
Herpesvirus Infection in a Breeding Population of Two Coexisting Strix Owls

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Field Sampling of Strix Owls
2.2. Field Sampling of Mice
2.3. DNA Extraction and PCR of a DNA Polymerase Gene Region Using Herpesvirus Consensus Primers
2.4. Detection, Sequencing, and Phylogenetic Analysis of PCR Products
3. Results
3.1. Herpesvirus Detection in Owls and Yellow-Necked Mouse Populations
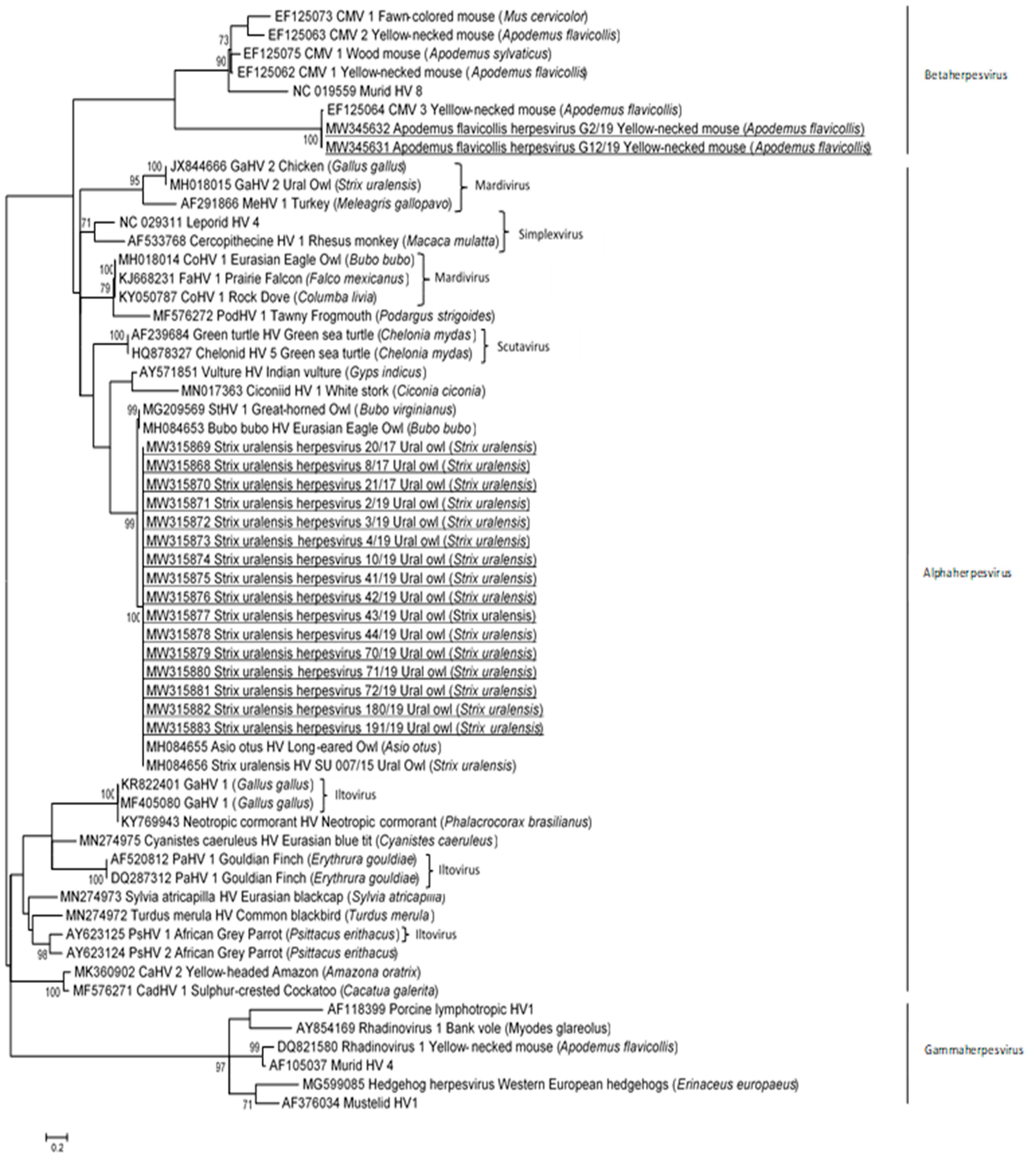
3.2. Phylogenetic and Sequence Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Newton, I. Population Limitation in Birds, 1st ed.; Academic Press: London, UK, 1998; 597p. [Google Scholar]
- Cunningham, A.A.; Daszak, P.; Wood, J.L.N. One Health, emerging infectious diseases and wildlife: Two decades of progress? Philos. Trans. R. Soc. B 2017, 372, 20160167. [Google Scholar] [CrossRef] [Green Version]
- Movalli, P.; Krone, O.; Osborn, D.; Pain, D. Monitoring contaminants, emerging infectious diseases and environmental change with raptors, and links to human health. Bird Study 2018, 65, S96–S109. [Google Scholar] [CrossRef] [Green Version]
- Korpimäki, E.; Hakkarainen, H.; Ilmonen, P.; Wiehn, J. Detrimental effects of blood parasites on parental effort and reproductive success of Tengmalm’s owls (Aegolius funereus) and Eurasian kestrels (Falco tinnunculus). In Ecology and Conservation of Owls; Newton, I., Kavanagh, R., Olson, J., Taylor, I.R., Eds.; CSIRO Publishing: Collingwood, VIC, Australia, 2002; pp. 66–71. [Google Scholar]
- Molina-López, R.A.; Valverdú, N.; Martin, M.; Mateu, E.; Obon, E.; Cerdà-Cuéllar, M.; Darwich, L. Wild raptors as carriers of antimicrobial resistant Salmonella and Campylobacter strains. Vet. Rec. 2011, 168, 565. [Google Scholar] [CrossRef] [PubMed]
