
Pathophysiology of Fever and Application of Infrared Thermography (IRT) in the Detection of Sick Domestic Animals: Recent Advances

 ,
,  ,
,  ,
, 
 , , ,
, , , 
Abstract
:Simple Summary
Abstract
Methodology
1. Introduction
2. Pathophysiology of Fever and Adverse Effects
2.1. Pathophysiology of Fever
2.2. Adverse Effects of Fever
2.3. Difference between Fever and Hyperthermia
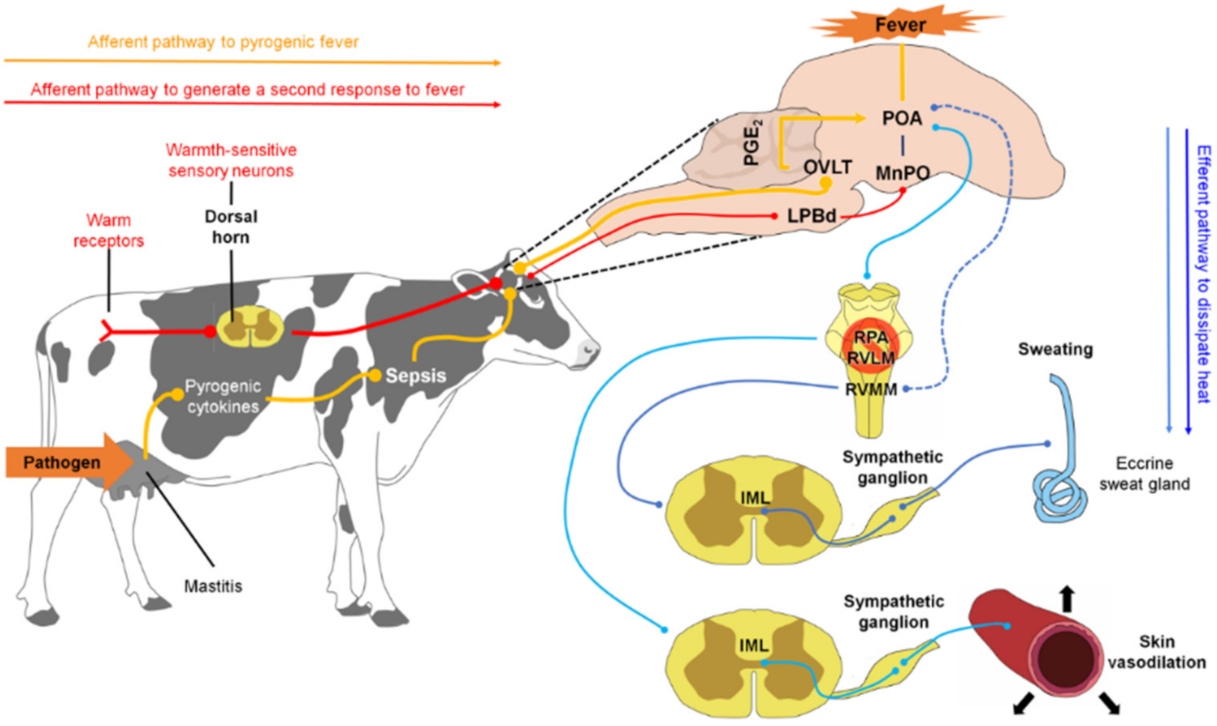
3. How Does an Animal with a Fever Thermoregulate? (Neurophysiological Responses to Temperature Control)
3.1. Central Modulation of Fever
Return to Homeostasis
3.2. Metabolic Mechanisms of Thermoregulation in the Febrile Response at the Central and Peripheral Levels and Cellular Events
3.2.1. Thermogenesis in Brown Adipose Tissue (BAT)
3.2.2. Thermogenesis in Musculoskeletal Tissue (Shivering)
3.2.3. Responses in Cutaneous Microcirculation
3.2.4. Sudomotor Response
3.3. Behavioral Mechanism of Thermoregulation during Fever
3.3.1. Thermoregulatory Behavior in Mammals
3.3.2. Thermoregulatory Behavior in Birds
4. Importance of IRT in the Detection of Sick Farm Animals
4.1. Coccidiosis
4.1.1. Rabbits
4.1.2. Birds
4.2. Mastitis
4.2.1. Bovines
4.2.2. Sheep
4.3. Foot Problems
4.3.1. Bovines
4.3.2. Horses
4.3.3. Birds
4.4. Bovine Respiratory Complex Diseases

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Objective | Findings | Implications | Authors |
|---|---|---|---|---|
| Rabbits | To determine the changes in the thermal profile of rabbits infected with Eimeria intestinalis through thermographic images to detect the variations caused by the infection. | The inoculated group with the highest number of oocysts showed a statistically significant decrease in rectal, ocular surface, and auricular pavilion temperatures, in contrast to the remaining groups (inoculated with a lower amount and uninoculated group control). | Since coccidiosis significantly affects the thermal state of animals, it is suggested that IRT can be considered a reliable method to diagnose this disease. | Vadlejch et al. [138] |
| Horses | To assess the correlation of hoof surface temperature and digital blood vessel ultrasound measurements to determine whether these measurements can be used as predictors of clinical lameness. | There is a 0.99 correlation between the hoof surface temperature and the velocity of blood flow in the distal limb, indicating that the temperature could be used to predict the velocity with which the blood flows from the foot. | Infrared thermography could be used to detect vasodilation and vasoconstriction responses in horses’ feet, which could be associated with lameness and its degree. | Douthit et al. [165] |
| Bovines | To evaluate the effectiveness of thermography in the early detection of clinical mastitis induced by E. coli lipopolysaccharides. | An increase of 1–1.5 °C was detected in the quarters that presented induced mastitis 2 h before the diagnostic tests related to milk. | Thermography can be a valuable tool for rapidly detecting changes in udder surface temperature associated with clinical mastitis, even during the dry period. | Hovinen et al. [155]; Metzner et al. [161] |
| To evaluate the subclinical mastitis detection capacity of infrared thermography compared to California Mastitis Test (CMT). | A 2.35 °C increase was detected in quarters with subclinical mastitis than healthy quarters; there is a positive correlation between thermographic data and the CMT score (0.86). | Thermography is sensitive enough to detect superficial thermal changes in the udder caused by subclinical mastitis, a pathology challenging to diagnose. | Polat et al. [153] | |
| To evaluate the use of IRT as a method of detection of cattle infected with foot-and-mouth disease (FMD). | With the IRT, it was possible to detect the infection of 58.3% of the steers inoculated directly and 50% of the animals that had contact with the first ones, who presented an increase in the temperature of the feet between 4.7 and 8.9 °C, before developing foot injuries. | IRT can be used to detect FMDV-infected animals early, in the preclinical phase (before presenting foot lesions) and during the pre-viremic phase, or those that must be sampled, to speed up the implementation of quarantines. | Rainwater-Lovett et al. [159] | |
| To determine the efficiency of IRT as a method for the early diagnosis of respiratory diseases in bovines. | Values obtained by the IRT method were more efficient than the clinical values of hematological measurements and physical examinations. The authors even describe that the method has predictive potential when detecting true positives. | Considering that early diagnosis of these diseases is necessary to reduce bad practices with antibiotics, IRT is an efficient, non-invasive method that reflects results in real-time compared to other methods. | Schaefer et al. [186] | |
| Sheep | To assess whether IRT can be used as a diagnostic tool for mastitis in sheep. | With the IRT, it was possible to correctly classify the mastitis status of the animals in 73%, where it was detected that the subclinical mastitis group had a higher temperature (36.3 °C) than in the clinical mastitis groups (35.89 °C) and healthy sheep (36.06 °C). | Unlike when milk components are analyzed, IRT can be used as an auxiliary method to diagnose subclinical and clinical mastitis in sheep. | Martins et al. [143] |
| Birds | To assess the detection of footpad dermatitis (FPD) through variations in the surface temperature of the footpad and the whole plantar foot surface in turkeys. | It was possible to detect a mild FPD in most turkeys through IRT, which presented lower surface plantar foot temperatures, the larger their area with discoloration, was associated with hyperkeratosis or ischemic necrosis. | IRT is a reliable tool that can be used to detect subclinical pathologies in the turkey foot. | Moe et al. [174] |
