
Relative Late Gestational Muscle and Adipose Thickness Reflect the Amount of Mobilization of These Tissues in Periparturient Dairy Cattle

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Management and Experimental Design
2.2. Feed Intake
2.3. Body Weights and Body Condition Scoring
2.4. Milk Sampling and Analysis
2.5. Blood Sampling and Metabolite Analysis
2.6. Statistical Analysis
3. Results
3.1. Daily Feed Intake
3.2. Tissue Mobilization as Assessed by Ultrasound Evaluation
3.3. Body Weight and Body Condition
3.4. Blood Metabolites and Insulin
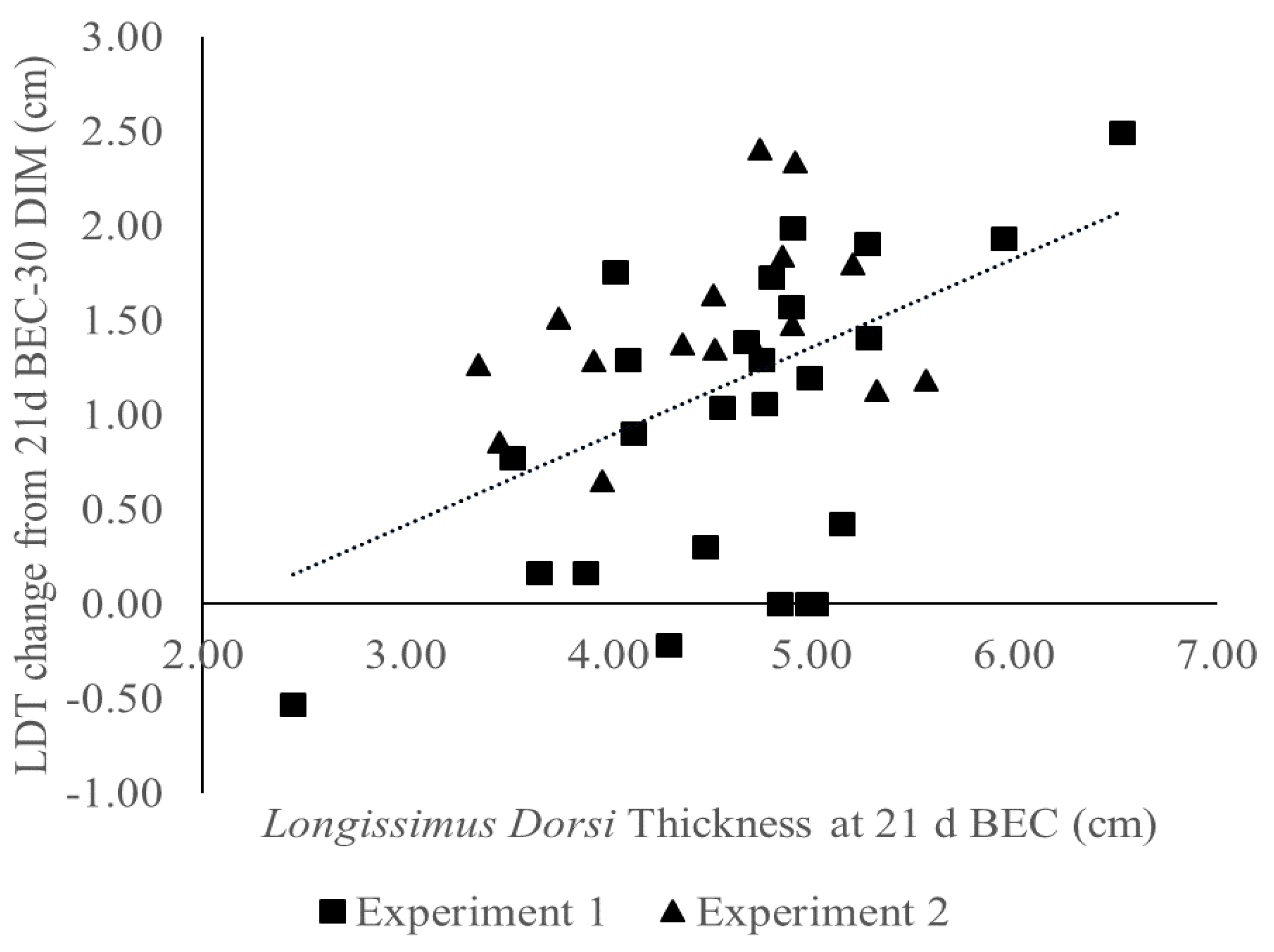
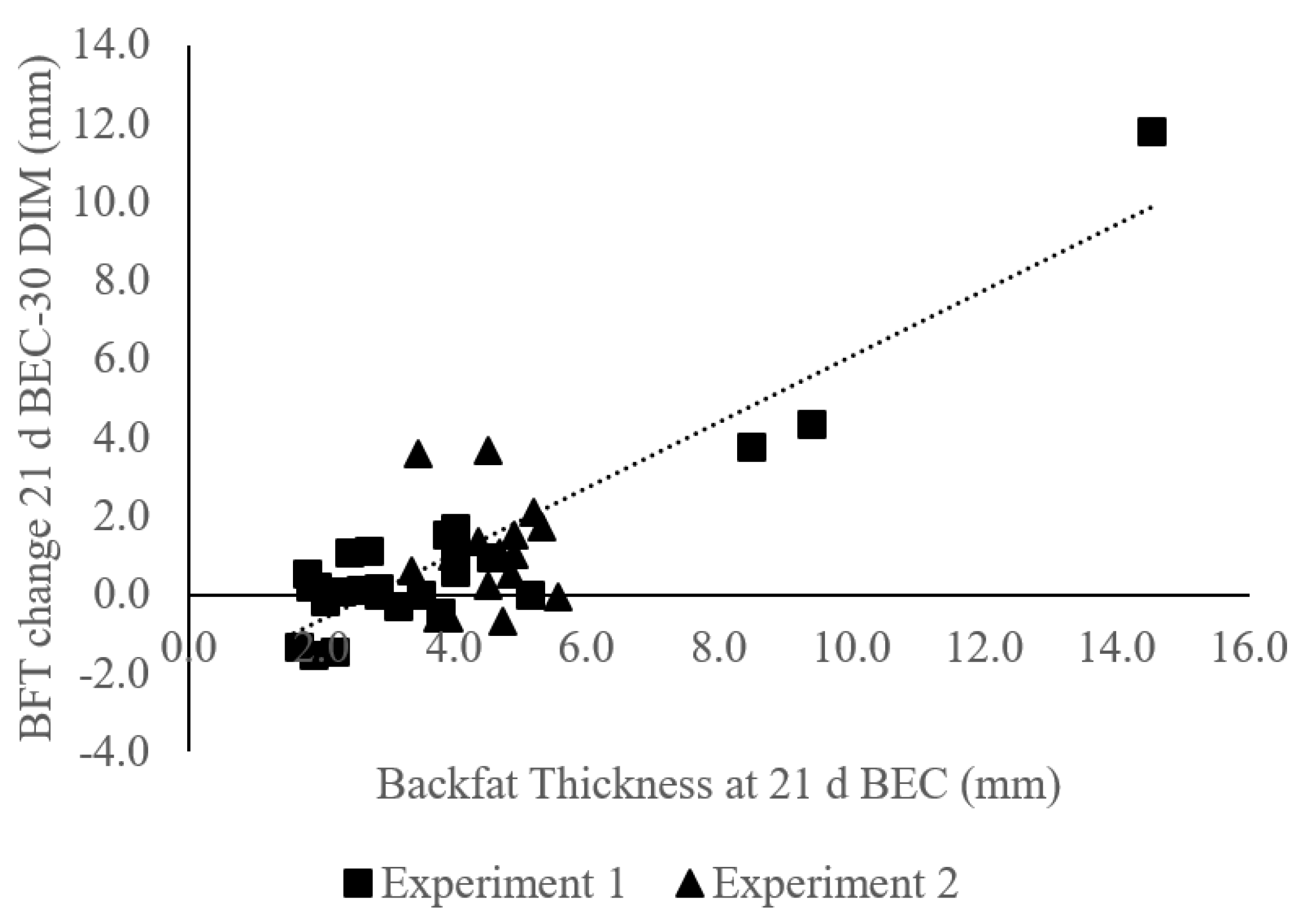
3.5. Cows Mobilized Tissue According to Tissue Thickness at Enrollment
3.6. Production Outcomes
4. Discussion
4.1. Longissimus Dorsi Thickness (LDT) Impact on Muscle Mobilization
4.2. LDT Impact on Adipose Mobilization
4.3. Enrollment Tissue Thickness Negatively Impacted Production
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Acknowledgments
Conflicts of Interest
References
- Bauman, D.E.; Bruce Currie, W. Partitioning of Nutrients During Pregnancy and Lactation: A Review of Mechanisms Involving Homeostasis and Homeorhesis. J. Dairy Sci. 1980, 63, 1514–1529. [Google Scholar] [CrossRef]
- Komaragiri, M.V.S.; Erdman, R.A. Factors Affecting Body Tissue Mobilization in Early Lactation Dairy Cows. 1. Effect of Dietary Protein on Mobilization of Body Fat and Protein. J. Dairy Sci. 1997, 80, 929–937. [Google Scholar] [CrossRef]
- Aschenbach, J.R.; Kristensen, N.B.; Donkin, S.S.; Hammon, H.M.; Penner, G.B. Gluconeogenesis in dairy cows: The secret of making sweet milk from sour dough. IUBMB Life 2010, 62, 869–877. [Google Scholar] [CrossRef]
- De Koster, J.; Hostens, M.; Hermans, K.; Van den Broeck, W.; Opsomer, G. Validation of different measures of insulin sensitivity of glucose metabolism in dairy cows using the hyperinsulinemic euglycemic clamp test as the gold standard. Domest. Anim. Endocrinol. 2016, 57, 117–126. [Google Scholar] [CrossRef]
- Grummer, R.R. Impact of changes in organic nutrient metabolism on feeding the transition dairy cow. J. Anim. Sci. 1995, 73, 2820–2833. [Google Scholar] [CrossRef]
