estrogen antagonist tamoxifen in the shell gland of Gallus gallus domesticus: Involvement of anti-oxidant system and estrogen. Stress 2021, 24, 261–272. [Google Scholar] [CrossRef]
- Fry, D.M. Reproductive effects in birds exposed to pesticides and industrial chemicals. Environ. Health Perspect. 1995, 103, 165–171. [Google Scholar]
- Ottinger, M.A.; Quinn, M.J., Jr.; Lavoie, E.; Abdelnabi, M.A.; Thompson, N.; Hazelton, J.L.; Wu, J.M.; Beavers, J.; Jaber, M. Consequences of endocrine disrupting chemicals on reproductive endocrine function in birds: Establishing reliable end points of exposure. Domest. Anim. Endocrinol. 2005, 29, 411–419. [Google Scholar] [CrossRef] [PubMed]
- Vézina, F.; Salvante, K.G.; Williams, T.D. The metabolic cost of avian egg formation: Possible impact of yolk precursor pro-duction? J. Exp. Biol. 2003, 206, 4443–4451. [Google Scholar] [CrossRef] [Green Version]
- Dutta, S.; Inamdar, S.; Tso, J.; Aga, D.S.; Sims, J.T. Free and Conjugated Estrogen Exports in Surface-Runoff from Poultry Litter–Amended Soil. J. Environ. Qual. 2010, 39, 1688–1698. [Google Scholar] [CrossRef] [Green Version]
- Hakk, H.; Millner, P.; Larsen, G. Decrease in Water-Soluble 17β-Estradiol and Testosterone in Composted Poultry Manure with Time. J. Environ. Qual. 2005, 34, 943–950. [Google Scholar] [CrossRef]
- Bird, M.D.; Karavitis, J.; Kovacs, E.J. Sex differences and estrogen modulation of the cellular immune response after injury. Cell. Immunol. 2008, 252, 57–67. [Google Scholar] [CrossRef] [Green Version]
- Ottinger, M.A.; Lavoie, E.; Thompson, N.; Barton, A.; Whitehouse, K.; Barton, M.; Abdelnabi, M.; Quinn, M., Jr.; Panzica, G.; Viglietti-Panzica, C. Neuroendocrine and behavioral effects of embryonic exposure to endocrine disrupting chemicals in birds. Brain Res. Rev. 2008, 57, 376–385. [Google Scholar] [CrossRef] [PubMed]
- Nichols, D.J.; Daniel, T.C.; Moore, P.A., Jr.; Edwards, D.R.; Pote, D.H. Runoff of Estrogen Hormone 17β-Estradiol from Poultry Litter Applied to Pasture; Wiley Online Library: Hoboken, NJ, USA, 1997. [Google Scholar]
- Soma, K.K.; Alday, N.A.; Hau, M.; Schlinger, B.A. Dehydroepiandrosterone metabolism by 3β-hydroxysteroid dehydrogen-ase/Δ5–Δ4 isomerase in adult zebra finch brain: Sex difference and rapid effect of stress. Endocrinology 2004, 145, 1668–1677. [Google Scholar] [CrossRef] [PubMed]
