
Dietary Lactobacillus casei K17 Improves Lipid Metabolism, Antioxidant Response, and Fillet Quality of Micropterus salmoides

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diet and Culture Conditions of M. salmoides
2.2. Sample Collection
2.3. Determination of Fillet Water, Crude Protein, Crude Fat, Crude Ash, and Collagen of Fish Samples
2.4. Quality Analysis of Fish Fillets
2.5. Liver and Fillet Histology
2.6. Determination of Antioxidant and Lipid Indexes in Liver and Blood
2.7. Statistical Analysis
3. Results
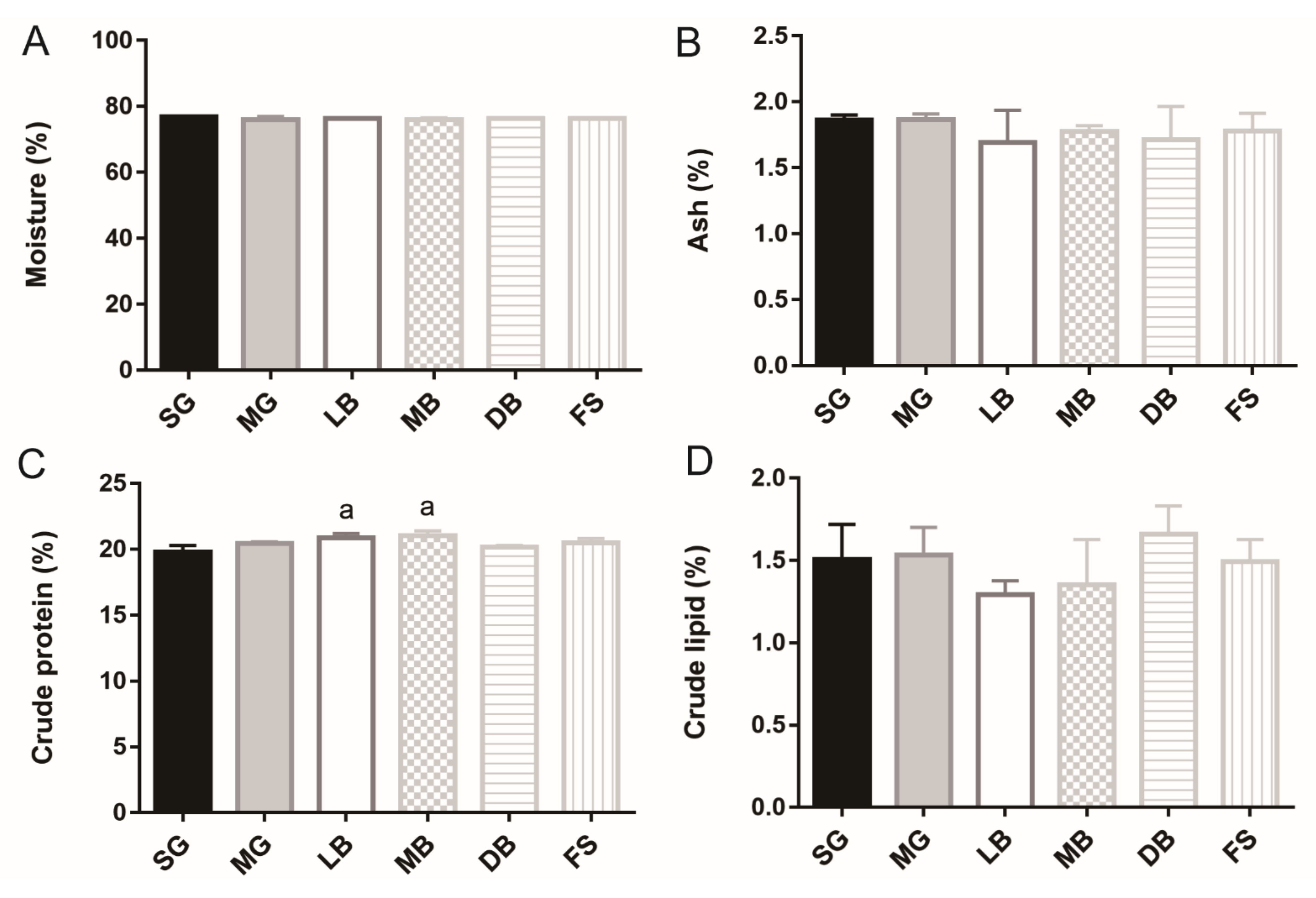
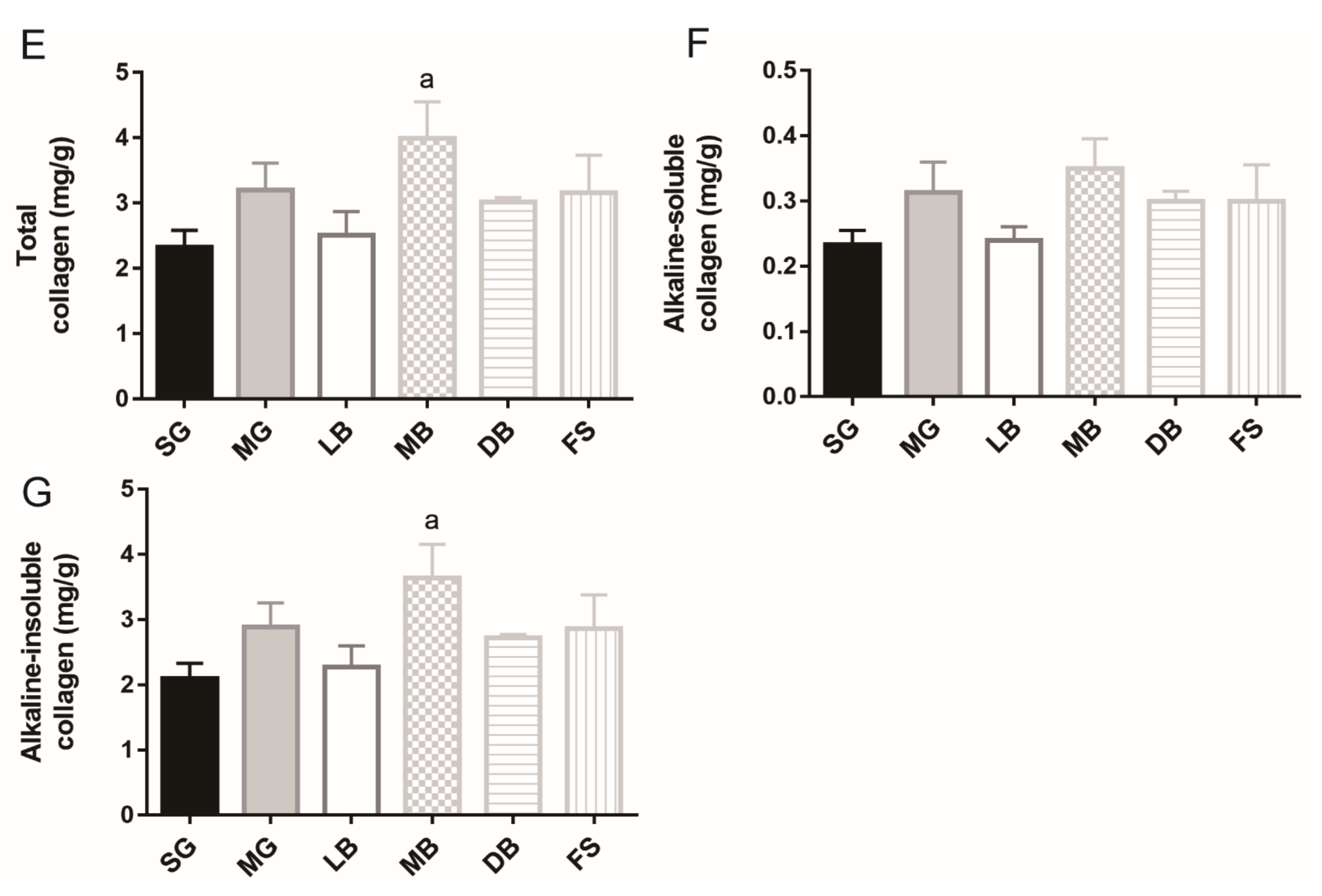
3.1. Fillet Moisture, Crude Protein, Crude Fat, Crude Ash, and Collagen of M. salmoides
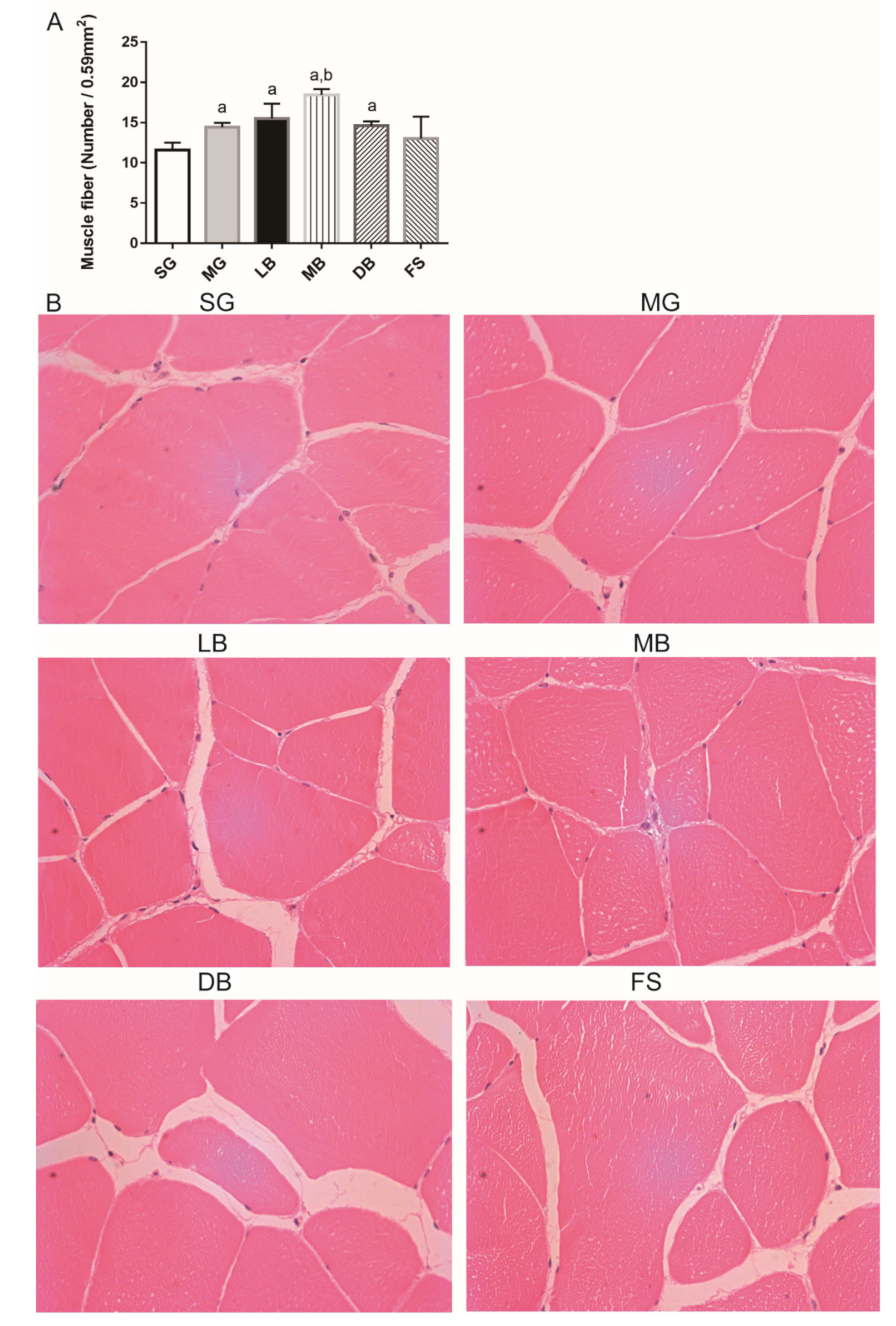
3.2. Fillet Histology of M. salmoides
3.3. Fillet Texture
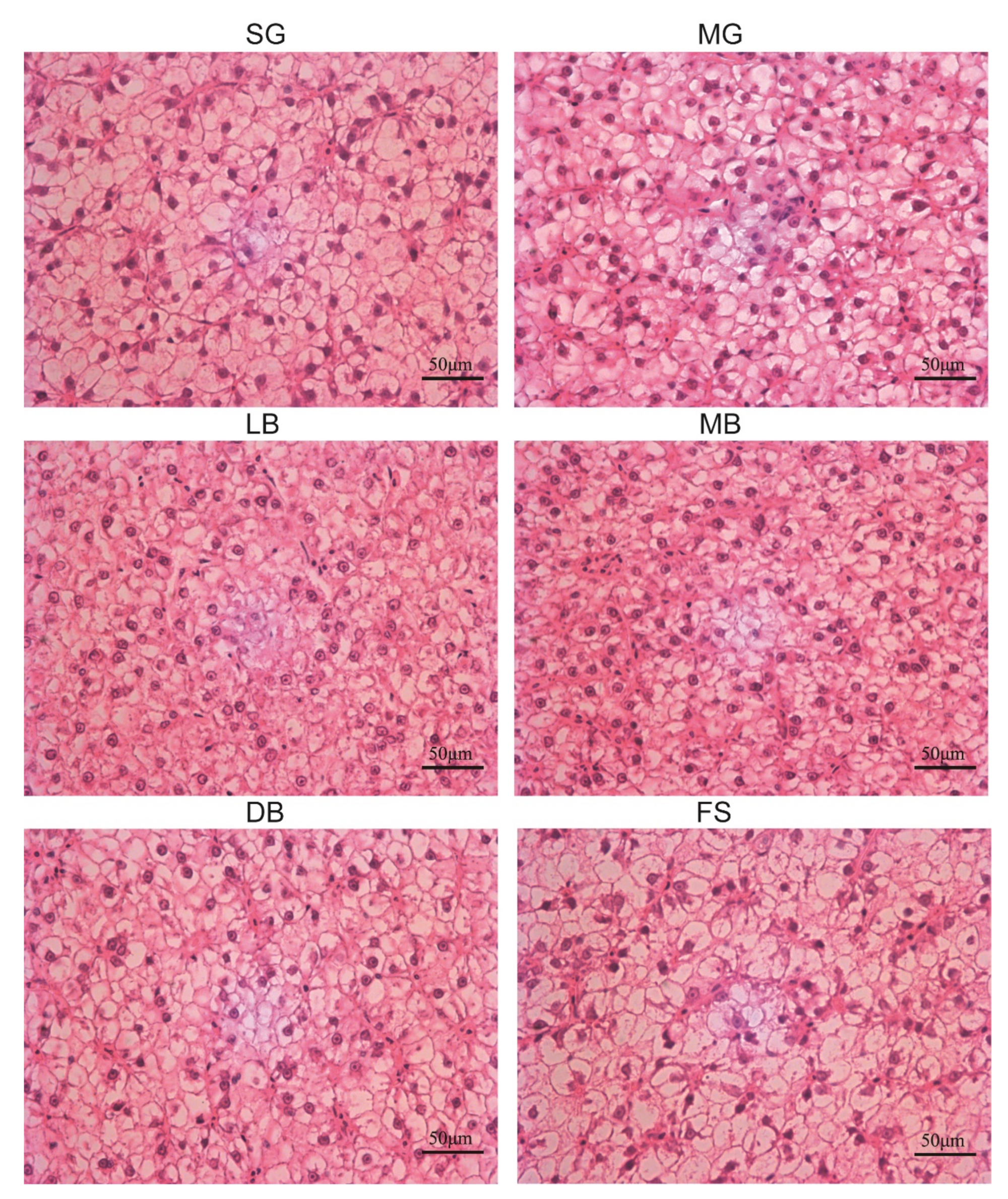