| Assess the variations in the thermal profile of broilers during Eimeria tenella infection and provide ranges or criteria for future evaluations with the IRT method. | At 15 days after inoculation, a statistically significant decrease in temperature was observed in areas such as the shank and the bill surface in the group administered the highest amount of the purified strain. | IRT can be used as an effective method to detect diseases such as coccidiosis, and the areas described as most sensitive to temperature changes in broilers are the shanks and the bill. | Knizkova et al. [139] |
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kluger, M.J. Fever: Role of pyrogens and cryogens. Physiol. Rev. 1991, 71, 93–127. [Google Scholar] [CrossRef]
- Roth, J.; Horowitz, M. Inflammation, fever, and body temperature under febrile conditions. J. Basic Clin. Physiol. Pharmacol. 2017, 28, 519–520. [Google Scholar] [CrossRef] [PubMed]
- Roth, J.; Blatteis, C.M. Mechanisms of fever production and lysis: Lessons from experimental LPS fever. Compr. Physiol. 2014, 4, 1563–1604. [Google Scholar] [CrossRef]
- Bartfai, T.; Conti, B. Fever. Sci. World J. 2010, 10, 490–503. [Google Scholar] [CrossRef]
- Sevegnani, K.B.; Fernandes, D.P.B.; Modenese-Gorla Da Silva, S.H. Evaluation of thermorregulatory capacity of dairy buffaloes using infrared thermography. Eng. Agric. 2016, 36, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Mota-Rojas, D.; Titto, C.G.; Orihuela, A.; Martínez-Burnes, J.; Gómez-Prado, J.; Torres-Bernal, F.; Flores-Padilla, K.; Carvajal-de la Fuente, V.; Wang, D. Physiological and behavioral mechanisms of thermoregulation in mammals. Animals 2021, 11, 1733. [Google Scholar] [CrossRef]
- Broom, D.M. Behaviour and welfare in relation to pathology. Appl. Anim. Behav. Sci. 2006, 97, 73–83. [Google Scholar] [CrossRef]
- Narayan, E.; Perakis, A.; Meikle, W. Using thermal imaging to monitor body temperature of koalas (Phascolarctos cinereus) in a zoo setting. Animals 2019, 9, 1094. [Google Scholar] [CrossRef] [Green Version]
- Kluger, M.J.; Kozak, W.; Conn, C.A.; Leon, L.R.; Soszynski, D. The adaptive value of fever. Infect. Dis. Clin. N. Am. 1996, 10, 1–20. [Google Scholar] [CrossRef]
- Zeisberger, E. From humoral fever to neuroimmunological control of fever. J. Therm. Biol. 1999, 24, 287–326. [Google Scholar] [CrossRef]
- Fidel, R.-R.; Farias, J.M. La fiebre. Rev. Fac. Med. 2014, 57, 20–33. [Google Scholar]
- Prajitha, N.; Athira, S.S.; Mohanan, P.V. Pyrogens, a polypeptide produces fever by metabolic changes in hypothalamus: Mechanisms and detections. Immunol. Lett. 2018, 204, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Steiner, A.A.; Ivanov, A.I.; Serrats, J.; Hosokawa, H.; Phayre, A.N.; Robbins, J.R.; Roberts, J.L.; Kobayashi, S.; Matsumura, K.; Sawchenko, P.E.; et al. Cellular and molecular bases of the initiation of fever. PLoS Biol. 2006, 4, 1517–1524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janssens, S.; Beyaert, R. Role of toll-like receptors in pathogen recognition. Clin. Microbiol. Rev. 2003, 16, 637–646. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mackowiak, P.A. Concepts of fever. Arch. Intern. Med. 1998, 158, 1870–1881. [Google Scholar] [CrossRef] [Green Version]
- Walter, E.J.; Hanna-Jumma, S.; Carraretto, M.; Forni, L. The pathophysiological basis and consequences of fever. Crit. Care 2016, 20, 1–10. [Google Scholar] [CrossRef] [Green Version]
- Dantzer, R.; Konsman, J.-P.; Bluthé, R.-M.; Kelley, K.W. Neural and humoral pathways of communication from the immune system to the brain: Parallel or convergent? Auton. Neurosci. 2000, 85, 60–65. [Google Scholar] [CrossRef]
- Kita, Y.; Yoshida, K.; Tokuoka, S.M.; Hamano, F.; Yamazaki, M.; Sakimura, K.; Kano, M.; Shimizu, T. Fever is mediated by conversion of endocannabinoid 2-arachidonoylglycerol to prostaglandin E2. PLoS ONE 2015, 10, e0133663. [Google Scholar] [CrossRef] [Green Version]
- Li, Z.; Perlik, V.; Feleder, C.; Tang, Y.; Blatteis, C.M. Kupffer cell-generated PGE 2 triggers the febrile response of guinea pigs to intravenously injected LPS. Am. J. Physiol. Integr. Comp. Physiol. 2006, 290, R1262–R1270. [Google Scholar] [CrossRef] [Green Version]
- Ricciotti, E.; Fitzgerald, G.A. Prostaglandins and inflammation. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 986–1000. [Google Scholar] [CrossRef]
- Lazarus, M. The differential role of prostaglandin E2 receptors EP3 and EP4 in regulation of fever. Mol. Nutr. Food Res. 2006, 50, 451–455. [Google Scholar] [CrossRef]
- Simons, C.T.; Kulchitsky, V.A.; Sugimoto, N.; Homer, L.D.; Székely, M.; Romanovsky, A.A. Signaling the brain in systemic inflammation: Which vagal branch is involved in fever genesis? Am. J. Physiol. Regul. Integr. Comp. Physiol. 1998, 275, 63–68. [Google Scholar] [CrossRef]
- Hildebrandt, B.; Wust, P.; Ahler, O.; Dieing, A.; Sreenivasa, G.; Kerner, T.; Felix, R.; Riess, H. The cellular and molecular basis of hyperthermia. Crit. Rev. Oncol. Hematol. 2002, 43, 33–56. [Google Scholar] [CrossRef]
- Lambert, H.; Carder, G. Positive and negative emotions in dairy cows: Can ear postures be used as a measure? Behav. Process. 2019, 158, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Vlad, M.; Ionescu, N.; Ispas, A.T.; Giuvărăşteanu, I.; Ungureanu, E.; Stoica, C. Cambios morfológicos durante la hipertermia aguda experimental a corto plazo. Rom. J. Morphol. Embryol 2010, 51, 739–744. [Google Scholar] [PubMed]
- Badoer, E. Role of the hypothalamic PVN in the regulation of renal sympathetic nerve activity and blood flow during hyperthermia and in heart failure. Am. J. Physiol. Physiol. 2010, 298, F839–F846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Diehl, K.A.; Crawford, E.; Shinko, P.D.; Tallman, R.D.; Oglesbee, M.J. Alterations in hemostasis associated with hyperthermia in a canine model. Am. J. Hematol. 2000, 64, 262–270. [Google Scholar] [CrossRef]
- White, M.G.; Luca, L.E.; Nonner, D.; Saleh, O.; Hu, B.; Barrett, E.F.; Barrett, J.N. Cellular mechanisms of neuronal damage from hyperthermia. Prog. Brain Res. 2007, 162, 347–371. [Google Scholar] [PubMed]