- Karell, P.; Bensch, S.; Ahola, K.; Asghar, M. Pale and dark morphs of tawny owls show different patterns of telomere dynamics in relation to disease status. Proc. R. Soc. B 2017, 284, 2017112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gailbreath, K.L.; Oaks, J.L. Herpesviral inclusion body disease in owls and falcons is caused by the pigeon herpesvirus (Columbid herpesvirus 1). J. Wildl. Dis. 2008, 44, 427–433. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinkerton, M.E.; Wellehan, J.F.X.; Johnson, A.J.; Childress, A.L.; Fitzgerald, S.D.; Kinsel, M.J. Columbid herpesvirus-1 in two Cooper’s hawks (Accipiter cooperii) with fatal inclusion body disease. J. Wildl. Dis. 2008, 44, 622–628. [Google Scholar] [CrossRef] [Green Version]
- de Thoisy, B.; Lavergne, A.; Semelin, J.; Pouliquen, J.F.; Blanchard, F.; Hansen, E.; Lacoste, V. Outbreaks of disease possibly due to a natural avian herpesvirus infection in a colony of young magnificent frigatebirds (Fregata magnificens) in French Guiana. J. Wildl. Dis. 2009, 45, 802–807. [Google Scholar] [CrossRef] [Green Version]
- Amery-Gale, J.; Hartley, C.A.; Vaz, P.K.; Marenda, M.S.; Owens, J.; Eden, P.A.; Devlin, J.M. Avian viral surveillance in Victoria, Australia, and detection of two novel avian herpesviruses. PLoS ONE 2018, 13, e0194457. [Google Scholar] [CrossRef] [Green Version]
- Zlabravec, Z.; Krapez, U.; Slavec, B.; Vrezec, A.; Rojs, O.Z.; Racnik, J. Detection and phylogenetic analysis of herpesviruses detected in wild owls in Slovenia. Avian Dis. 2018, 62, 397–403. [Google Scholar] [CrossRef] [PubMed]
- Bürki, F.; Burtscher, H.; Sibalin, M. Herpesvirus strigis: A new avian herpesvirus. Arch. F Virusforsch. 1973, 43, 14–24. [Google Scholar] [CrossRef]
- Kaleta, E.F.; Docherty, D. Avian herpesviruses. In Infectious Disease of Wild Birds; Thomas, N., Hunter, D., Atkinson, C., Eds.; Blackwell Publishers: Ames, IA, USA, 2007; pp. 63–86. [Google Scholar]
- International Committee on Taxonomy of Viruses (ICTV). Available online: https://ictv.global/taxonomy/ (accessed on 16 August 2021).