3.4. Liver Histology
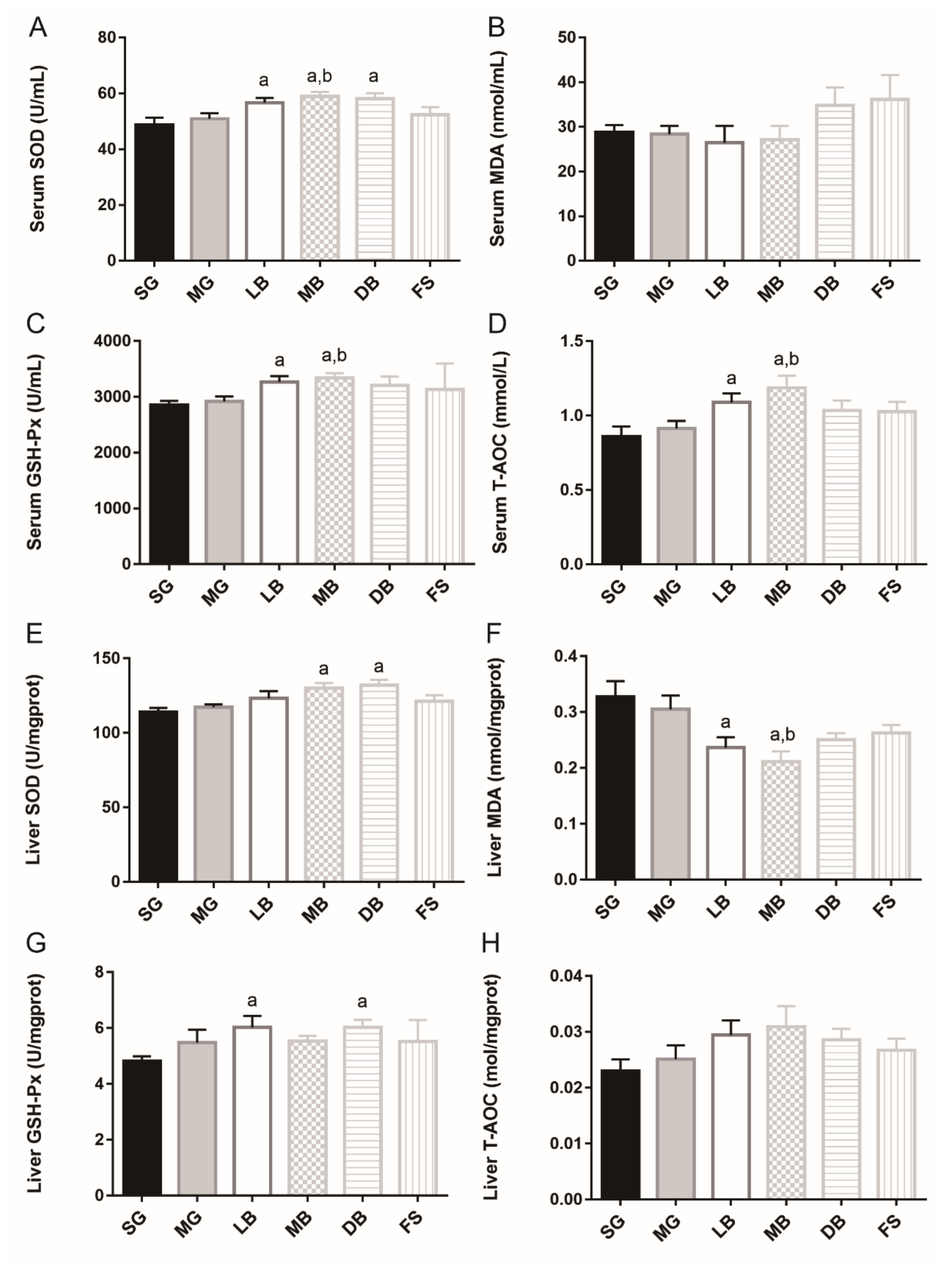
3.5. Antioxidant Response
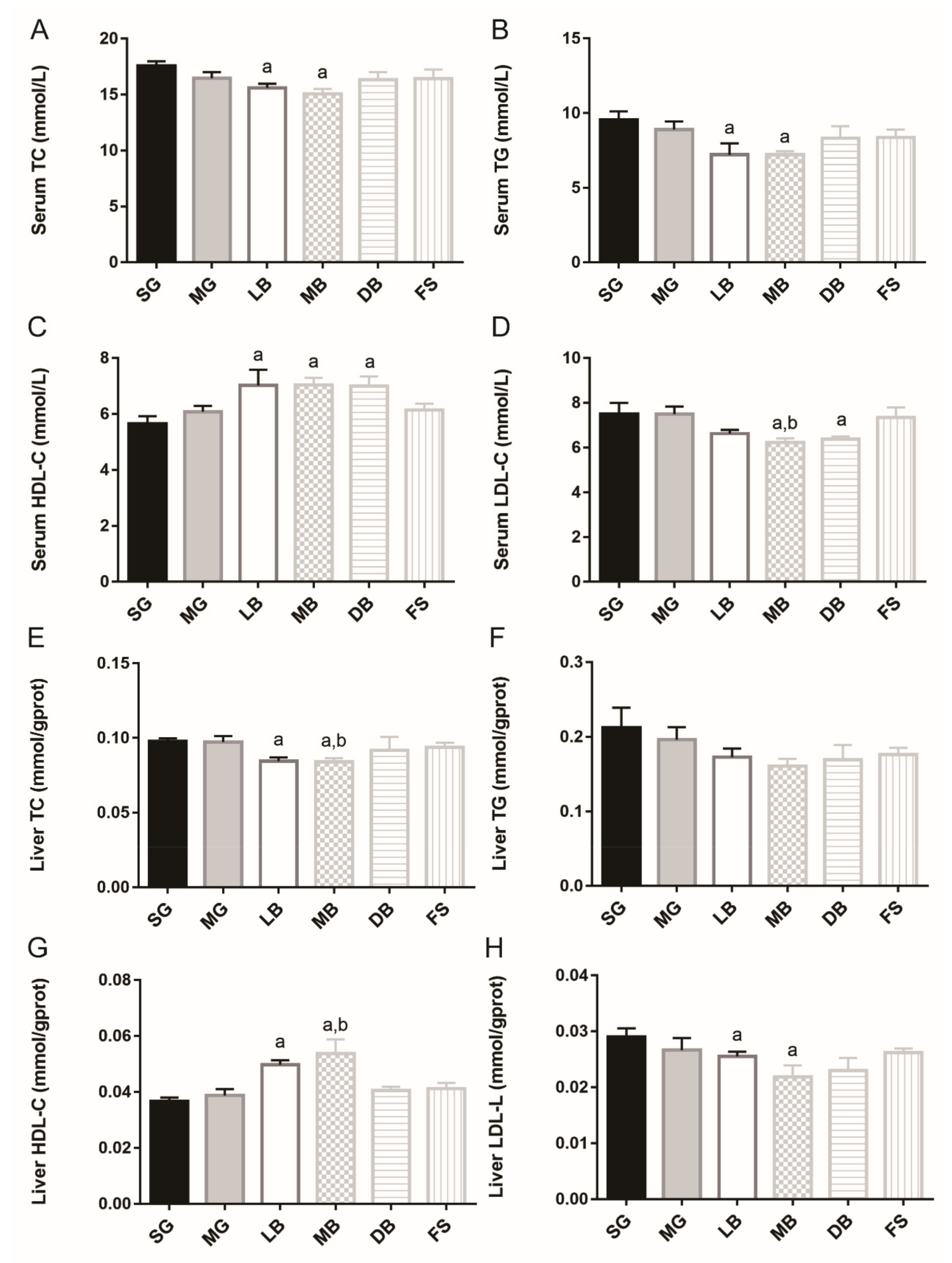
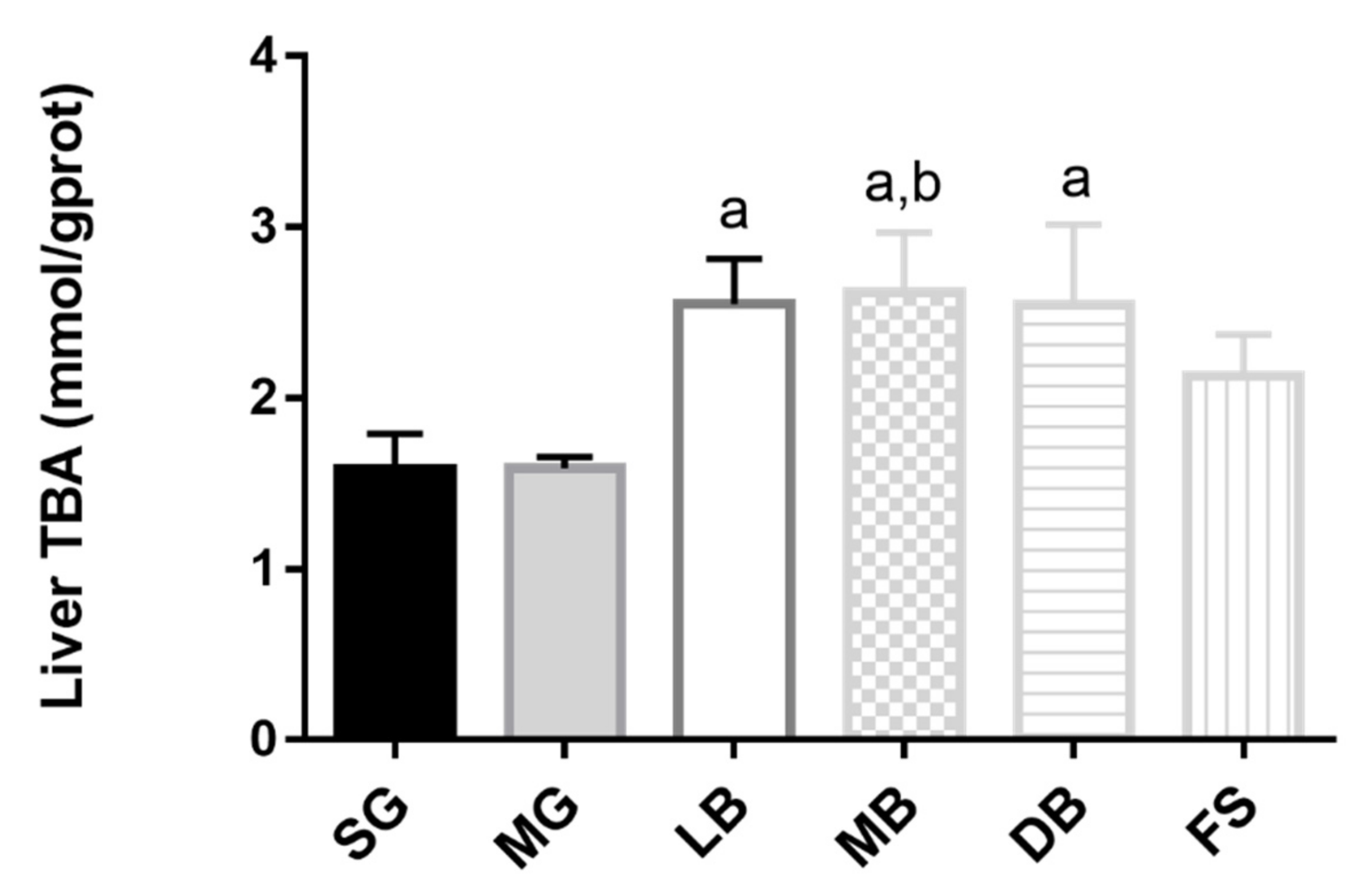
3.6. Lipid Metabolism of M. salmoides
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Coyle, S.D.; Tidwell, J.H.; Webster, C.D. Response of Largemouth Bass Micropterus salmoides to dietary supplementation of lysine, methionine, and highly unsaturated fatty acids. J. World Aquacult. Soc. 2007, 31, 89–95. [Google Scholar] [CrossRef]
- Gong, Y.; Yang, F.; Hu, J.; Liu, C.; Liu, H.; Han, D.; Jin, J.; Yang, Y.; Zhu, X.; Yi, J.; et al. Effects of dietary yeast hydrolysate on the growth, antioxidant response, immune response and disease resistance of largemouth bass (Micropterus salmoides). Fish Shellfish. Immunol. 2019, 94, 548–557. [Google Scholar] [CrossRef]
