Author Contributions
R.T.d.A.S.: Investigation, Methodology, Data curation, Writing—Original draft; M.V.F.d.S.: Conceptualization, Validation, Resources, Project administration; M.V.d.C.: Methodology, Data curation, Validation, Supervision; G.D.G.: Writing—Review and Editing, Visualization, Supervision; V.J.d.S.: Writing—Review and Editing; A.C.L.d.M.: Validation, Resources, Writing—Review and Editing, Project administration; J.P.M.: Writing—Review and Editing; R.E.P.R.: Investigation, Methodology, Writing—Review; J.C.B.D.J.: Writing—Review and Editing, Supervision. All authors have read and agreed to the published version of the manuscript.
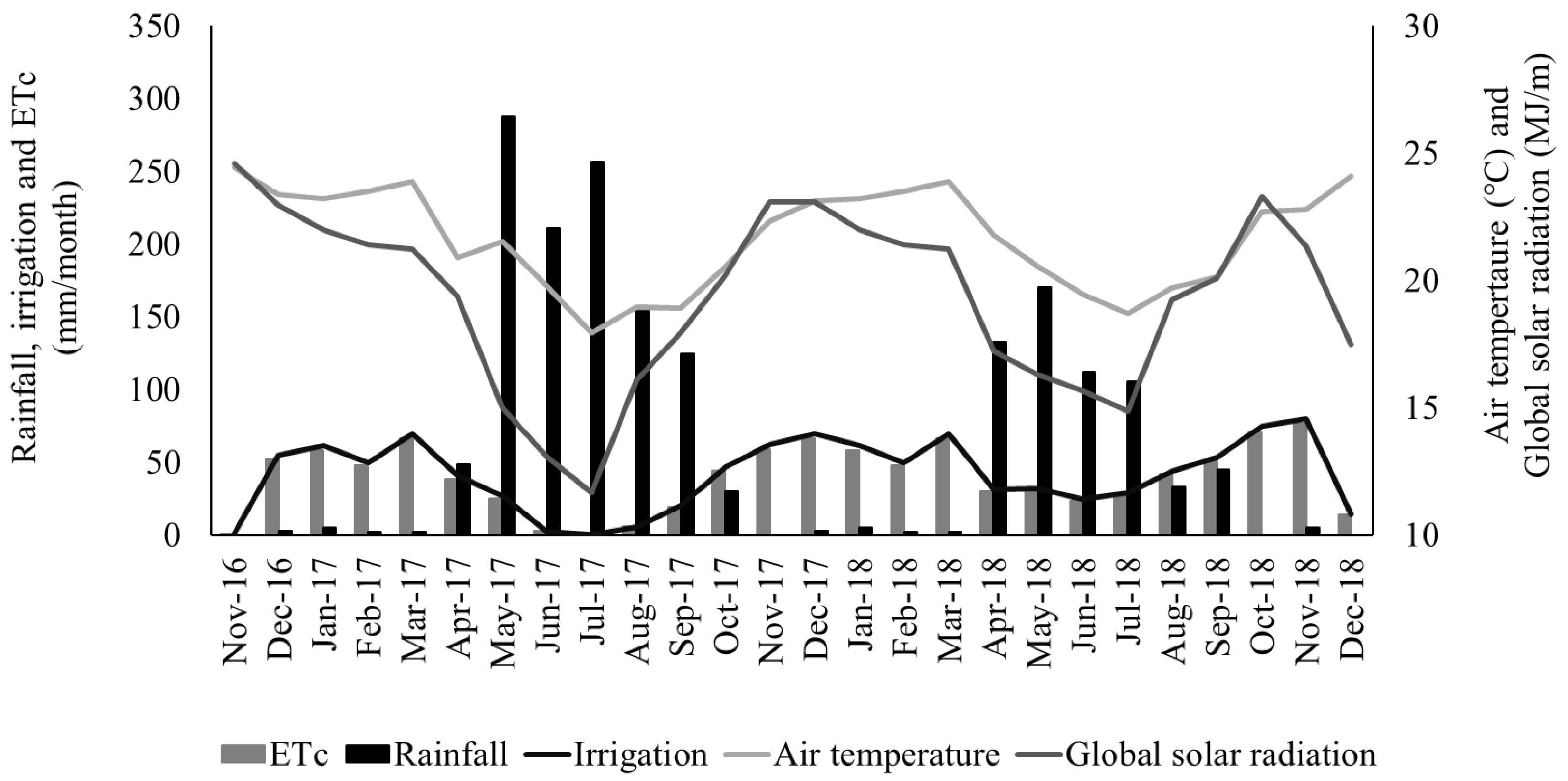
Figure 1.