- Berg, C.; Halldin, K.; Fridolfsson, A.-K.; Brandt, I.; Brunström, B. The avian egg as a test system for endocrine disrupters: Effects of diethylstilbestrol and ethynylestradiol on sex organ develo. Sci. Total Environ. 1999, 233, 57–66. [Google Scholar] [CrossRef]
- Heimovics, S.A.; Trainor, B.; Soma, K. Rapid Effects of Estradiol on Aggression in Birds and Mice: The Fast and the Furious: Fig. Integr. Comp. Biol. 2015, 55, 281–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adkins-Regan, E.; Ottinger, M.A.; Park, J. Maternal transfer of estradiol to egg yolks alters sexual differentiation of avian offspring. J. Exp. Zool. 1995, 271, 466–470. [Google Scholar] [CrossRef]
- Yilmaz, B.; Terekeci, H.; Sandal, S.; Kelestimur, F. Endocrine disrupting chemicals: Exposure, effects on human health, mecha-nism of action, models for testing and strategies for prevention. Rev. Endocr. Metab. Disord. 2020, 21, 127–147. [Google Scholar] [CrossRef]
- Adeel, M.; Zain, M.; Fahad, S.; Rizwan, M.; Ameen, A.; Yi, H.; Baluch, M.A.; Lee, J.Y.; Rui, Y. Natural and synthetic estrogens in leafy vegetable and their risk associated to human health. Environ. Sci. Pollut. Res. 2018, 25, 36712–36723. [Google Scholar] [CrossRef]
- Chen, X.; Li, Y.; Jiang, L.; Hu, B.; Wang, L.; An, S.; Zhang, X. Uptake, accumulation, and translocation mechanisms of steroid estrogens in plants. Sci. Total Environ. 2021, 753, 141979. [Google Scholar] [CrossRef]
- Zhou, X.; Yang, Z.; Luo, Z.; Li, H.; Chen, G. Endocrine disrupting chemicals in wild freshwater fishes: Species, tissues, sizes and human health risks. Environ. Pollut. 2019, 244, 462–468. [Google Scholar] [CrossRef]
- Snoj, T.; Majdič, G. Mechanisms in endocrinology: Estrogens in consumer milk: Is there a risk to human reproductive health? Eur. J. Endocrinol. 2018, 179, R275–R286. [Google Scholar] [CrossRef] [PubMed]
- Ribeiro, E.; Ladeira, C.; Viegas, S. EDCs Mixtures: A Stealthy Hazard for Human Health? Toxics 2017, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Kasonga, T.K.; Coetzee, M.A.A.; Kamika, I.; Ngole-Jeme, V.M.; Momba, M.N.B. Endocrine-disruptive chemicals as contami-nants of emerging concern in wastewater and surface water: A review. J. Environ. Manag. 2021, 277, 111485. [Google Scholar] [CrossRef] [PubMed]