- Zaremba, J. Hyperthermia in ischemic stroke. Med. Sci. Monit. 2004, 10, 148–153. [Google Scholar]
- White, M.G.; Emery, M.; Nonner, D.; Barrett, J.N. Caspase activation contributes to delayed death of heat-stressed striatal neurons. J. Neurochem. 2003, 87, 958–968. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hinoue, A.; Fushiki, S.; Nishimura, Y.; Shiota, K. In utero exposure to brief hyperthermia interferes with the production and migration of neocortical neurons and induces apoptotic neuronal death in the fetal mouse brain. Dev. Brain Res. 2001, 132, 59–67. [Google Scholar] [CrossRef]
- Henrique, F.L.; Zanella, A.J.; Bezerra, H.V.A.; Polato, H.Z.; Fernandes, A.C.; Hooper, H.B.; Pulido-Rodríguez, L.F.; Titto, E.A.L.; Pereira, A.M.F.; Titto, C.G. Stress during first gestation of ewes impairs memory and learning of male offspring. Vet. Res. Commun. 2021. [Google Scholar] [CrossRef]
- Lifshitz, A. Fiebre y otras formas de elevación térmica. Rev. Investig. Clín. 2007, 59, 130–138. [Google Scholar]
- Gómez, A.-E. Trastornos de la temperatura corporal: Etiología y medidas preventivas. Offarm Farm. Soc. 2007, 26, 48–53. [Google Scholar]
- Olmos, S.S. Fiebre. Farm. Prof. 2017, 31, 18–23. [Google Scholar]
- Madden, C.J.; Morrison, S.F. Central nervous system circuits that control body temperature. Neurosci. Lett. 2019, 696, 225–232. [Google Scholar] [CrossRef] [PubMed]
- Mota-Rojas, D.; Napolitano, F.; Strappini, A.; Orihuela, A.; Ghezzi, M.D.; Hernández-ávalos, I.; Mora-Medina, P.; Whittaker, A.L. Pain at the slaughterhouse in ruminants with a focus on the neurobiology of sensitisation. Animals 2021, 11, 1085. [Google Scholar] [CrossRef]
- Oka, T. Psychogenic fever: How psychological stress affects body temperature in the clinical population. Temperature 2015, 2, 368–378. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Aguiar Bittencourt, M.; Melleu, F.F.; Marino-Neto, J. Stress-induced core temperature changes in pigeons (Columba livia). Physiol. Behav. 2015, 139, 449–458. [Google Scholar] [CrossRef] [Green Version]
- Pedernera-Romano, C.; Ruiz de la Torre, J.L.; Badiella, L.; Manteca, X. Effect of perphenazine enanthate on open-field test behaviour and stress-induced hyperthermia in domestic sheep. Pharmacol. Biochem. Behav. 2010, 94, 329–332. [Google Scholar] [CrossRef]
- Raison, C.L.; Capuron, L.; Miller, A.H. Cytokines sing the blues: Inflammation and the pathogenesis of depression. Trends Immunol. 2006, 27, 24–31. [Google Scholar] [CrossRef] [Green Version]
- Evans, S.S.; Repasky, E.A.; Fisher, D.T. Fever and the thermal regulation of immunity: The immune system feels the heat. Nat. Rev. Immunol. 2015, 15, 335–349. [Google Scholar] [CrossRef]
- Schieber, A.M.P.; Ayres, J.S. Thermoregulation as a disease tolerance defense strategy. Pathog. Dis. 2016, 74, 106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saper, C.B.; Romanovsky, A.A.; Scammell, T.E. Neural circuitry engaged by prostaglandins during the sickness syndrome. Nat. Neurosci. 2012, 15, 1088–1095. [Google Scholar] [CrossRef]
- Oka, T. Stress-induced hyperthermia and hypothermia. In Handbook of Clinical Neurology; Elsevier, B.V.: Amsterdam, The Netherlands, 2018; Volume 157, pp. 599–621. [Google Scholar]
- Hasday, J.D.; Thompson, C.; Singh, I.S. Fever, immunity, and molecular adaptations. In Comprehensive Physiology; Wiley: Hoboken, NJ, USA, 2014; pp. 109–148. [Google Scholar]
- Evans, S.S.; Wang, W.-C.; Bain, M.D.; Burd, R.; Ostberg, J.R.; Repasky, E.A. Fever-range hyperthermia dynamically regulates lymphocyte delivery to high endothelial venules. Blood 2001, 97, 2727–2733. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.-T.; Zhong, L.; Mace, T.A.; Repasky, E.A. Elevation in body temperature to fever range enhances and prolongs subsequent responsiveness of macrophages to endotoxin challenge. PLoS ONE 2012, 7, e30077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tan, C.L.; Cooke, E.K.; Leib, D.E.; Lin, Y.C.; Daly, G.E.; Zimmerman, C.A.; Knight, Z.A. Warm-sensitive neurons that control body temperature. Cell 2016, 167, 47–59.e15. [Google Scholar] [CrossRef] [Green Version]
- Contreras, C.; Nogueiras, R.; Diéguez, C.; Rahmouni, K.; López, M. Traveling from the hypothalamus to the adipose tissue: The thermogenic pathway. Redox Biol. 2017, 12, 854–863. [Google Scholar] [CrossRef]
- Nakamura, K.; Morrison, S.F. Central efferent pathways for cold-defensive and febrile shivering. J. Physiol. 2011, 589, 3641–3658. [Google Scholar] [CrossRef]
- Ootsuka, Y.; Tanaka, M. Control of cutaneous blood flow by central nervous system. Temperature 2015, 2, 392–405. [Google Scholar] [CrossRef]
- Mota-Rojas, D.; Olmos-Hernández, A.; Verduzco-Mendoza, A.; Lecona-Butrón, H.; Martínez-Burnes, J.; Mora-Medina, P.; Gómez-Prado, J.; Orihuela, A. Infrared thermal imaging associated with pain in laboratory animals. Exp. Anim. 2021, 70, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Green, H.D.; Howard, W.B.; Kenan, L.F. Autonomic control of blood flow in hind paw of the dog. Am. J. Physiol. Content 1956, 187, 469–472. [Google Scholar] [CrossRef] [PubMed]
- Koss, M.C. Characterization of adrenoceptor subtypes in cat cutaneous vasculature. J. Pharmacol. Exp. Ther. 1990, 254, 221–227. [Google Scholar] [PubMed]
- Ruffolo, R.R. Distribution and function of peripheral α-adrenoceptors in the cardiovascular system. Pharmacol. Biochem. Behav. 1985, 22, 827–833. [Google Scholar] [CrossRef]
- Grayson, J. Cold and warmth vasoconstrictor responses in the skin of man. Heart 1951, 13, 167–176. [Google Scholar] [CrossRef] [Green Version]
- Plazas, R.A.S.; Díaz Ávila, V. Mecanismos fisiológicos de la termorregulación en animales de producción. Rev. Colomb. Cienc. Anim. 2011, 4, 88–94. [Google Scholar]
- Tanaka, M.; Nagashima, K.; McAllen, R.M.; Kanosue, K. Role of the medullary raphé in the thermoregulatory vasomotor control in rats. J. Physiol. 2002, 540, 657–664. [Google Scholar] [CrossRef] [PubMed]