Rainfall, crop evapotranspiration (ETc), daily irrigation, air temperature, and global solar radiation recorded during the experimental period at Garanhuns, Pernambuco, Brazil.
Figure 1.
Rainfall, crop evapotranspiration (ETc), daily irrigation, air temperature, and global solar radiation recorded during the experimental period at Garanhuns, Pernambuco, Brazil.
Figure 2.
Herbage accumulation rate (HAR) (kg dry matter/ha per day) as affected by irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
Figure 2.
Herbage accumulation rate (HAR) (kg dry matter/ha per day) as affected by irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
Figure 3.
Stem neutral detergent fiber (NDF) (g/kg dry matter) as affected by genotype for elephantgrass at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
Figure 3.
Stem neutral detergent fiber (NDF) (g/kg dry matter) as affected by genotype for elephantgrass at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
Figure 4.
Stem carbohydrate fraction B2 (g/kg carbohydrate) as affected by season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
Figure 4.
Stem carbohydrate fraction B2 (g/kg carbohydrate) as affected by season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
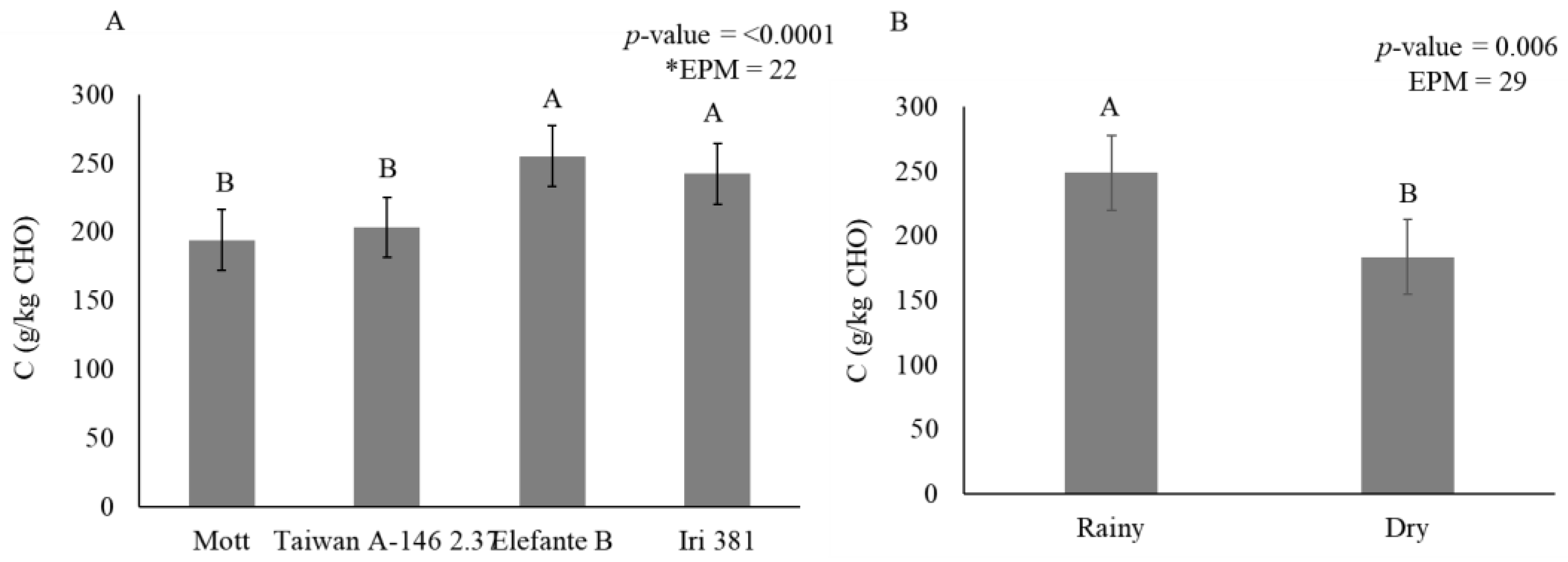
Figure 5.