- Barakat, R.; Oakley, O.; Kim, H.; Jin, J.; Ko, C.J. Extra-gonadal sites of estrogen biosynthesis and function. BMB Rep. 2016, 49, 488–496. [Google Scholar] [CrossRef] [PubMed]
- Słowikowski, B.K.; Lianeri, M.; Jagodziński, P.P. Exploring estrogenic activity in lung cancer. Mol. Biol. Rep. 2017, 44, 35–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, K.; Cao, X.; He, Y.; Xu, Y. Progress in the molecular understanding of central regulation of body weight by estrogens. Obesity 2015, 23, 919–926. [Google Scholar] [CrossRef] [Green Version]
- De Morentin, P.B.M.; García, I.G.; Martins, L.; Lage, R.; Fernandez, R.L.; Martinez-Sanchez, N.; Ruiz-Pino, F.; Liu, J.; Morgan, D.A.; Pinilla, L.; et al. Estradiol Regulates Brown Adipose Tissue Thermogenesis via Hypothalamic AMPK. Cell Metab. 2014, 20, 41–53. [Google Scholar] [CrossRef] [Green Version]
- Torre, D.; Lolli, F.; Ciana, P.; Maggi, A.; Della Torre, S. Sexual Dimorphism and Estrogen Action in Mouse Liver. Single Mol. Single Cell Seq. 2017, 1043, 141–151. [Google Scholar] [CrossRef]
- Nerozzi, C.; Recuero, S.; Galeati, G.; Bucci, D.; Spinaci, M.; Yeste, M. Effects of Roundup and its main component, glyphosate, upon mammalian sperm function and survival. Sci. Rep. 2020, 10, 1–9. [Google Scholar] [CrossRef]
- Mortimer, S.T. A critical review of the physiological importance and analysis of sperm movement in mammals. Hum. Reprod. Updat. 1997, 3, 403–439. [Google Scholar] [CrossRef] [Green Version]
- Jørgensen, N.; Joensen, U.N.; Jensen, T.K.; Jensen, M.B.; Almstrup, K.; Olesen, I.A.; Juul, A.; Andersson, A.-M.; Carlsen, E.; Petersen, J.H.; et al. Human semen quality in the new millennium: A prospective cross-sectional population-based study of 4867 men. BMJ Open 2012, 2, e000990. [Google Scholar] [CrossRef] [Green Version]
- Stewart, M.K.; Mattiske, D.M.; Pask, A.J. Exogenous Oestrogen Impacts Cell Fate Decision in the Developing Gonads: A Po-tential Cause of Declining Human Reproductive Health. Int. J. Mol. Sci. 2020, 21, 8377. [Google Scholar] [CrossRef] [PubMed]