- Morrison, S.F.; Nakamura, K. Central mechanisms for thermoregulation. Annu. Rev. Physiol. 2019, 81, 285–308. [Google Scholar] [CrossRef]
- Tanaka, M.; McKinley, M.J.; McAllen, R.M. Roles of two preoptic cell groups in tonic and febrile control of rat tail sympathetic fibers. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2009, 296, 1248–1257. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, K.; Matsumura, K.; Kaneko, T.; Kobayashi, S.; Katoh, H.; Negishi, M. The rostral raphe pallidus nucleus mediates pyrogenic transmission from the preoptic area. J. Neurosci. 2002, 22, 4600–4610. [Google Scholar] [CrossRef] [Green Version]
- Rathner, J.A.; Madden, C.J.; Morrison, S.F. Central pathway for spontaneous and prostaglandin E2-evoked cutaneous vasoconstriction. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2008, 295, 343–354. [Google Scholar] [CrossRef]
- Rice, P.; Martin, E.; He, J.-R.; Frank, M.; DeTolla, L.; Hester, L.; O’Neill, T.; Manka, C.; Benjamin, I.; Nagarsekar, A.; et al. Febrile-range hyperthermia augments neutrophil accumulation and enhances lung injury in experimental gram-negative bacterial pneumonia. J. Immunol. 2005, 174, 3676–3685. [Google Scholar] [CrossRef] [Green Version]
- Hales, J.R.S. Thermoregulatory requirements for circulatory adjustments to promote heat loss in animals. J. Therm. Biol. 1983, 8, 219–224. [Google Scholar] [CrossRef]
- Zhao, Z.D.; Yang, W.Z.; Gao, C.; Fu, X.; Zhang, W.; Zhou, Q.; Chen, W.; Ni, X.; Lin, J.K.; Yang, J.; et al. A hypothalamic circuit that controls body temperature. Proc. Natl. Acad. Sci. USA 2017, 114, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greenberg, R.; Cadena, V.; Danner, R.M.; Tattersall, G. Heat loss may explain bill size differences between birds occupying different habitats. PLoS ONE 2012, 7, e40933. [Google Scholar] [CrossRef]
- Hagan, A.A.; Heath, J.E. Regulation of heat loss in the duck by vasomotion in the bill. J. Therm. Biol. 1980, 5, 95–101. [Google Scholar] [CrossRef]
- Phillips, P.K.; Sanborn, A.F. An infrared, thermographic study of surface temperature in three ratites: Ostrich, emu and double-wattled cassowary. J. Therm. Biol. 1994, 19, 423–430. [Google Scholar] [CrossRef]
- Tattersall, G.J.; Andrade, D.V.; Abe, A.S. Heat exchange from the toucan bill reveals a controllable vascular thermal radiator. Science 2009, 325, 468–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryeland, J.; Weston, M.A.; Symonds, M.R.E. Bill size mediates behavioural thermoregulation in birds. Funct. Ecol. 2017, 31, 885–893. [Google Scholar] [CrossRef] [Green Version]
- Drake, R.; Wayne, A.; Mitchell, A. Gray. Anatomía Básica, 2nd ed.; Elsevier: Amsterdam, The Netherlands, 2018. [Google Scholar]
- Baker, L.B. Physiology of sweat gland function: The roles of sweating and sweat composition in human health. Temperature 2019, 6, 211–259. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K. The physiology, pharmacology, and biochemistry of the eccrine sweat gland. Rev. Physiol. Biochem. Pharmacol. 1977, 79, 51–131. [Google Scholar] [CrossRef]
- Bavera, G.; Beguet, H. Termor Regulación Corporal y Ambientación; Facultad Agronomía y Veterinaria UNRC: Córdoba, Argentina, 2003. [Google Scholar]
- de Mattos Nascimento, M.R.B.; Dias, E.A.; dos Santos, T.R.; Ayres, G.F.; Nascimento, C.C.N.; Beletti, M.E. Effects of age on histological parameters of the sweat glands of Nellore cattle. Rev. Ceres 2015, 62, 129–132. [Google Scholar] [CrossRef] [Green Version]
- Hoffman, T.C.M.; Walsberg, G.E.; DeNardo, D.F. Cloacal evaporation: An important and previously undescribed mechanism for avian thermoregulation. J. Exp. Biol. 2007, 210, 741–749. [Google Scholar] [CrossRef] [Green Version]
- Wenger, C.B. Heat of evaporation of sweat: Thermodynamic considerations. J. Appl. Physiol. 1972, 32, 456–459. [Google Scholar] [CrossRef]
- Joyner, M.J.; Coyle, E.F. Endurance exercise performance: The physiology of champions. J. Physiol. 2008, 586, 35–44. [Google Scholar] [CrossRef]
- Magoun, H.W.; Harrison, F.; Brobeck, J.R.; Ranson, S.W. Activation of heat loss mechanism by local heating of the brain. J. Neurophysiol. 1938, 1, 101–114. [Google Scholar] [CrossRef]
- Davison, M.; Koss, M. Brainstem loci for activation of electrodermal response in the cat. Am. J. Physiol. Content 1975, 229, 930–934. [Google Scholar] [CrossRef]
- Shafton, A.D.; McAllen, R.M. Location of cat brain stem neurons that drive sweating. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2013, 304, 804–809. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sato, K. The mechanism of eccrine sweat secretion. In Exercise, Heat, and Thermoregulation; Gisolfi, D.R.L.C.V., Nadel, E.R., Eds.; Brown & Benchmark: Dubuque, IA, USA, 1993. [Google Scholar]
- Sato, K.; Ohtsuyama, M.; Samman, G. Eccrine sweat gland disorders. J. Am. Acad. Dermatol. 1991, 24, 1010–1014. [Google Scholar] [CrossRef]
- Sato, K. The physiology and pharmacology of the eccrine sweat gland. In Biochemistry and Physiology of the Skin; Goldsmith, L., Ed.; Oxford University Press: New York, NY, USA, 1983; pp. 596–641. [Google Scholar]
- Nejsum, L.N.; Kwon, T.-H.; Jensen, U.B.; Fumagalli, O.; Frokiaer, J.; Krane, C.M.; Menon, A.G.; King, L.S.; Agre, P.C.; Nielsen, S. Functional requirement of aquaporin-5 in plasma membranes of sweat glands. Proc. Natl. Acad. Sci. USA 2002, 99, 511–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inoue, R.; Sohara, E.; Rai, T.; Satoh, T.; Yokozeki, H.; Sasaki, S.; Uchida, S. Immunolocalization and translocation of aquaporin-5 water channel in sweat glands. J. Dermatol. Sci. 2013, 70, 26–33. [Google Scholar] [CrossRef]
- Xie, L.; Jin, L.; Feng, J.; Lv, J. The expression of AQP5 and UTs in the sweat glands of uremic patients. Biomed. Res. Int. 2017, 2017, 8629783. [Google Scholar] [CrossRef]
- Costill, D.L. Sweating: Its composition and effects on body fluids. Ann. N. Y. Acad. Sci. 1977, 301, 160–174. [Google Scholar] [CrossRef]