Stem carbohydrate fraction C (g/kg carbohydrate) as affected by genotype (A) and season (B) for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
Figure 5.
Stem carbohydrate fraction C (g/kg carbohydrate) as affected by genotype (A) and season (B) for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil. *EPM = Standard error of the means.
Table 1.
Herbage accumulation rate (HAR) and stem accumulation rate (SAR; kg dry matter/ha per day), as affected by the interaction between genotype and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 1.
Herbage accumulation rate (HAR) and stem accumulation rate (SAR; kg dry matter/ha per day), as affected by the interaction between genotype and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Rainy | Dry | SEM 2 | p-Value |
|---|
| HAR |
| Mott | 122 Ac 1 | 55 Bab | 23 | 0.007 |
| Taiwan A-146 2.37 | 146 Aab | 39 Bb |
| Elefante B | 124 Acb | 49 Bab |
| Iri 381 | 150 Aa | 63 Ba |
| SAR |
| Mott | 57 Ac | 19 Ba | 8 | 0.001 |
| Taiwan A-146 2.37 | 71 Ab | 17 Ba |
| Elefante B | 89 Aab | 22 Ba |
| Iri 381 | 90 Aa | 30 Ba |
Table 2.
Leaf accumulation rate (LAR; kg dry matter/ha per day) as affected by the interaction between genotype and irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 2.
Leaf accumulation rate (LAR; kg dry matter/ha per day) as affected by the interaction between genotype and irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Irrigated | Rainfed | SEM 2 | p-Value |
|---|
| Mott | 66 Aa 1 | 36 Ba | 9 | 0.048 |
| Taiwan A-146 2.37 | 49 Ab | 30 Ba |
| Elefante B | 56 Aab | 25 Ba |
| Iri 381 | 59 Aab | 34 Ba |
Table 3.
Leaf accumulation rate (LAR; kg dry matter/ha per day) and leaf:stem (L:S) ratio, as affected by the interaction between irrigation and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 3.
Leaf accumulation rate (LAR; kg dry matter/ha per day) and leaf:stem (L:S) ratio, as affected by the interaction between irrigation and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Season | Irrigated | Rainfed | SEM 2 | p-Value |
|---|
| LAR |
| Rainy | 69 Aa 1 | 59 Aa | 12 | 0.005 |
| Dry | 55 Aa | 14 Bb |
| L:S |
| Rainy | 0.84 Aa | 0.77 Ab | 0.12 | <0.0001 |
| Dry | 1.32 Ba | 2.01 Aa |
Table 4.
Leaf crude protein (CP), neutral detergent fiber (NDF), acid detergent fiber (ADF), and in vitro dry matter digestibility (IVDDM) (g/kg dry matter) as affected by the interaction between irrigation, genotype, and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 4.