- Zhang, T.; Huang, P.D.; Zhao, Y.F. Thinking of ulva, Jiangsu taste how to continue? Where is the high-quality industry of Micropterus salmoides? Sci. Fish Farming 2019, 12, 20–23. [Google Scholar]
- Wang, X.; Hu, J.; Wang, Y.; Zhou, J. Research progress of nutritional requirement for largemouth bass (Micropterus salmoides). Feed. Res. 2019, 8, 212–221. [Google Scholar]
- Chen, Y.; Liu, Y.; Yang, H.; Yuan, Y.; Liu, F.; Tian, L.; Liang, G.; Yuan, R. Effect of dietary oxidized fish oil on growth performance, body composition, antioxidant defence mechanism and liver histology of juvenile largemouth bass Micropterus salmoides. Aquacult. Nutr. 2012, 18, 321–331. [Google Scholar] [CrossRef]
- Yun, B.; Xue, M.; Wang, J.; Fan, Z.; Wu, X.; Zheng, Y.; Qin, Y. Effects of lipid sources and lipid peroxidation on feed intake, growth, and tissue fatty acid compositions of largemouth bass (Micropterus salmoides). Aquacult. Int. 2013, 21, 97–110. [Google Scholar] [CrossRef]
- Rathore, S.S.; Murthy, H.S.; Abdullah-Al Mamun, M.; Nasren, S.; Rakesh, K.; Kumar, B.T.N.; Abhiman, P.B.; Khandagale, A.S. Nano-selenium supplementation to ameliorate nutrition physiology, immune response, antioxidant system and disease resistance against Aeromonas hydrophilain Monosex Nile Tilapia (Oreochromis niloticus). Biol. Trace. Elem. 2020, 6, 256–266. [Google Scholar] [CrossRef]
- Xu, Y.; Zheng, Y.; Li, W.; Liu, Y.; Ding, Z. Gene expression, antioxidation and growth were considerably promoted by feeding dietary vitamin E and alanyl-glutamine dipeptide supplementation in juvenile tilapia in cold freshwater. Aquacult. Nutr. 2020, 26, 2159–2168. [Google Scholar] [CrossRef]