- Robinson, S.; Robinson, A.H. Chemical composition of sweat. Physiol. Rev. 1954, 34, 202–220. [Google Scholar] [CrossRef] [PubMed]
- Costill, D.L.; Cote, R.; Fink, W. Muscle water and electrolytes following varied levels of dehydration in man. J. Appl. Physiol. 1976, 40, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.C.; Steiner, A.A.; Branco, L.G.S.; Romanovsky, A.A. Cold-seeking behavior as a thermoregulatory strategy in systemic inflammation. Eur. J. Neurosci. 2006, 23, 3359–3367. [Google Scholar] [CrossRef] [PubMed]
- Kanosue, K.; Nakayama, T.; Tanaka, H.; Yanase, M.; Yasuda, H. Modes of action of local hypothalamic and skin thermal stimulation on salivary secretion in rats. J. Physiol. 1990, 424, 459–471. [Google Scholar] [CrossRef] [PubMed]
- Roberts, W.W.; Mooney, R.D. Brain areas controlling thermoregulatory grooming prone extension, locomotion, and tail vasodilation in rats. J. Comp. Physiol. Psychol. 1974, 86, 470–480. [Google Scholar] [CrossRef]
- Lipton, J. Effects of preoptic lesions on heat-escape responding and colonic temperature in the rat. Physiol. Behav. 1968, 3, 165–169. [Google Scholar] [CrossRef]
- Carlisle, H.J. Effect of preoptic and anterior hypothalamic lesions on behavioral thermoregulation in the cold. J. Comp. Physiol. Psychol. 1969, 69, 391–402. [Google Scholar] [CrossRef]
- Satinoff, E.; Rutstein, J. Behavioral thermoregulation in rats with anterior hypothalamic lesions. J. Comp. Physiol. Psychol. 1970, 71, 77–82. [Google Scholar] [CrossRef]
- Roberts, W.W.; Martin, J.R. Effects of lesions in central thermosensitive areas on thermoregulatory responses in rat. Physiol. Behav. 1977, 19, 503–511. [Google Scholar] [CrossRef]
- Lendez, P.A.; Martinez Cuesta, L.; Nieto Farias, M.V.; Vater, A.A.; Ghezzi, M.D.; Mota-Rojas, D.; Dolcini, G.L.; Ceriani, M.C. Alterations in TNF-α and its receptors expression in cows undergoing heat stress. Vet. Immunol. Immunopathol. 2021, 235, 110232. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.; Mascovich, A.; Gillin, J.C.; Willoughby, R.; Pike, J.; Smith, T.L. Partial sleep deprivation reduces natural killer cell activity in humans. Psychosom. Med. 1994, 56, 493–498. [Google Scholar] [CrossRef] [PubMed]
- Gregory, N. Physiological mechanisms causing sickness behaviour and suffering in diseased animals. Anim. Welf. 1998, 7, 293–305. [Google Scholar]
- Dallman, M.F. Stress and sickness decrease food intake and body weight. In Coping with Challenge: Welfare in Animals Including Humans; Dahlem University Press: Berlin, Germany, 2001; pp. 300–316. [Google Scholar]
- Gregory, N. Physiology and Behaviour of Animal Suffering; Gregory, N.G., Ed.; Blackwell Publishing: Oxford, UK, 2004; ISBN 9780470752494. [Google Scholar]
- Briese, E. Selected temperature correlates with intensity of fever in rats. Physiol. Behav. 1997, 61, 659–660. [Google Scholar] [CrossRef]
- Terrien, J.; Perret, M.; Aujard, F. Behavioral thermoregulation in mammals: A review. Front. Biosci. 2011, 16, 1428–1444. [Google Scholar] [CrossRef] [Green Version]
- Hanya, G.; Kiyono, M.; Hayaishi, S. Behavioral thermoregulation of wild Japanese macaques: Comparisons between two subpopulations. Am. J. Primatol. 2007, 69, 802–815. [Google Scholar] [CrossRef]
- Warnecke, L.; Schleucher, E.; Geiser, F. Basking behaviour in relation to energy use and food availability in one of the smallest marsupials. Physiol. Behav. 2010, 101, 389–393. [Google Scholar] [CrossRef]
- Gilbert, C.; McCafferty, D.; Le Maho, Y.; Martrette, J.-M.; Giroud, S.; Blanc, S.; Ancel, A. One for all and all for one: The energetic benefits of huddling in endotherms. Biol. Rev. 2010, 85, 545–569. [Google Scholar] [CrossRef]
- Gaughan, J.B.; Bonner, S.; Loxton, I.; Mader, T.L.; Lisle, A.; Lawrence, R. Effect of shade on body temperature and performance of feedlot steers1. J. Anim. Sci. 2010, 88, 4056–4067. [Google Scholar] [CrossRef] [Green Version]
- Sullivan, M.L.; Cawdell-Smith, A.J.; Mader, T.L.; Gaughan, J.B. Effect of shade area on performance and welfare of short-fed feedlot cattle. J. Anim. Sci. 2011, 89, 2911–2925. [Google Scholar] [CrossRef]
- Kotze, J.; Bennett, N.C.; Scantlebury, M. The energetics of huddling in two species of mole-rat (Rodentia: Bathyergidae). Physiol. Behav. 2008, 93, 215–221. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Šumbera, R.; Zelová, J.; Kunc, P.; Knížková, I.; Burda, H. Patterns of surface temperatures in two mole-rats (Bathyergidae) with different social systems as revealed by IR-thermography. Physiol. Behav. 2007, 92, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Caro, T. Contrasting coloration in terrestrial mammals. Philos. Trans. R. Soc. B Biol. Sci. 2009, 364, 537–548. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, M.B.; Langman, V.A.; Richard Taylor, C. Panting in dogs: Paths of air flow in response to heat and exercise. Respir. Physiol. 1981, 43, 327–338. [Google Scholar] [CrossRef]
- Hales, J.R.; Brown, G. Net energetic and thermoregulatory efficiency during panting in the sheep. Comp. Biochem. Physiol. Part A Physiol. 1974, 49, 413–422. [Google Scholar] [CrossRef]
- Krönert, H.; Pleschka, K. Lingual blood flow and its hypothalamic control in the dog during panting. Pflug. Arch. Eur. J. Physiol. 1976, 367, 25–31. [Google Scholar] [CrossRef] [PubMed]
- Pleschka, K. The part played by lingual blood flow in the canine heat loss mechanism. In Central Control Mechanisms and Related Topics in Honor of S.C. Wang; Wang, H., Blumenthal, M.R., Ngai, S.H., Eds.; Futura: New York, NY, USA, 1980. [Google Scholar]
- Mack, L.A.; Felver-Gant, J.N.; Dennis, R.L.; Cheng, H.W. Genetic variations alter production and behavioral responses following heat stress in 2 strains of laying hens. Poult. Sci. 2013, 92, 285–294. [Google Scholar] [CrossRef]
- Quiles, A.; Hevia, M. Termorregulación en las Gallinas—BM Editores: Mexico City, Mexico. 2018. Available online: https://bmeditores.mx/avicultura/termorregulacion-en-las-gallinas-1590/ (accessed on 21 June 2021).