- Stewart, M.K.; Mattiske, D.M.; Pask, A.J. Estrogen suppresses SOX9 and activates markers of female development in a human testis-derived cell line. BMC Mol. Cell Biol. 2020, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Dostalova, P.; Zatecka, E.; Dvorakova-Hortova, K. Of Oestrogens and Sperm: A Review of the Roles of Oestrogens and Oestrogen Receptors in Male Reproduction. Int. J. Mol. Sci. 2017, 18, 904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wee, S.Y.; Aris, A.Z. Endocrine disrupting compounds in drinking water supply system and human health risk implication. Environ. Int. 2017, 106, 207–233. [Google Scholar] [CrossRef] [PubMed]
- Shemesh, M.; Shore, L.S. Effects of environmental estrogens on reproductive parameters in domestic animals. Isr. J. Vet. Med. 2012, 67, 6–10. [Google Scholar]
- Beck, A.B.; Gardiner, M.R. Clover disease of sheep in Western Australia. J. Dep. Agric. West. Aust. Ser. 4 1965, 6, 388–400. [Google Scholar]
- Setchell, K.; Gosselin, S.; Welsh, M.; Johnston, J.; Balistreri, W.; Kramer, L.; Dresser, B.; Tarr, M. Dietary estrogens—A probable cause of infertility and liver disease in captive cheetahs. Gastroenterology 1987, 93, 225–233. [Google Scholar] [CrossRef]
- Song, R.X.-D.; Santen, R.J. Apoptotic action of estrogen. Apoptosis 2003, 8, 55–60. [Google Scholar] [CrossRef]
- Crescitelli, M.C.; Rauschemberger, M.B.; Cepeda, S.; Sandoval, M.; Massheimer, V.L. Role of estrone on the regulation of osteoblastogenesis. Mol. Cell. Endocrinol. 2019, 498, 110582. [Google Scholar] [CrossRef]
- Chen, J.R.; Plotkin, L.I.; Aguirre, J.I.; Han, L.; Jilka, R.L.; Kousteni, S.; Manolagas, S.C. Transient versus sustained phos-phorylation and nuclear accumulation of ERKs underlie anti-versus pro-apoptotic effects of estrogens. J. Biol. Chem. 2005, 280, 4632–4638. [Google Scholar] [CrossRef] [Green Version]
- Watson, C.S.; Yow-Jiun, J.; Kochukov, M.Y. Nongenomic actions of estradiol compared with estrone and estriol in pituitary tumor cell signaling and proliferation. FASEB J. 2008, 22, 3328–3336. [Google Scholar] [CrossRef] [Green Version]
- Gitsch, E.; Janisch, H.; Spona, J. HPL and Estriol Serum Levels and Placental Perfusion as Indexes of Placental Function in EPH Gestosis and Placental Insufficiency. Gynecol. Obstet. Investig. 1980, 11, 102–112. [Google Scholar] [CrossRef]