- Van der Drift, S.G.A.; Houweling, M.; Schonewille, J.T.; Tielens, A.G.M.; Jorritsma, R. Protein and fat mobilization and associations with serum β-hydroxybutyrate concentrations in dairy cows. J. Dairy Sci. 2012, 95, 4911–4920. [Google Scholar] [CrossRef]
- Drackley, J.K. ADSA Foundation Scholar Award Biology of Dairy Cows During the Transition Period: The Final Frontier? J. Dairy Sci. 1999, 82, 2259–2273. [Google Scholar] [CrossRef]
- Bell, A.W. Regulation of Organic Nutrient Metabolism During Transition From Late Pregnancy To Early Lactation Efficiency At Pasture View Project. Artic. J. Anim. Sci. 1995. [Google Scholar] [CrossRef]
- De Koster, J.D.; Opsomer, G. Insulin Resistance in Dairy Cows. Vet. Clin. N. Am. Food Anim. Pract. 2013, 29, 299–322. [Google Scholar] [CrossRef]
- Weber, C.; Hametner, C.; Tuchscherer, A.; Losand, B.; Kanitz, E.; Otten, W.; Singh, S.P.; Bruckmaier, R.M.; Becker, F.; Kanitz, W.; et al. Variation In Fat Mobilization During Early Lactation Differently Affects Feed Intake, Body Condition, And Lipid And Glucose Metabolism In High-Yielding Dairy Cows. J. Dairy Sci. 2013, 96, 165–180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBlanc, S.J. Monitoring metabolic health of dairy cattle in the transition period. J. Reprod. Dev. 2010, 56, S29–S35. [Google Scholar] [CrossRef] [Green Version]
- Grummer, R.R. Nutritional and management strategies for the prevention of fatty liver in dairy cattle. Vet. J. 2008, 176, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Pires, J.A.A.; Delavaud, C.; Faulconnier, Y.; Pomiès, D.; Chilliard, Y. Effects of body condition score at calving on indicators of fat and protein mobilization of periparturient Holstein-Friesian cows. J. Dairy Sci. 2013, 96, 6423–6439. [Google Scholar] [CrossRef]
- Kokkonen, T.; Taponen, J.; Anttila, T.; Syrjälä-Qvist, L.; Delavaud, C.; Chilliard, Y.; Tuori, M.; Tesfa, A.T. Effect of body fatness and glucogenic supplement on lipid and protein mobilization and plasma leptin in dairy cows. J. Dairy Sci. 2005, 88, 1127–1141. [Google Scholar] [CrossRef] [Green Version]
- Suarez-Trujillo, A.; Wernert, G.; Sun, H.; Steckler, T.S.; Huff, K.; Cummings, S.; Franco, J.; Klopp, R.N.; Townsend, J.R.; Grott, M.; et al. Exposure to chronic light–dark phase shifts during the prepartum nonlactating period attenuates circadian rhythms, decreases blood glucose, and increases milk yield in the subsequent lactation. J. Dairy Sci. 2020, 103, 2784–2799. [Google Scholar] [CrossRef] [PubMed]