- Ritchie, B.W. Avian Viruses: Function and Control; Wingers Publishing: Lake Worth, FL, USA, 1995; 525p. [Google Scholar]
- Gleeson, M.D.; Moore, B.A.; Edwards, S.G.; Stevens, S.; Childress, A.L.; Wellehan, J.F.X.; Robertson, J.; Murphy, C.J.; Hawkins, M.G.; Paul-Murphy, J. A novel herpesvirus associated with chronic superficial keratitis and proliferative conjunctivitis in a great horned owl (Bubo virginianus). Vet. Ophthalmol. 2018, 22, 67–75. [Google Scholar] [CrossRef]
- Barkhoff, M. Die Krankheiten des Uhus (Bubo bubo) und Ihre Bedeutung für Die Wiedereinbürgerung in der Bundesrepublik Deutschland. Ph.D. Dissertation, Justus Liebig-Universität Giessen, Giessen, Germany, 1987. [Google Scholar]
- Burtscher, H.; Sibalin, M. Herpesvirus strigis: Host spectrum and distribution in infected owls. J. Wildl. Dis. 1975, 11, 164–169. [Google Scholar] [CrossRef]
- Heidenreich, M.; Kaleta, E.F. Hepatosplenitis infectiosa strigum: Beitrag zum Wirtsspektrum und zur Übertragbarkeit des Eulen-Herpesvirus. Fortschr. Der Vet. 1978, 28, 198–203. [Google Scholar]
- Zarrouk, K.; Piret, J.; Boivin, G. Herpesvirus DNA polymerases: Structures, functions and inhibitors. Virus Res. 2017, 15, 177–192. [Google Scholar] [CrossRef]
- VanDevanter, D.R.; Warrener, P.; Bennett, L.; Schultz, E.R.; Coulter, S.; Garber, R.L.; Rose, T.M. Detection and analysis of diverse herpesviral species by consensus primer PCR. J. Clin. Microbiol. 1996, 34, 1666–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ehlers, B.; Borchers, K.; Grund, C.; Frolich, K.; Ludwig, H.; Buhk, H.J. Detection of new DNA polymerase genes of known and potentially novel herpesviruses by PCR with degenerate and deoxyinosine-substituted primers. Virus Genes 1999, 18, 211–220. [Google Scholar] [CrossRef]
- Kaleta, E.F.; Lierz, M.W. Herpesviruses of free-living and pet birds. In A Laboratory Manual for the Isolation, Identification, and Characterization of Avian Pathogens, 6th ed.; Williams, S.M., Dufour-Zavala, L., Jackwood, M.W., Lee, M.D., Lupiani, B., Reed, W.M., Spackman, E., Woolcock, P.R., Eds.; American Association of Avian Pathologists: Athens, GA, USA, 2016; pp. 221–232. [Google Scholar]
- Phalen, D.N.; Alvarado, C.; Grillo, V.; Mason, P.; Dobson, E.; Holz, P. Prevalence of columbid herpesvirus infection in feral pigeons from New South Wales and Victoria, Australia, with spillover into a wild powerful owl (Ninox strenua). J. Wildl. Dis. 2017, 53, 543–551. [Google Scholar] [CrossRef]
- Thomas, N.J.; Hunter, D.B.; Atkinson, C.T. (Eds.) Infectious Diseases of Wild Birds; Wiley-Blackwell: New York, NY, USA, 2007; 498p. [Google Scholar]
- Vrezec, A.; Tome, D. Altitudinal segregation between Ural owl Strix uralensis and tawny owl S. aluco: Evidence for competitive exclusion in raptorial birds. Bird Study 2004, 51, 264–269. [Google Scholar] [CrossRef] [Green Version]