- Mawet, M.; Maillard, C.; Klipping, C.; Zimmerman, Y.; Foidart, J.-M.; Bennink, H.J.T.C. Unique effects on hepatic function, lipid metabolism, bone and growth endocrine parameters of estetrol in combined oral contraceptives. Eur. J. Contracept. Reprod. Health Care 2015, 20, 463–475. [Google Scholar] [CrossRef] [PubMed]
- Holinka, C.F.; Diczfalusy, E.; Bennink, H.J.C. Estetrol: A unique steroid in human pregnancy. J. Steroid Biochem. Mol. Biol. 2008, 110, 138–143. [Google Scholar] [CrossRef]
- Grandi, G.; Del Savio, M.C.; da Silva-Filho, A.L.; Facchinetti, F. Estetrol (E4): The new estrogenic component of combined oral contraceptives. Expert Rev. Clin. Pharmacol. 2020, 13, 327–330. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delclos, K.B.; Weis, C.C.; Bucci, T.J.; Olson, G.; Mellick, P.; Sadovova, N.; Latendresse, J.R.; Thorn, B.; Newbold, R.R. Overlapping but distinct effects of genistein and ethinyl estradiol (EE2) in female Sprague-Dawley rats in multigenerational reproductive and chronic toxicity studies. Reprod. Toxicol. 2009, 27, 117–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Li, Y.; Chen, Q.; Liu, Z. Long-term exposure of xenoestrogens with environmental relevant concentrations disrupted spermatogenesis of zebrafish through altering sex hormone balance, stimulating germ cell proliferation, meiosis and enhancing apoptosis. Environ. Pollut. 2019, 244, 486–494. [Google Scholar] [CrossRef]
- Nadzialek, S.; Pigneur, L.-M.; Wéron, B.; Kestemont, P. Bcl-2 and Caspase 3 mRNA levels in the testes of gudgeon, Gobio gobio, exposed to ethinylestradiol (EE2). Aquat. Toxicol. 2010, 98, 304–310. [Google Scholar] [CrossRef] [PubMed]



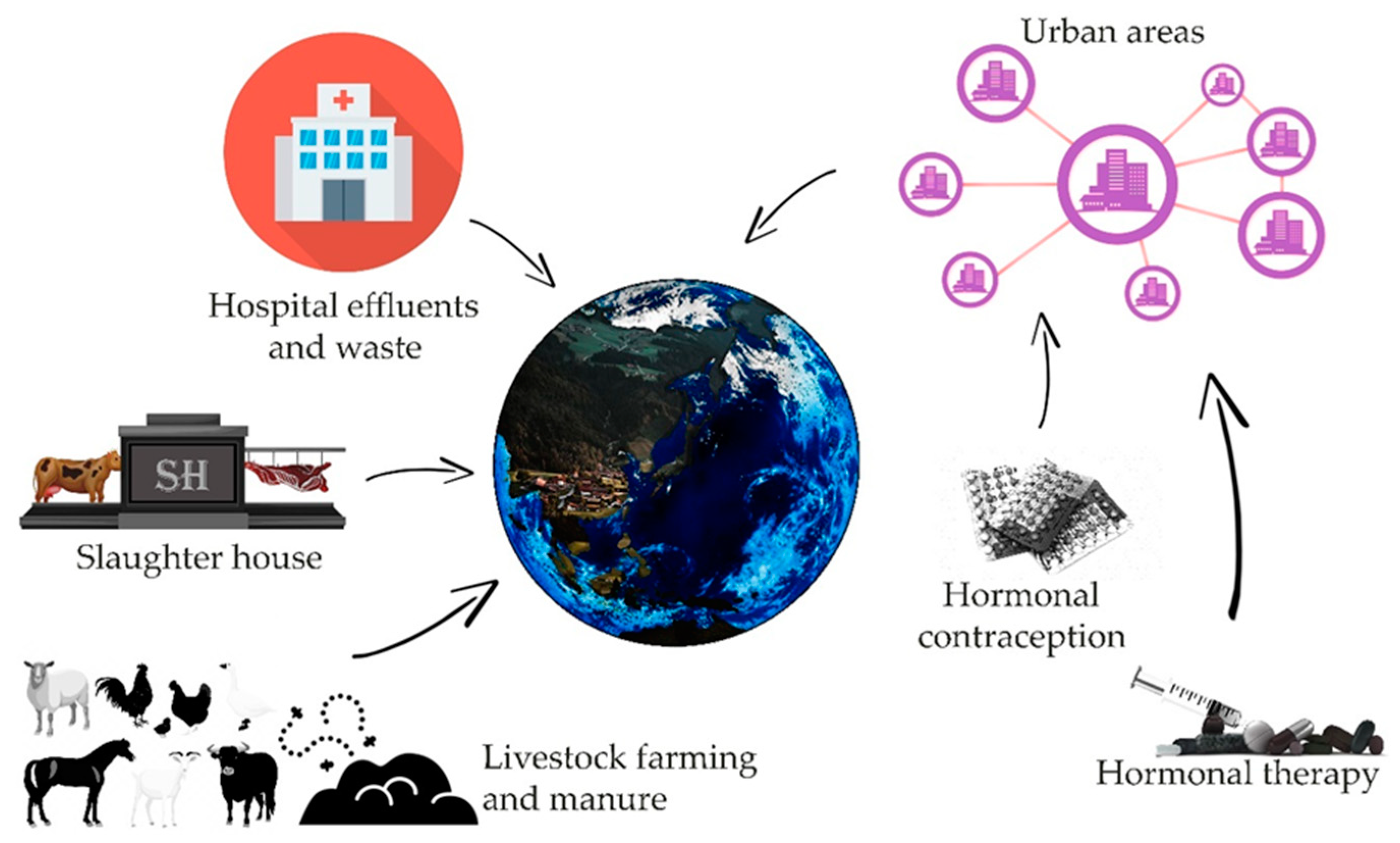{kind=link}
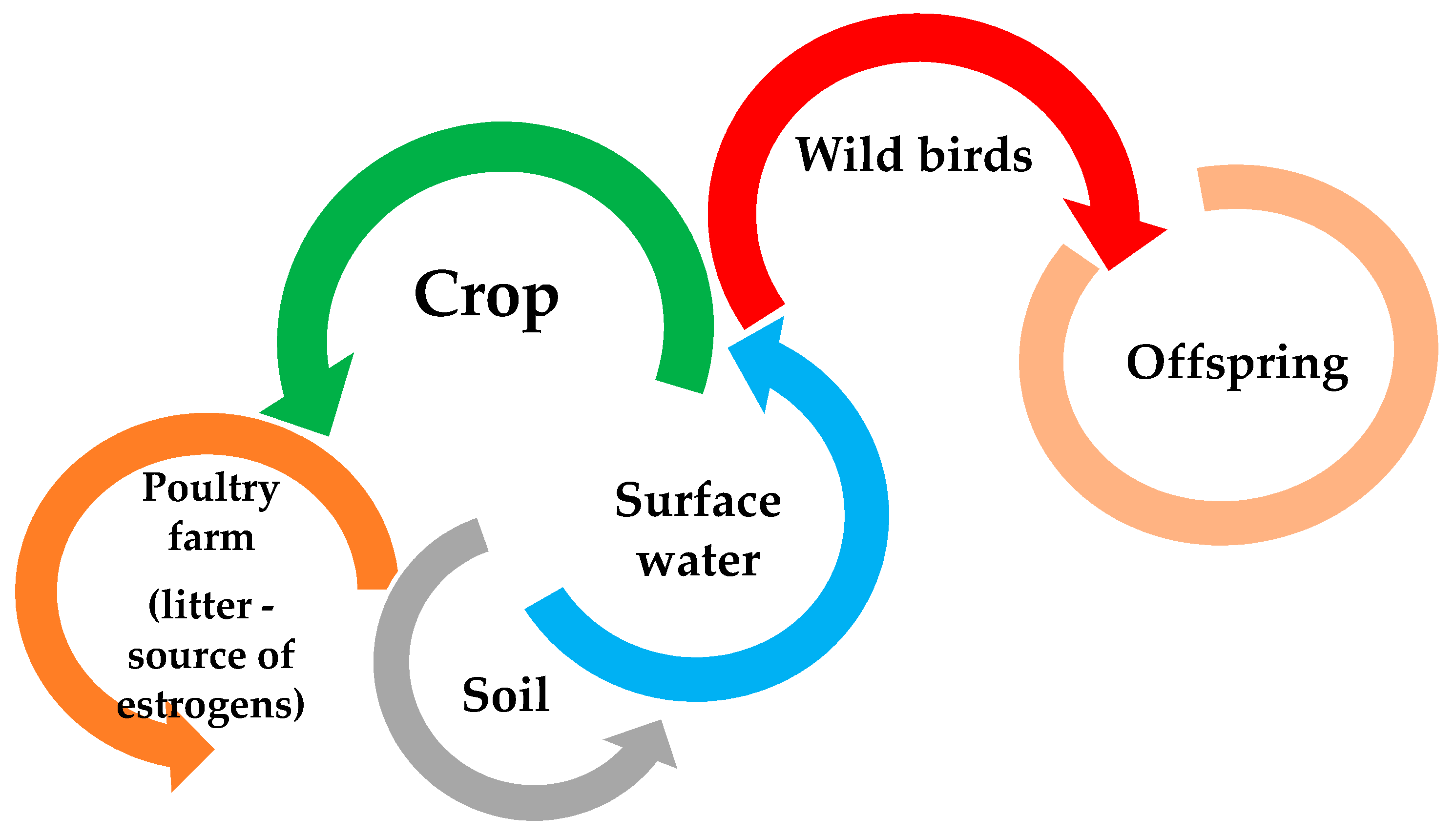{kind=link}
| Estrogen | Skeletal Formula | Role | Effects of Toxicity | References |
|---|---|---|---|---|
| Estrone (E1) |  | Controls the estrous or menstrual cycle in females. Has a vital role in a endocrine system of postmenopausal woman. Plays role in osteoblastogenesis. | Gonadal abnormalities, reduced fertility, males mating behavior impaired, elevated levels of thyrotropin and vitellogenin in blood plasma, elevated caspase-3 level and lowered cholinesterase activity, can induce proapoptotic and anti-apoptotic activity, neurotoxicity. | [5,62,85,129,133,134] |