- McCabe, C.J.; Suarez-Trujillo, A.; Teeple, K.A.; Casey, T.M.; Boerman, J.P. Chronic prepartum light-dark phase shifts in cattle disrupt circadian clocks, decrease insulin sensitivity and mammary development, and are associated with lower milk yield through 60 days postpartum. J. Dairy Sci. 2021, 104, 2422–2437. [Google Scholar] [CrossRef] [PubMed]
- Greiner, S.P.; Rouse, G.H.; Wilson, D.E.; Cundiff, L.V.; Wheeler, T.L. The relationship between ultrasound measurements and carcass fat thickness and longissimus muscle area in beef cattle. J. Anim. Sci. 2003, 81, 676–682. [Google Scholar] [CrossRef]
- Bruckmaier, R.M.; Lehmann, E.; Hugi, D.; Hammon, H.M.; Blum, J.W. Ultrasonic measurement of longissimus dorsi muscle and backfat, associated with metabolic and endocrine traits, during fattening of intact and castrated male cattle. Livest. Prod. Sci. 1998, 53, 123–134. [Google Scholar] [CrossRef]
- Schwager-Suter, R.; Stricker, C.; Erdin, D.; Künzi, N. Relationship between body condition scores and ultrasound measurements of subcutaneous fat and m. longissimus dorsi in dairy cows differing in size and type. Anim. Sci. 2020, 71, 465–470. [Google Scholar] [CrossRef]
- Schäff, C.; Börner, S.; Hacke, S.; Kautzsch, U.; Sauerwein, H.; Spachmann, S.K.; Schweigel-Röntgen, M.; Hammon, H.M.; Kuhla, B. Increased muscle fatty acid oxidation in dairy cows with intensive body fat mobilization during early lactation. J. Dairy Sci. 2013, 96, 6449–6460. [Google Scholar] [CrossRef]
- Megahed, A.A.; Hiew, M.W.H.; Ragland, D.; Constable, P.D. Changes in skeletal muscle thickness and echogenicity and plasma creatinine concentration as indicators of protein and intramuscular fat mobilization in periparturient dairy cows. J. Dairy Sci. 2019, 102, 5550–5565. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirments of Dairy Cattle, 7th ed.; National Academies Press: Washington, DC, USA, 2001. [Google Scholar]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods For Dietary Fiber, Neutral Detergent Fiber, And Nonstarch Polysaccharides In Relation To Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef]