- Bhadauria, P. Management of Heat Stress in Poultry Production System; ICAR-Agricultural Technology Application Research Institute: Ludhiana, India, 2017. [Google Scholar]
- Fedde, M.R. Relationship of structure and function of the avian respiratory system to disease susceptibility. Poult. Sci. 1998, 77, 1130–1138. [Google Scholar] [CrossRef]
- Du Plessis, K.L.; Martin, R.O.; Hockey, P.A.R.; Cunningham, S.J.; Ridley, A.R. The costs of keeping cool in a warming world: Implications of high temperatures for foraging, thermoregulation and body condition of an arid-zone bird. Glob. Chang. Biol. 2012, 18, 3063–3070. [Google Scholar] [CrossRef]
- Cunningham, S.J.; Martin, R.O.; Hockey, P.A. Can behaviour buffer the impacts of climate change on an arid-zone bird? Ostrich 2015, 86, 119–126. [Google Scholar] [CrossRef]
- Stewart, M.; Stafford, K.J.; Dowling, S.K.; Schaefer, A.L.; Webster, J.R. Eye temperature and heart rate variability of calves disbudded with or without local anaesthetic. Physiol. Behav. 2008, 93, 789–797. [Google Scholar] [CrossRef] [PubMed]
- Soerensen, D.D.; Pedersen, L.J. Infrared skin temperature measurements for monitoring health in pigs: A review. Acta Vet. Scand. 2015, 57, 1–11. [Google Scholar] [CrossRef] [Green Version]
- McManus, C.; Tanure, C.B.; Peripolli, V.; Seixas, L.; Fischer, V.; Gabbi, A.M.; Menegassi, S.R.O.; Stumpf, M.T.; Kolling, G.J.; Dias, E.; et al. Infrared thermography in animal production: An overview. Comput. Electron. Agric. 2016, 123, 10–16. [Google Scholar] [CrossRef]
- Johnson, S.R.; Rao, S.; Hussey, S.B.; Morley, P.S.; Traub-Dargatz, J.L. Thermographic eye temperature as an index to body temperature in ponies. J. Equine Vet. Sci. 2011, 31, 63–66. [Google Scholar] [CrossRef]
- Edgar, J.L.; Nicol, C.J.; Pugh, C.A.; Paul, E.S. Surface temperature changes in response to handling in domestic chickens. Physiol. Behav. 2013, 119, 195–200. [Google Scholar] [CrossRef] [PubMed]
- Cook, N.J.; Chabot, B.; Lui, T.; Bench, C.J.; Schaefer, A.L. Infrared thermography detects febrile and behavioural responses to vaccination of weaned piglets. Animal 2015, 9, 339–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Y.-Y.; Hao, S.; Zhang, M.; Zhang, X.; Wang, D. Aquaporins, evaporative water loss and thermoregulation in heat-acclimated Mongolian gerbils (Meriones unguiculatus). J. Therm. Biol. 2020, 91, 102641. [Google Scholar] [CrossRef]
- Alsaaod, M.; Büscher, W. Detection of hoof lesions using digital infrared thermography in dairy cows. J. Dairy Sci. 2012, 95, 735–742. [Google Scholar] [CrossRef]
- Napolitano, F.; Mota-Rojas, D.; Guerrero Legarreta, I.; Orihuela, A. The Latin American River Buffalo, Recent Findings, 3rd ed.; BM Editores Press: Mexico City, Mexico, 2020; pp. 1–1545. Available online: https://www.lifescienceglobal.com/journals/journal-of-buffalo-science/97-abstract/jbs/4550-el-bufalo-de-agua-en-latinoamerica-hallazgos-recientes (accessed on 18 December 2020).
- Mota-Rojas, D.; Pereira, A.M.F.; Wang, D.; Martínez-Burnes, J.; Ghezzi, M.; Hernández-Avalos, I.; Lendez, P.; Mora-Medina, P.; Casas-Alvarado, A.; Olmos-Hernández, A.; et al. Clinical applications and factors involved in validating thermal windows used in infrared thermography in cattle and river buffalo to assess health and productivity. Animals 2021, 11, 2247. [Google Scholar] [CrossRef]
- Hovinen, M.; Pyörälä, S. Invited review: Udder health of dairy cows in automatic milking. J. Dairy Sci. 2011, 94, 547–562. [Google Scholar] [CrossRef] [PubMed]
- Nahm, F.S. Infrared thermography in pain medicine. Korean J. Pain 2013, 26, 219–222. [Google Scholar] [CrossRef]
- De Gussem, M. Coccidiosis in poultry: Review on diagnosis, control, prevention and interaction with overall gut health. In Proceedings of the 16th European Symposium on Poultry Nutrition, Strasbourg, France, 26–30 August 2007; pp. 253–261. [Google Scholar]
- Witlock, D.R.; Ruff, M.D.; Chute, M.B. Physiological basis of Eimeria tenella-induced mortality in individual chickens. J. Parasitol. 1981, 67, 65–69. [Google Scholar] [CrossRef] [PubMed]
- Vadlejch, J.; Knížková, I.; Makovcová, K.; Kunc, P.; Jankovská, I.; Janda, K.; Borkovcová, M.; Langrová, I. Thermal profile of rabbits infected with Eimeria intestinalis. Vet. Parasitol. 2010, 171, 343–345. [Google Scholar] [CrossRef]
- Knížková, I.; Kunc, P.; Langrová, I.; Vadlejch, J.; Jankovská, I. Thermal profile of broilers infected by Eimeria tenella. In Proceedings of the 2018 International Conference on Quantitative InfraRed Thermography, Berlin, Germany, 25–29 June 2018; pp. 9–12. [Google Scholar]
- South, K.E.; Haynes, K.; Jackson, A.C. Diagnosis of hypothermia in the European hedgehog, Erinaceus europaeus, using infrared thermography. J. Therm. Biol. 2020, 90, 102574. [Google Scholar] [CrossRef]
- Kastberger, G.; Stachl, R. Infrared imaging technology and biological applications. Behav. Res. Methods Instrum. Comput. 2003, 35, 429–439. [Google Scholar] [CrossRef] [Green Version]
- do Nascimento, G.R.; Nääs, I.A.; Baracho, M.S.; Pereira, D.F.; Neves, D.P. Termografia infravermelho na estimativa de conforto térmico de frangos de corte. Rev. Bras. Eng. Agríc. Ambient. 2014, 18, 658–663. [Google Scholar] [CrossRef] [Green Version]
- Martins, R.F.S.; do Prado Paim, T.; de Abreu Cardoso, C.; Stéfano Lima Dallago, B.; de Melo, C.B.; Louvandini, H.; McManus, C. Mastitis detection in sheep by infrared thermography. Res. Vet. Sci. 2013, 94, 722–724. [Google Scholar] [CrossRef]