Leaf crude protein (CP), neutral detergent fiber (NDF), acid detergent fiber (ADF), and in vitro dry matter digestibility (IVDDM) (g/kg dry matter) as affected by the interaction between irrigation, genotype, and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Rainy | Dry | SEM 2 | p-Value |
|---|
| Irrigated | Rainfed | Irrigated | Rainfed |
|---|
| CP |
| Mott | 96 Aaα 1 | 88 Aaβ | 89 Baα | 140 Aaα | 3 | 0.005 |
| Taiwan A-146 2.37 | 86 Aabα | 82 Aabβ | 83 Babα | 109 Abα |
| Elefante B | 73 Abα | 76 Abβ | 78 Bbα | 107 Abα |
| Iri 381 | 73 Abα | 75 Abβ | 79 Bbα | 96 Abα |
| NDF |
| Mott | 609 Abα | 600 Abα | 610 Abα | 537 Bbβ | 8 | 0.0038 |
| Taiwan A-146 2.37 | 683 Aaα | 688 Aaα | 687 Aaα | 626 Baβ |
| Elefante B | 684 Aaα | 697 Aaα | 686 Aaα | 612 Baβ |
| Iri 381 | 697 Aaα | 670 Aaα | 690 Aaα | 611 Baβ |
| ADF |
| Mott | 297 Abα | 296 Abα | 310 Abα | 280 Bbβ | 4 | 0.016 |
| Taiwan A-146 2.37 | 320 Aaα | 338 Aaα | 338 Aaα | 336 Aaα |
| Elefante B | 331 Aaα | 348 Aaα | 350 Aaα | 332 Aaα |
| Iri 381 | 345 Aaα | 357 Aaα | 324 Aabα | 324 Aaβ |
| IVDDM |
| Mott | 658 Aaα | 697 Aaβ | 686 Baα | 726 Aaα | 21 | 0.0004 |
| Taiwan A-146 2.37 | 595 Aaα | 573 Aaβ | 593 Baα | 695 Aaα |
| Elefante B | 542 Abα | 548 Abβ | 537 Bbα | 610 Abα |
| Iri 381 | 563 Abα | 522 Abβ | 555 Bbα | 619 Abα |
Table 5.
Leaf lignin (g/kg dry matter) as affected by the interaction between irrigation and genotype for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 5.
Leaf lignin (g/kg dry matter) as affected by the interaction between irrigation and genotype for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Irrigated | Rainfed | SEM 2 | p-Value |
|---|
| Mott | 71 Ab 1 | 53 Bb | 3 | 0.038 |
| Taiwan A-146 2.37 | 99 Aab | 64 Bab |
| Elefante B | 112 Aa | 77 Ba |
| Iri 381 | 116 Aa | 88 Ba |
Table 6.
Stem crude protein (CP), acid detergent fiber (ADF), and lignin concentration (g/kg dry matter), as affected by the interaction between irrigation, genotype, and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 6.
Stem crude protein (CP), acid detergent fiber (ADF), and lignin concentration (g/kg dry matter), as affected by the interaction between irrigation, genotype, and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Rainy | Dry | SEM 2 | p-Value |
|---|
| Irrigated | Rainfed | Irrigated | Rainfed |
|---|
| CP |
| Mott | 53 Aaα 1 | 49 Aaβ | 55 Baα | 69 Aaα | 4 | <0.0001 |
| Taiwan A-146 2.37 | 40 Abα | 40 Abβ | 47 Bbα | 67 Aaα |
| Elefante B | 43 Abα | 41 Abβ | 42 Bbα | 52 Abα |
| Iri 381 | 37 Acα | 32 Acβ | 36 Bcα | 52 Abα |
| ADF |
| Mott | 367 Abα | 360 Abα | 377 Abα | 308 Bbβ | 10 | 0.012 |
| Taiwan A-146 2.37 | 433 Aaα | 423 Aaα | 427 Aaα | 362 Baβ |
| Elefante B | 415 Aaα | 401 Aaα | 416 Aaα | 353 Baβ |
| Iri 381 | 448 Aaα | 434 Aaα | 443 Aaα | 376 Baβ |
| Lignin |
| Mott | 112 Abα | 120 Abα | 119 Abα | 99 Bbβ | 9 | 0.001 |
| Taiwan A-146 2.37 | 147 Aaα | 146 Aaα | 148 Aaα | 125 Baβ |
| Elefante B | 145 Aaα | 147 Aaα | 132 Aaα | 122 Baβ |
| Iri 381 | 144 Aaα | 141 Aaα | 143 Aaα | 129 Baβ |
Table 7.