- Wildman, E.E.; Jones, I.G.M.; Wagner, P.E.; Boman, R.L.; Troutt, H.F.; Lesch, T.N. A Dairy Cow Body Condition Scoring System and Its Relationship to Selected Production Characteristics. J. Dairy Sci. 1982, 65, 495–501. [Google Scholar] [CrossRef]
- Iwersen, M.; Falkenberg, U.; Voigtsberger, R.; Foderung, D.; Heuwieser, W. Evaluation of an electronic cowside test to detect moderate ketosis in dairy cows. J. Dairy Sci. 2009, 2618–2624. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Wang, Y.; Yuan, B.; Liu, S.; Man, S.; Xu, H.; Lu, X. A novel LC-MS/MS Assay For the Simultaneous Determination of Melatonin and Its Two Major Metabolites, 6-Hydroxymelatonin and 6-Sulfatoxymelatonin in Dog Plasma: Application to A Pharmacokinetic Study. J. Pharm. Biomed. Anal. 2016, 117, 390–397. [Google Scholar] [CrossRef]
- Doepel, L.; Lapierre, H.; Kennelly, J.J. Peripartum Performance and Metabolism of Dairy Cows in Response to Prepartum Energy and Protein Intake. J. Dairy Sci. 2002, 85, 2315–2334. [Google Scholar] [CrossRef] [Green Version]
- Wyss, M.; Kaddurah-Daouk, R. Creatine and Creatinine Metabolism. Physiol. Rev. 2000, 80, 1107–1213. [Google Scholar] [CrossRef]
- Houweling, M.; van der Drift, S.G.A.; Jorritsma, R.; Tielens, A.G.M. Technical Note: Quantification of Plasma 1- And 3-Methylhistidine in Dairy Cows by High-Performance Liquid Chromatography-Tandem Mass Spectrometry. J. Dairy Sci. 2012, 95, 3125–3130. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reynolds, C.K.; Dürst, B.; Lupoli, B.; Humphries, D.J.; Beever, D.E. Visceral Tissue Mass and Rumen Volume in Dairy Cows during the Transition from Late Gestation to Early Lactation. J. Dairy Sci. 2004, 87, 961–971. [Google Scholar] [CrossRef] [Green Version]
- Bünemann, K.; Von Soosten, D.; Frahm, J.; Kersten, S.; Meyer, U.; Hummel, J.; Zeyner, A.; Dänicke, S. Effects of Body Condition and Concentrate Proportion of the Ration on Mobilization of Fat Depots and Energetic Condition in Dairy Cows During Early Lactation Based on Ultrasonic Measurements. Animals 2019, 9, 131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strieder-Barboza, C.; Zondlak, A.; Kayitsinga, J.; Pires, A.F.A.; Contreras, G.A. Lipid Mobilization Assessment in Transition Dairy Cattle Using Ultrasound Image Biomarkers. Livest. Sci. 2015, 177, 159–164. [Google Scholar] [CrossRef]