- LokeshBabu, D.S.; Jeyakumar, S.; Vasant, P.J.; Sathiyabarathi, M.; Manimaran, A.; Kumaresan, A.; Pushpadass, H.A.; Sivaram, M.; Ramesha, K.P.; Kataktalware, M.A.; et al. Monitoring foot surface temperature using infrared thermal imaging for assessment of hoof health status in cattle: A review. J. Therm. Biol. 2018, 78, 10–21. [Google Scholar] [CrossRef]
- Sommerhäuser, J.; Kloppert, B.; Wolter, W.; Zschöck, M.; Sobiraj, A.; Failing, K. The epidemiology of Staphylococcus aureus infections from subclinical mastitis in dairy cows during a control programme. Vet. Microbiol. 2003, 96, 91–102. [Google Scholar] [CrossRef]
- Pugh, G.; Coates, G.; Adnett, N. Performance indicators and widening participation in UK higher education. High. Educ. Q. 2005, 59, 19–39. [Google Scholar] [CrossRef]
- Jeykumar, M.; Vinodkumar, G.; Bashir, B.; Krovvidi, S. Antibiogram of mastitis pathogens in the milk of crossbred cows in Namakkal district, Tamil Nadu. Vet. World 2013, 6, 354. [Google Scholar] [CrossRef]
- Hovinen, M.; Siivonen, J.; Taponen, S.; Hänninen, L.; Pastell, M.; Aisla, A.-M.; Pyörälä, S. Detection of clinical mastitis with the Help of a thermal camera. J. Dairy Sci. 2008, 91, 4592–4598. [Google Scholar] [CrossRef]
- Jones, B.F.; Plassmann, P. Digital infrared thermal imaging of human skin. IEEE Eng. Med. Biol. Mag. 2002, 21, 41–48. [Google Scholar] [CrossRef] [PubMed]
- McGavin, M.D.; Zachary, J.F. Pathologic Basis of Veterinary Disease, 4th ed.; Elsevier: St. Louis, MO, USA, 2007; pp. 1–560. [Google Scholar]
- Loughmiller, J.A.; Spire, M.F.; Dritz, S.S.; Fenwick, B.W.; Hosni, M.H.; Hogge, S.B. Relationship between mean body surface temperature measured by use of infrared thermography and ambient temperature in clinically normal pigs and pigs inoculated with Actinobacillus pleuropneumoniae. Am. J. Vet. Res. 2001, 62, 676–681. [Google Scholar] [CrossRef] [PubMed]
- Metzner, M.; Sauter-Louis, C.; Seemueller, A.; Petzl, W.; Zerbe, H. Infrared thermography of the udder after experimentally induced Escherichia coli mastitis in cows. Vet. J. 2015, 204, 360–362. [Google Scholar] [CrossRef]
- Polat, B.; Colak, A.; Cengiz, M.; Yanmaz, L.E.; Oral, H.; Bastan, A.; Kaya, S.; Hayirli, A. Sensitivity and specificity of infrared thermography in detection of subclinical mastitis in dairy cows. J. Dairy Sci. 2010, 93, 3525–3532. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alvergnas, M.; Strabel, T.; Rzewuska, K.; Sell-Kubiak, E. Claw disorders in dairy cattle: Effects on production, welfare and farm economics with possible prevention methods. Livest. Sci. 2019, 222, 54–64. [Google Scholar] [CrossRef]
- Tadich, N.; Flor, E.; Green, L. Associations between hoof lesions and locomotion score in 1098 unsound dairy cows. Vet. J. 2010, 184, 60–65. [Google Scholar] [CrossRef] [Green Version]
- Kofler, J. Pathogenesis and treatment of toe lesions in cattle including “nonhealing” toe lesions. Vet. Clin. N. Am. Food Anim. Pract. 2017, 33, 301–328. [Google Scholar] [CrossRef] [PubMed]
- Somers, J.G.C.J.; Schouten, W.G.P.; Frankena, K.; Noordhuizen-Stassen, E.N.; Metz, J.H.M. Development of claw traits and claw lesions in dairy cows kept on different floor systems. J. Dairy Sci. 2005, 88, 110–120. [Google Scholar] [CrossRef] [Green Version]
- Mülling, C.K.W.; Green, L.; Barker, Z.; Scaife, J.; Amory, J.; Speijers, M. Risks factors associated with foot lameness in dairy cattle and suggested approach for lameness reduction. In Proceedings of the XXIV World Buiatrics Congress, Nice, France, 15–19 October 2006; pp. 15–19. [Google Scholar]
- Rainwater-Lovett, K.; Pacheco, J.M.; Packer, C.; Rodriguez, L.L. Detection of foot-and-mouth disease virus infected cattle using infrared thermography. Vet. J. 2009, 180, 317–324. [Google Scholar] [CrossRef]
- Woolhouse, M.; Chase-Topping, M.; Haydon, D.; Friar, J.; Matthews, L.; Hughes, G.; Shaw, D.; Wilesmith, J.; Donaldson, A.; Cornell, S.; et al. Foot-and-mouth disease under control in the UK. Nature 2001, 411, 258–259. [Google Scholar] [CrossRef] [PubMed]
- Jeffcott, L.B.; Rossdale, P.D.; Freestone, J.; Frank, C.J.; Towers-Clark, P.F. An assessment of wastage in thoroughbred racing from conception to 4 years of age. Equine Vet. J. 1982, 14, 185–198. [Google Scholar] [CrossRef] [PubMed]
- Kaneene, J.B.; Ross, W.A.; Miller, R. The Michigan equine monitoring system. II. Frequencies and impact of selected health problems. Prev. Vet. Med. 1997, 29, 277–292. [Google Scholar] [CrossRef]
- Harris, P.; Bailey, S.R.; Elliott, J.; Longland, A. Countermeasures for pasture-associated laminitis in ponies and horses. J. Nutr. 2006, 136, 2114S–2121S. [Google Scholar] [CrossRef]
- Robinson, N.E. Step by painful step: Increasing knowledge about laminitis. Equine Vet. J. 1998, 30, 89–90. [Google Scholar] [CrossRef]
- Douthit, T.L.; Bormann, J.M.; Bello, N.M. Assessing the association between hoof thermography and hoof doppler ultrasonography for the diagnosis of lameness in horses. J. Equine Vet. Sci. 2014, 34, 275–280. [Google Scholar] [CrossRef] [Green Version]
- Hettich, E.; Hinostroza, M.F.; van Schaik, G.; Tadich, N. Factores asociados a la presentación de cojeras en 50 rebaños lecheros de la X. Región, Chile. Arch. Med. Vet. 2007, 39. [Google Scholar] [CrossRef]
- Godoy Pinto, A.; García Liñeiro, A. Infosura en equinos. Av. Cienc. Vet. 2011, 26. [Google Scholar] [CrossRef]
- Douthit, T.L.; Bormann, J.M.; Gradert, K.C.; Lomas, L.W.; DeWitt, S.F.; Kouba, J.M. The impact of endophyte-infected fescue consumption on digital circulation and lameness in the distal thoracic limb of the horse12. J. Anim. Sci. 2012, 90, 3101–3111. [Google Scholar] [CrossRef]
- Seifalian, A.M.; Stansby, G.; Jackson, A.; Howell, K.; Hamilton, G. Comparison of laser doppler perfusion imaging, laser doppler flowmetry, and thermographic imaging for assessment of blood flow in human skin. Eur. J. Vasc. Surg. 1994, 8, 65–69. [Google Scholar] [CrossRef]