Stem neutral detergent fiber (NDF) and in vitro digestible dry matter (IVDDM) (g/kg dry matter), as affected by the interaction between irrigation and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 7.
Stem neutral detergent fiber (NDF) and in vitro digestible dry matter (IVDDM) (g/kg dry matter), as affected by the interaction between irrigation and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Season | Irrigated | Rainfed | SEM 2 | p-Value |
|---|
| NDF |
| Rainy | 723 Aa 1 | 709 Aa | 10 | <0.0001 |
| Dry | 706 Aa | 649 Bb |
| IVDDM |
| Rainy | 365 Aa | 378 Ab | 8 | 0.0002 |
| Dry | 364 Ba | 411 Aa |
Table 8.
Stem in vitro dry matter digestibility (g/kg dry matter) as affected by the interaction between genotype and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 8.
Stem in vitro dry matter digestibility (g/kg dry matter) as affected by the interaction between genotype and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Rainy | Dry | SEM 2 | p-Value |
|---|
| Mott | 367 Ba 1 | 416 Aa | 11 | 0.0002 |
| Taiwan A-146 2.37 | 379 Ba | 415 Aa |
| Elefante B | 317 Bb | 366 Ab |
| Iri 381 | 326 Bb | 378 Ab |
Table 9.
Leaf protein fractionation (g/kg crude protein) as affected by the interaction between irrigation, genotype, and season for elephantgrass at Garanhuns, Pernambuco, Brazil.
Table 9.
Leaf protein fractionation (g/kg crude protein) as affected by the interaction between irrigation, genotype, and season for elephantgrass at Garanhuns, Pernambuco, Brazil.
| Genotype | Rainy | Dry | SEM 2 | p-Value |
|---|
| Irrigated | Rainfed | Irrigated | Rainfed |
|---|
| A |
| Mott | 160 Aaα 1 | 170 Aaβ | 160 Baα | 200 Aaα | 7 | 0.002 |
| Taiwan A-146 2.37 | 130 Aabα | 120 Abβ | 120 Babα | 170 Aabα |
| Elefante B | 100 Abα | 90 Acβ | 90 Bbα | 120 Abα |
| Iri 381 | 100 Abα | 70 Acβ | 70 Bbα | 140 Abα |
| B1 + B2 |
| Mott | 570 Aaα | 560 Aaβ | 590 Aaα | 600 Aaα | 12 | <0.0001 |
| Taiwan A-146 2.37 | 570 Aaα | 570 Aaα | 580 Aaα | 550 Aaα |
| Elefante B | 540 Aaα | 540 Aaβ | 560 Baα | 590 Aaα |
| Iri 381 | 530 Aaα | 560 Aaα | 570 Aaα | 560 Aaα |
| B3 |
| Mott | 210 Abα | 220 Abα | 200 Abα | 160 Bbβ | 7 | <0.0001 |
| Taiwan A-146 2.37 | 240 Aabα | 250 Aabα | 240 Aaα | 240 Aaα |
| Elefante B | 280 Aaα | 290 Aaα | 260 Aaα | 230 Baβ |
| Iri 381 | 280 Aaα | 280 Aaα | 270 Aaα | 240 Baβ |
| C |
| Mott | 60 Abα | 50 Abα | 50 Abα | 40 Bbβ | 2 | 0.0006 |
| Taiwan A-146 2.37 | 60 Abα | 60 Abα | 60 Abα | 40 Bbβ |
| Elefante B | 80 Aaα | 80 Aaα | 90 Aaα | 60 Baβ |
| Iri 381 | 90 Aaα | 90 Aaα | 90 Aaα | 60 Baβ |
Table 10.
Stem protein fractionations (g/kg crude protein) as affected by the interaction between irrigation, genotype, and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 10.