- Mann, S.; Nydam, D.V.; Abuelo, A.; Leal Yepes, F.A.; Overton, T.R.; Wakshlag, J.J. Insulin Signaling, Inflammation, And Lipolysis in Subcutaneous Adipose Tissue of Transition Dairy Cows either Overfed Energy during the Prepartum Period or Fed A Controlled-Energy Diet. J. Dairy Sci. 2016, 99, 6737–6752. [Google Scholar] [CrossRef]
- Akter, S.H.; Häussler, S.; Dänicke, S.; Müller, U.; von Soosten, D.; Rehage, J.; Sauerwein, H. Physiological and Conjugated Linoleic Acid-Induced Changes of Adipocyte Size in Different Fat Depots of Dairy Cows During Early Lactation. J. Dairy Sci. 2011, 94, 2871–2882. [Google Scholar] [CrossRef] [Green Version]
- Roche, J.R.; Bell, A.W.; Overton, T.R.; Loor, J.J. Nutritional Management of The Transition Cow in the 21st Century-A Paradigm Shift in Thinking. Anim. Prod. Sci. 2013, 53, 1000–1023. [Google Scholar] [CrossRef]
- Tamminga, S.; Luteijn, P.A.; Meijer, R.G.M. Changes in Composition and Energy Content of Liveweight Loss of Dairy Cows with Time After Parturition. Livest. Prod. Sci. 1997, 52, 31–38. [Google Scholar] [CrossRef]
- Larsen, M.; Galindo, C.; Ouellet, D.R.; Maxin, G.; Kristensen, N.B.; Lapierre, H. Abomasal Amino Acid Infusion in Postpartum Dairy Cows: Effect on Whole-Body, Splanchnic, and Mammary Amino Acid Metabolism. J. Dairy Sci. 2015, 98, 7944–7961. [Google Scholar] [CrossRef] [Green Version]
- Larsen, M.; Lapierre, H.; Kristensen, N.B. Abomasal Protein Infusion In Postpartum Transition Dairy Cows: Effect On Performance And Mammary Metabolism. J. Dairy Sci. 2014, 97, 5608–5622. [Google Scholar] [CrossRef]
- Bell, A.W.; Slepetis, R.; Ehrhardt, R.A. Growth and Accretion of Energy and Protein in the Gravid Uterus During Late Pregnancy in Holstein Cows. J. Dairy Sci. 1995, 75, 1954–1961. [Google Scholar] [CrossRef]
- Stalker, L.A.; Ciminski, L.A.; Adams, D.C.; Klopfenstein, T.J.; Clark, R.T. Effects Of Weaning Date And Prepartum Protein Supplementation On Cow Performance And Calf Growth. Rangel. Ecol. Manag. 2007, 60, 578–587. [Google Scholar] [CrossRef]
- Batistel, F.; Alharthi, A.S.; Yambao, R.R.C.; Elolimy, A.A.; Pan, Y.X.; Parys, C.; Loor, J.J. Methionine Supply during Late-Gestation Triggers Offspring Sex-Specific Divergent Changes in Metabolic and Epigenetic Signatures in Bovine Placenta. J. Nutr. 2019, 149, 6–17. [Google Scholar] [CrossRef]