| Estradiol (E2) |  | Controls the estrous or menstrual cycle in females. Regulates proper development of female reproductive system. Plays role in regulation of neuroendocrine, skeletal and immune systems in males and females. Most potent estrogen. | Gonadal abnormalities, reduced fertility, elevated level of vitellogenin in serum and tissues, can induce proapoptotic and anti-apoptotic activity, neurotoxicity. Impairs animals mating behavior, potentially cancerogenic | [25,77,94,133,135] |
| Estriol (E3) |  | Physiological role is not fully known Regulates uteroplacental blood flow and placental vascularization | Gonadal abnormalities, reduced fertility, elevated level of vitellogenin in serum and tissues | [61,62,85,129,136,137] |
| Estetrol (E4) |  | Physiological role unknown Inhibits ovulation Further research needed | Effects of toxicity are unknown May cause similar effects to other estrogens | [138,139,140] |
| Ethynyloestradiol (EE2) |  | Synthetic estrogen, mainly used in contraceptive pills and hormonal therapy Inhibits ovulation | Reduces fertility, gonadal abnormalities, decreased body weight, accelerated vaginal opening, alteration of estrous cycle, impairment of behavior, formation of lesions, elevated level of vitellogenin in serum and tissues, can induce proapoptotic and anti-apoptotic activity | [5,17,70,141,142,143] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wojnarowski, K.; Podobiński, P.; Cholewińska, P.; Smoliński, J.; Dorobisz, K. Impact of Estrogens Present in Environment on Health and Welfare of Animals. Animals 2021, 11, 2152. https://doi.org/10.3390/ani11072152
Wojnarowski K, Podobiński P, Cholewińska P, Smoliński J, Dorobisz K. Impact of Estrogens Present in Environment on Health and Welfare of Animals. Animals. 2021; 11(7):2152. https://doi.org/10.3390/ani11072152
Chicago/Turabian StyleWojnarowski, Konrad, Paweł Podobiński, Paulina Cholewińska, Jakub Smoliński, and Karolina Dorobisz. 2021. "Impact of Estrogens Present in Environment on Health and Welfare of Animals" Animals 11, no. 7: 2152. https://doi.org/10.3390/ani11072152
APA StyleWojnarowski, K., Podobiński, P., Cholewińska, P., Smoliński, J., & Dorobisz, K. (2021). Impact of Estrogens Present in Environment on Health and Welfare of Animals. Animals, 11(7), 2152. https://doi.org/10.3390/ani11072152