- Fontagné-Dicharry, S.; Véron, V.; Larroquet, L.; Godin, S.; Kaushik, S.J. Effect of selenium sources in plant-based diets on antioxidant status and oxidative stress-related parameters in rainbow trout juveniles under chronic stress exposure. Aquaculture 2020, 529, 735684. [Google Scholar] [CrossRef]
- Dawood, M.A.O.; Moustafa, E.M.; Elbialy, Z.I.; Farrag, F.; Lolo, E.E.E.; Abdel-Daim, H.A.; Abdel-Daim, M.M.; Van Doan, H. Lactobacillus plantarum L-137 and/or beta-glucan impacted the histopathological, antioxidant, immune-related genes and resistance of Nile tilapia (Oreochromis niloticus) against Aeromonas hydrophila. Res. Vet. Sci. 2020, 130, 212–221. [Google Scholar] [CrossRef]
- Chen, X.; Zhang, Z.; Fernandes, J.; Gao, Y.; Yin, P.; Liu, Y.; Tian, L.; Xie, S.; Niu, J. Beneficial effects on growth, haematic indicators, immune status, antioxidant function and gut health in Juvenile Nile tilapia (Oreochromis niloticus) by dietary administration of a multi-strain probiotic. Aquac. Nutr. 2020, 26, 1369–1382. [Google Scholar] [CrossRef]
- Anonymous. 2045 notice of the ministry of agriculture of the People’s Republic of China. China Feed 2014, 7, 39–42. [Google Scholar]
- Zhao, Y.; Li, J.; Yin, L.; Feng, L.; Liu, Y.; Jiang, W.; Wu, P.; Zhao, J.; Chen, D.; Zhou, X.; et al. Effects of dietary glutamate supplementation on flesh quality, antioxidant defense and gene expression related to lipid metabolism and myogenic regulation in Jian carp (Cyprinus carpio var. Jian). Aquaculture 2019, 502, 212–222. [Google Scholar] [CrossRef]