- Zachut, M.; Honig, H.; Striem, S.; Zick, Y.; Boura-Halfon, S.; Moallem, U. Periparturient Dairy Cows Do Not Exhibit Hepatic Insulin Resistance, Yet Adipose-Specific Insulin Resistance Occurs In Cows Prone To High Weight Loss. J. Dairy Sci. 2013, 96, 5656–5669. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Komaragiri, M.V.S.; Casper, D.P.; Erdman, R.A. Factors affecting body tissue mobilization in early lactation dairy cows. 2. Effect of dietary fat on mobilization of body fat and protein. J. Dairy Sci. 1998, 81, 169–175. [Google Scholar] [CrossRef]
- Jaurena, G.; Moorby, J.M. Lactation and body composition responses to fat and protein supplies during the dry period in under-conditioned dairy cows. J. Dairy Sci. 2017, 100, 1107–1121. [Google Scholar] [CrossRef] [Green Version]
- Douglas, G.N.; Overton, T.R.; Bateman, H.G.; Dann, H.M.; Drackley, J.K. Prepartal plane of nutrition, regardless of dietary energy source, affects periparturient metabolism and dry matter intake in Holstein cows. J. Dairy Sci. 2006, 89, 2141–2157. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients (g/100 g Diet DM) | First Experiment | Second Experiment | ||
|---|---|---|---|---|
| Prefresh | Lactating | Prefresh | Lactating | |
| Corn silage | 32.7 | 24.6 | 31.9 | 29.8 |
| Mixed mostly legume silage | 5.4 | 19.1 | 5.3 | 8.3 |
| Rye hay | 16.2 | 7.8 | - | - |
| Wheat straw | - | - | 15.2 | 6.9 |
| Alfalfa hay | - | - | - | 9.3 |
| Whole fuzzy cottonseed | - | 6.0 | - | - |
| Dry ground corn | 14.4 | 4.8 | 14.9 | 9.6 |
| High moisture corn | - | 17.4 | - | 14.8 |
| Cottonseed hulls | 6.6 | 7.8 | - | |
| Soybean meal | 6.4 | 5.5 | 6.7 | 5.8 |
| Soyplus 1 | 3.6 | 4.8 | 4.3 | 4.5 |
| Distillers grain with solubles | 0.7 | - | - | - |
| LysAAMet 2 | 0.6 | 0.8 | 0.7 | 0.8 |
| Calcium carbonate | 1.2 | 1.1 | 1.2 | 1.0 |
| Biochlor 3 | 7.0 | - | 6.9 | - |
| Vitamin and mineral mix | 2.9 4 | 3.1 5 | 3.0 4 | 2.7 5 |
| QLF 63/43 6 | 2.3 | 3.7 | 2.2 | 4.1 |
| Palmit 80 7 | - | 1.3 | - | 2.2 |
| Nutrient Composition (% diet DM unless otherwise stated) | ||||
| Dry matter (% of diet) 8 | 51.6 | 53.2 | 52.9 | 51.8 |
| Crude protein 9 | 16.9 | 14.2 | 15.9 | 13.8 |
| NDF 10 | 35.5 | 22.3 | 37.5 | 27.2 |
| ADF 10 | 20.5 | 15.8 | 24.4 | 18.4 |