- Mikkola, H. Owls of Europe, 1st ed.; T & A.D. Poyser: London, UK, 1983; 397p. [Google Scholar]
- Pavon-Jordan, D.; Karell, P.; Ahola, K.; Kolunen, H.; Pietiäinen, H.; Karstinen, T.; Brommer, J.E. Environmental correlates of annual survival differ between two ecologically similar and congeneric owls. IBIS 2013, 155, 823–834. [Google Scholar] [CrossRef]
- Vrh, P.; Vrezec, A. Interspecific territorial vocal activity of the Ural (Strix uralensis) towards tawny owl (Strix aluco), sympatric owl competitor: A playback experiment. Razpr. IV Razreda SAZU 2006, 47, 99–105. [Google Scholar]
- Lundberg, A. Are the Ural owl Strix uralensis and the tawny owl S. aluco parapatric in Scandinavia? Ornis Scand. 1980, 11, 116–120. [Google Scholar] [CrossRef]
- Stürzer, J.S. Bestandsentwicklung und Nahrungsökologie von Habichtkauz Strix uralensis und Waldkauz Strix aluco im Nationalpark Bayerischer Wald. Ornithol. Anz. 1998, 37, 109–119. [Google Scholar]
- Vrezec, A.; Saurola, P.; Avotins, A.; Kocijančič, S.; Sulkava, S. A comparative study of Ural owl Strix uralensis breeding season diet within its European breeding range, derived from nest box monitoring schemes. Bird Study 2018, 65, S85–S95. [Google Scholar] [CrossRef] [Green Version]
- Vrezec, A. Breeding density and altitudinal distribution of the Ural, tawny, and boreal owls in North Dinaric Alps (central Slovenia). J. Raptor Res. 2003, 37, 55–62. [Google Scholar]
- Sotenšek, B. Food Niches of Two Sympatric Owl Species Ural Owl (Strix uralensis) and Tawny Owl (Strix aluco) during Breeding Season. Bachelor’s Thesis, University of Ljubljana, Ljubljana, Slovenia, 2012. (In Slovenian). [Google Scholar]
- Skok, J.; Kryštufek, B. Dormice in small mammal assemblages in a mixed southern European forest. Peckiana 2012, 8, 69–75. [Google Scholar]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Boil. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [Green Version]
- llmonen, P.; Hakkarainen, H.; Koivunen, V.; Korpimäki, E.; Mullie, A.; Shutler, D. Parental effort and blood parasitism in Tengmalm’s owl: Effects of natural and experimental variation in food abundance. Oikos 1999, 86, 79–86. [Google Scholar] [CrossRef] [Green Version]
- Žlabravec, Z.; Trilar, T.; Slavec, B.; Krapež, U.; Vrezec, A.; Zorman Rojs, O.; Račnik, J. Detection of herpesviruses in passerine birds captured during autumn migration in Slovenia. J. Wildl. Dis. 2021, 57, 368–375. [Google Scholar] [CrossRef]
- Ratajc, U.; Breskvar, M.; Džeroski, S.; Vrezec, A. Differential responses of coexisting owls to annual small mammal population fluctuations in temperate mixed forest. Ibis. under review.