- Wienzek, H.; Freise, H.; Giesler, I.; Van Aken, H.K.; Sielenkaemper, A.W. Altered blood flow in terminal vessels after local application of ropivacaine and prilocaine. Reg. Anesth. Pain Med. 2007, 32, 233–239. [Google Scholar] [CrossRef]
- Eddy, A.; Van Hoogmoed, L.; Snyder, J. The role of thermography in the management of equine lameness. Vet. J. 2001, 162, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Turner, T.A. Diagnostic thermography. Vet. Clin. N. Am. Equine Pract. 2001, 17, 95–114. [Google Scholar] [CrossRef]
- Bergmann, S.; Ziegler, N.; Bartels, T.; Hübel, J.; Schumacher, C.; Rauch, E.; Brandl, S.; Bender, A.; Casalicchio, G.; Krautwald-Junghanns, M.-E.; et al. Prevalence and severity of foot pad alterations in German turkey poults during the early rearing phase. Poult. Sci. 2013, 92, 1171–1176. [Google Scholar] [CrossRef]
- Moe, R.O.; Flø, A.; Bohlin, J.; Vasdal, G.; Erlandsen, H.; Guneriussen, E.; Sjökvist, E.C.; Stubsjøen, S.M. Experimental factors affecting the within- and between-individual variation of plantar foot surface temperatures in turkeys (Meleagris gallopovo) recorded with infrared thermography. Infrared Phys. Technol. 2018, 92, 381–386. [Google Scholar] [CrossRef] [Green Version]
- Wilcox, C.S.; Patterson, J.; Cheng, H.W. Use of thermography to screen for subclinical bumblefoot in poultry. Poult. Sci. 2009, 88, 1176–1180. [Google Scholar] [CrossRef]
- Penguin Taxon Advisory. Penguin Husbandry Manual, 3rd ed.; AZA-Association of Zoos and Aquariums: Silver Spring, MD, USA, 2005. [Google Scholar]
- Mayne, R.K. A review of the aetiology and possible causative factors of foot pad dermatitis in growing turkeys and broilers. Worlds Poult. Sci. J. 2005, 61, 256–267. [Google Scholar] [CrossRef]
- Mayne, R.K.; Hocking, P.M.; Else, R.W. Foot pad dermatitis develops at an early age in commercial turkeys. Br. Poult. Sci. 2006, 47, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Jericho, K.W.F.; Kozub, G.C. Experimental infectious respiratory disease in groups of calves: Lobar distribution, variance, and sample-size requirements for vaccine evaluation. Can. J. Vet. Res. 2004, 68, 118–127. [Google Scholar] [PubMed]
- Lopez, A.; Martinson, S.A. Respiratory system, mediastinum and pleurae. In Pathologic Basis of Veterinary Disease; Zachary, J.F., Ed.; Elsevier: St. Louis, MO, USA, 2017; pp. 471–560. [Google Scholar]
- Taylor, J.D.; Fulton, R.W.; Lehenbauer, T.W.; Step, D.L.; Confer, A.W. The epidemiology of bovine respiratory disease: What is the evidence for preventive measures? Can. Vet. J. 2010, 51, 1351–1359. [Google Scholar]
- Mellon, M.; Benbrook, C.; Lutz Benbrook, K. Hogging It! Estimates of Antimicrobial Abuse in Livestock; Union of Concerned Scientists: Cambridge, MA, USA, 2001. [Google Scholar]
- Schaefer, A.L.; Cook, N.; Tessaro, S.V.; Deregt, D.; Desroches, G.; Dubeski, P.L.; Tong, A.K.W.; Godson, D.L. Early detection and prediction of infection using infrared thermography. Can. J. Anim. Sci. 2004, 84, 73–80. [Google Scholar] [CrossRef]
- Betancur-Hurtado, C.; Castañeda-Ternera, J.; Gonzalez, M. Immunopathology of bovine respiratory complex in calves in the Municipality of Monteria, Colombia. Rev. Cient. Fac. Cienc. Vet. Univ. Zulia 2017, 27, 95–102. [Google Scholar]
- Humblet, M.-F.; Coghe, J.; Lekeux, P.; Godeau, J.-M. Acute phase proteins assessment for an early selection of treatments in growing calves suffering from bronchopneumonia under field conditions. Res. Vet. Sci. 2004, 77, 41–47. [Google Scholar] [CrossRef] [PubMed]
- Schaefer, A.L.; Cook, N.J.; Church, J.S.; Basarab, J.; Perry, B.; Miller, C.; Tong, A.K.W. The use of infrared thermography as an early indicator of bovine respiratory disease complex in calves. Res. Vet. Sci. 2007, 83, 376–384. [Google Scholar] [CrossRef] [PubMed]
- Thrusfield, M. Diagnostic Testing. Veterinary Epidemiology, 2nd ed.; Blackwell Sci. Ltd.: Oxford, UK, 1995. [Google Scholar]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mota-Rojas, D.; Wang, D.; Titto, C.G.; Gómez-Prado, J.; Carvajal-de la Fuente, V.; Ghezzi, M.; Boscato-Funes, L.; Barrios-García, H.; Torres-Bernal, F.; Casas-Alvarado, A.; et al. Pathophysiology of Fever and Application of Infrared Thermography (IRT) in the Detection of Sick Domestic Animals: Recent Advances. Animals 2021, 11, 2316. https://doi.org/10.3390/ani11082316
Mota-Rojas D, Wang D, Titto CG, Gómez-Prado J, Carvajal-de la Fuente V, Ghezzi M, Boscato-Funes L, Barrios-García H, Torres-Bernal F, Casas-Alvarado A, et al. Pathophysiology of Fever and Application of Infrared Thermography (IRT) in the Detection of Sick Domestic Animals: Recent Advances. Animals. 2021; 11(8):2316. https://doi.org/10.3390/ani11082316
Chicago/Turabian StyleMota-Rojas, Daniel, Dehua Wang, Cristiane Gonçalves Titto, Jocelyn Gómez-Prado, Verónica Carvajal-de la Fuente, Marcelo Ghezzi, Luciano Boscato-Funes, Hugo Barrios-García, Fabiola Torres-Bernal, Alejandro Casas-Alvarado, and et al. 2021. "Pathophysiology of Fever and Application of Infrared Thermography (IRT) in the Detection of Sick Domestic Animals: Recent Advances" Animals 11, no. 8: 2316. https://doi.org/10.3390/ani11082316
APA StyleMota-Rojas, D., Wang, D., Titto, C. G., Gómez-Prado, J., Carvajal-de la Fuente, V., Ghezzi, M., Boscato-Funes, L., Barrios-García, H., Torres-Bernal, F., Casas-Alvarado, A., & Martínez-Burnes, J. (2021). Pathophysiology of Fever and Application of Infrared Thermography (IRT) in the Detection of Sick Domestic Animals: Recent Advances. Animals, 11(8), 2316. https://doi.org/10.3390/ani11082316