| Ash 11 | 7.8 | 7.9 | 5.7 | 5.1 |
| Starch 12 | 24.6 | 26.8 | 24.6 | 28.7 |
| NFC 12 | 37.2% | 42.0% | 37.2% | 43.3% |
| Metabolizable energy (Mcal/kg) 12 | 2.40 | 2.62 | 2.37 | 2.58 |
| 35 BEC | 0 DIM | 30 DIM | 60 DIM | p-Values | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| HM | LM | HM | LM | HM | LM | HM | LM | SEM 1 | Group 2 | Day | Group × Day | |
| BCS | 3.63 a | 3.22 c | 3.49 b | 3.19 c | 2.97 d | 2.74 e | 2.78 e | 2.72 e | 0.07 | <0.01 | <0.01 | 0.02 |
| BCS Δ per mo. | - | - | −0.05 a | −0.07 a | −0.47 b | −0.45 b | −0.15 a | −0.01 a | 0.13 | 0.13 | <0.01 | 0.06 |
| BW (kg) | 780 a | 722 b | 745 b | 699 c | 661 d | 628 d | 655 d | 632 d | 14.3 | 0.02 | <0.01 | 0.21 |
| BW Δ (kg) | - | - | −24.2 | −28.2 | −74.4 | −72.5 | 0.0 | −2.5 | 8.5 | 0.98 | <0.01 | 0.90 |
| LDT 3 Δ per mo. (cm) | - | - | −0.30 b,c | 0.51 a | −1.05 d | −0.61 c | −0.18 b | −0.20 b | 0.15 | <0.01 | <0.01 | <0.01 |
| Prepartum/Postpartum LDT Δ (cm) | - | - | −0.39 b | 0.43 a | - | - | −1.15 c | −0.95 c | 0.17 | <0.01 | <0.01 | 0.11 |
| Overall LDT Δ (cm) | - | - | - | - | - | - | −1.54 | −0.52 | 0.21 | <0.01 | - | - |
| BFT 4 Δ per mo. (mm) | - | - | 0.65 a | 0.03 a,b | −1.18 c | −0.46 b,c | −0.39 b,c | −0.09 a,b | 0.30 | 0.50 | <0.01 | 0.07 |
| Prepartum/Postpartum BFT Δ (mm) | - | - | 0.38 a | 0.00 a,b | - | - | −1.73 c | −0.68 b | 0.34 | 0.46 | <0.01 | 0.03 |
| Overall BFT Δ (mm) | - | - | - | - | - | - | −0.90 | −0.73 | 0.15 | 0.73 | - | - |
| Item | Group 1 | p-Values | ||||
|---|---|---|---|---|---|---|
| HM | LM | SEM | Group | Day | Group × Day | |
| Insulin (ng/mL) 2 | 0.63 | 0.56 | 0.04 | 0.56 | 0.24 | 0.98 |
| Glucose (mg/dL) 3 | 76.8 | 79.8 | 0.9 | 0.03 | 0.12 | 0.31 |
| BHB (mmol/L) 3 | 0.49 | 0.56 | 0.04 | 0.26 | 0.07 | 0.31 |
| NEFA (mmol/L) 4 | 0.18 | 0.18 | 0.03 | 0.96 | <0.01 | 0.54 |
| CRE (ng/mL) 2 | 3328 | 3086 | 94 | 0.06 | 0.10 | 0.29 |
| 3-MH (ng/mL) 2 | 453 | 395 | 29 | 0.09 | 0.14 | 0.04 |
| 3-MH:CRE2 | 0.136 | 0.121 | 0.007 | 0.30 | 0.15 | 0.11 |
| 2 DIM | 7 DIM | 14 DIM | 21 DIM | p-Value | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Item | HM | LM | HM | LM | HM | LM | HM | LM | SEM 1 | Group 2 | Day | Group × Day |
| Insulin (ng/mL) 3 | 0.21 | 0.37 | 0.18 | 0.41 | 0.15 | 0.31 | 0.31 | 0.24 | 0.10 | 0.26 | 0.76 | 0.27 |