Stem protein fractionations (g/kg crude protein) as affected by the interaction between irrigation, genotype, and season for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Rainy | Dry | SEM 2 | p-Value |
|---|
| Irrigated | Rainfed | Irrigated | Rainfed |
|---|
| A |
| Mott | 110 Aaα 1 | 120 Aaβ | 110 Baα | 150 Aaα | 8 | 0.001 |
| Taiwan A-146 2.37 | 120 Aaα | 110 Aaβ | 110 Baα | 160 Aaα |
| Elefante B | 80 Aaα | 80 Aaβ | 90 Baα | 130 Aaα |
| Iri 381 | 90 Aaα | 100 Aaβ | 90 Baα | 140 Aaα |
| B1 + B2 |
| Mott | 360 Aaα | 360 Aaβ | 360 Baα | 440 Aaα | 7 | <0.0001 |
| Taiwan A-146 2.37 | 300 Aaα | 290 Aaβ | 350 Baα | 410 Aaα |
| Elefante B | 230 Abα | 230 Abβ | 220 Bbα | 360 Abα |
| Iri 381 | 260 Abα | 240 Abβ | 250 Bbα | 340 Abα |
| B3 |
| Mott | 410 Abα | 430 Abα | 420 Abα | 330 Bbβ | 6 | <0.0001 |
| Taiwan A-146 2.37 | 450 Abα | 450 Abα | 400 Abα | 340 Bbα |
| Elefante B | 550 Aaα | 540 Aaα | 560 Aaα | 390 Baβ |
| Iri 381 | 500 Aaα | 520 Aaα | 510 Aaα | 420 Baβ |
| C |
| Mott | 120 Aaα | 90 Abα | 110 Aaα | 80 Bbβ | 2 | 0.001 |
| Taiwan A-146 2.37 | 130 Aaα | 150 Aaα | 140 Aaα | 90 Babβ |
| Elefante B | 140 Aaα | 150 Aaα | 130 Aaα | 120 Baβ |
| Iri 381 | 150 Aaα | 140 Aaα | 150 Aaα | 100 Baβ |
Table 11.
Leaf carbohydrate fractionation (g/kg carbohydrate) as affected by the interaction between genotype and irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 11.
Leaf carbohydrate fractionation (g/kg carbohydrate) as affected by the interaction between genotype and irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Irrigated | Rainfed | SEM 2 | p-Value |
|---|
| A + B1 |
| Mott | 390 Ba 1 | 460 Aa | 6 | 0.001 |
| Taiwan A-146 2.37 | 330 Bb | 420 Ab |
| Elefante B | 335 Bb | 400 Ab |
| Iri 381 | 360 Bb | 410 Ab |
| B2 |
| Mott | 460 Ab | 430 Bb | 7 | <0.0001 |
| Taiwan A-146 2.37 | 500 Aa | 450 Bab |
| Elefante B | 505 Aa | 480 Ba |
| Iri 381 | 480 Aa | 460 Ba |
| C |
| Mott | 150 Aa | 110 Ba | 1 | <0.0001 |
| Taiwan A-146 2.37 | 170 Aa | 130 Ba |
| Elefante B | 160 Aa | 120 Ba |
| Iri 381 | 160 Aa | 130 Ba |
Table 12.
Stem carbohydrate fractions A + B1 and B2 (g/kg carbohydrate) as affected by the interaction between genotype and irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
Table 12.
Stem carbohydrate fractions A + B1 and B2 (g/kg carbohydrate) as affected by the interaction between genotype and irrigation for elephantgrass genotypes at Garanhuns, Pernambuco, Brazil.
| Genotype | Irrigated | Rainfed | SEM 2 | p-Value |
|---|
| A + B1 |
| Mott | 262 Ba 1 | 353 Aa | 5 | <0.0001 |
| Taiwan A-146 2.37 | 217 Bb | 270 Ab |
| Elefante B | 218 Bb | 253 Ab |
| Iri 381 | 218 Bb | 253 Ab |
| B2 |
| Mott | 505 Ab | 415 Bb | 13 | <0.0001 |
| Taiwan A-146 2.37 | 491 Ab | 409 Bb |
| Elefante B | 579 Aa | 483 Ba |
| Iri 381 | 564 Aa | 491 Ba |


 ,
, 





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}