- Kaleta, E.F. Herpesviruses of birds: A review. Avian Pathol. 1990, 19, 193–211. [Google Scholar] [CrossRef]
- Wink, M.; El-Sayed, A.A.; Sauer-Gürth, H.; Gonzalez, J. Molecular phylogeny of owls (Strigiformes) inferred from DNA sequences of the mitochondrial cytochrome b and the nuclear RAG-1 gene. Ardea 2009, 97, 581–591. [Google Scholar] [CrossRef] [Green Version]
- Kang, H.; Lia, B.; Ma, X.; Xua, Y. Evolutionary progression of mitochondrial gene rearrangements and phylogenetic relationships in Strigidae (Strigiformes). Gene 2018, 674, 8–14. [Google Scholar] [CrossRef]
- Petty, S.J.; Saurola, P. Strix aluco: Tawny Owl. In EBCC Atlas of European Breeding Birds: Their Distribution and Abundance; Hagemeijer, W.J.M., Blair, M.J., Brouwer, P., Eds.; T & AD Poyser: London, UK, 1997; pp. 410–411. [Google Scholar]
- Pietiäinen, H.; Saurola, P. Strix uralensis: Ural Owl. In EBCC Atlas of European Breeding Birds: Their Distribution and Abundance; Hagemeijer, W.J.M., Blair, M.J., Brouwer, P., Eds.; T & AD Poyser: London, UK, 1997; pp. 412–413. [Google Scholar]
- Huntley, B.; Green, R.E.; Collingham, Y.C.; Willis, S.G. A Climatic Atlas of European Breeding Birds; Lynx Edicions: Barcelona, Spain, 2007; 521p. [Google Scholar]
- Estep, R.D.; Messaoudi, I.; Wong, S.W. Simian herpesviruses and their risk to humans. Vaccine 2010, 28, B78–B84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huff, J.L.; Barry, P.A. B-Virus (Cercopithecine herpesvirus 1) infection in humans and macaques: Potential for zoonotic disease. Emerg. Infect. Dis. 2003, 9, 246–250. [Google Scholar] [CrossRef]
- Vindevogel, H.; Duchatel, J.P. Miscellaneous herpesvirus infections. In Diseases of Poultry, 10th ed.; Calnek, B.W., Barnes, J.H., Beard, C.W., McDougald, L.R., Saif, Y.M., Eds.; Iowa State University Press: Ames, IA, USA, 1997; pp. 757–761. [Google Scholar]
- Ehlers, B.; Küchler, J.; Yasmum, N.; Dural, G.; Voigt, S.; Schmidt-Chanasit, J.; Jäkel, T.; Matuschka, F.R.; Richter, D.; Essbauer, S.; et al. Identification of novel rodent herpesviruses, including the first gammaherpesvirus of Mus musculus. J. Virol. 2007, 81, 8091–8100. [Google Scholar] [CrossRef] [Green Version]
- Davison, A.J.; Clements, J.B. Herpesviruses: General properties. In Virology, 10th ed.; Mahy, B.W.J., Meulen, V., Eds.; Edward Arnold Ltd.: London, UK, 2005; pp. 488–505. [Google Scholar]


{kind=link}
| Tawny Owl | Ural Owl | |
|---|---|---|
| Infected birds | 0.0% (115) | 29.1% (55) |
| Infected adults | 0.0% (27) | 18.7% (16) |
| Infected young | 0.0% (88) | 33.3% (39) |
| Infected nests | 0.0% (30) | 53.0% (17) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Žlabravec, Z.; Vrezec, A.; Slavec, B.; Kuhar, U.; Zorman Rojs, O.; Račnik, J. Herpesvirus Infection in a Breeding Population of Two Coexisting Strix Owls. Animals 2021, 11, 2519. https://doi.org/10.3390/ani11092519
Žlabravec Z, Vrezec A, Slavec B, Kuhar U, Zorman Rojs O, Račnik J. Herpesvirus Infection in a Breeding Population of Two Coexisting Strix Owls. Animals. 2021; 11(9):2519. https://doi.org/10.3390/ani11092519
Chicago/Turabian StyleŽlabravec, Zoran, Al Vrezec, Brigita Slavec, Urška Kuhar, Olga Zorman Rojs, and Joško Račnik. 2021. "Herpesvirus Infection in a Breeding Population of Two Coexisting Strix Owls" Animals 11, no. 9: 2519. https://doi.org/10.3390/ani11092519
APA StyleŽlabravec, Z., Vrezec, A., Slavec, B., Kuhar, U., Zorman Rojs, O., & Račnik, J. (2021). Herpesvirus Infection in a Breeding Population of Two Coexisting Strix Owls. Animals, 11(9), 2519. https://doi.org/10.3390/ani11092519