| Glucose (mg/dL) 3 | 68.1 | 69.8 | 70.8 | 64.6 | 66.3 | 64.8 | 69.6 | 68.8 | 2.2 | 0.32 | 0.17 | 0.13 |
| BHB (mmol/L) 3 | 0.97 | 1.04 | 1.15 | 1.11 | 1.08 | 0.99 | 0.95 | 0.87 | 0.17 | 0.80 | 0.48 | 0.94 |
| NEFA (mmol/L) 3 | 0.64 | 0.54 | 0.72 | 0.62 | 0.63 | 0.63 | 0.75 | 0.51 | 0.09 | 0.15 | 0.72 | 0.38 |
| CRE (ng/mL) 3 | 3588 | 3477 | 3398 | 3407 | 3173 | 3178 | 3196 | 2994 | 125 | 0.57 | <0.01 | 0.50 |
| 3-MH (ng/mL) 3 | 493 a,b,c | 437 c,d | 538 a | 414 d | 581 a | 471 c | 520 a,b | 428 c,d | 34 | 0.02 | <0.01 | 0.17 |
| 3-MH:CRE 3 | 0.137 c | 0.122 c | 0.163 b | 0.118 c | 0.188 a | 0.143 b,c | 0.167 b | 0.141 b,c | 0.010 | <0.01 | <0.01 | 0.07 |
| Item | HM | LM | SEM | Group p-Values 1 |
|---|---|---|---|---|
| Calf Birthweight (kg) | 45.2 | 41.8 | 0.7 | <0.01 |
| Milk Yield (kg/d) 2 | 38.8 | 41.6 | 0.8 | 0.02 |
| Fat% 3 | 4.33 | 4.05 | 0.12 | 0.09 |
| Protein% 3 | 2.83 | 2.88 | 0.03 | 0.28 |
| Lactose%3 | 4.75 | 4.71 | 0.04 | 0.42 |
| Fat Yield (g) 3 | 1703 | 1612 | 66 | 0.32 |
| Protein Yield (g) 3 | 1102 | 1160 | 34 | 0.21 |
| Lactose Yield (g) 3 | 1843 | 1918 | 57 | 0.35 |
| Milk Urea Nitrogen (mg/dL) 3 | 7.44 | 8.08 | 0.30 | 0.12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
McCabe, C.; Suarez-Trujillo, A.; Casey, T.; Boerman, J. Relative Late Gestational Muscle and Adipose Thickness Reflect the Amount of Mobilization of These Tissues in Periparturient Dairy Cattle. Animals 2021, 11, 2157. https://doi.org/10.3390/ani11082157
McCabe C, Suarez-Trujillo A, Casey T, Boerman J. Relative Late Gestational Muscle and Adipose Thickness Reflect the Amount of Mobilization of These Tissues in Periparturient Dairy Cattle. Animals. 2021; 11(8):2157. https://doi.org/10.3390/ani11082157
Chicago/Turabian StyleMcCabe, Conor, Aridany Suarez-Trujillo, Theresa Casey, and Jacquelyn Boerman. 2021. "Relative Late Gestational Muscle and Adipose Thickness Reflect the Amount of Mobilization of These Tissues in Periparturient Dairy Cattle" Animals 11, no. 8: 2157. https://doi.org/10.3390/ani11082157
APA StyleMcCabe, C., Suarez-Trujillo, A., Casey, T., & Boerman, J. (2021). Relative Late Gestational Muscle and Adipose Thickness Reflect the Amount of Mobilization of These Tissues in Periparturient Dairy Cattle. Animals, 11(8), 2157. https://doi.org/10.3390/ani11082157