- Yang, G.; Cao, H.; Jiang, W.; Hu, B.; Jian, S.; Wen, C.; Kajbaf, K.; Kumar, V.; Tao, Z.; Peng, M. Dietary supplementation of Bacillus cereus as probiotics in Pengze crucian carp (Carassius auratus var. Pengze): Effects on growth performance, fillet quality, serum biochemical parameters and intestinal histology. Aquac. Res. 2019, 50, 2207–2217. [Google Scholar] [CrossRef]
- Cao, H.; Yu, R.; Zhang, Y.; Hu, B.; Jian, S.; Wen, C.; Kajbaf, K.; Kumar, V.; Yang, G. Effects of dietary supplementation with β-glucan and Bacillus subtilis on growth, fillet quality, immune capacity, and antioxidant status of Pengze crucian carp (Carassius auratus var. Pengze). Aquaculture 2019, 508, 106–112. [Google Scholar] [CrossRef]
- Xu, S.; Liu, T.; Radji, C.A.; Yang, J.; Chen, L. Isolation, identification, and evaluation of new lactic acid bacteria strains with both cellular antioxidant and bile salt hydrolase activities in Vitro. J. Food Prot. 2016, 79, 1919–1928. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, L. Impact of a novel nano-protectant on the viability of probiotic Bacterium Lactobacillus casei K17. Foods 2021, 10, 529. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Wang, H.; Hu, K.; Jiao, L.; Xia, L. Effect of Dietary Supplementation of Lactobacillus Casei YYL3 and L. Plantarum YYL5 on Growth, Immune Response and Intestinal Microbiota in Channel Catfish. Animals 2019, 9, 1005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhu, Z.; Li, R.; Wang, X.; Leng, X.; Chen, L. Impact of supplementary Lactobacillus casei K17 on growth and gut health of largemouth bass Micropterus salmoides. Aquac. Rep. 2021, 20, 100734. [Google Scholar] [CrossRef]
- Soltani, M.; Abdy, E.; Alishahi, M.; Mirghaed, A.T.; Hosseini-Shekarabi, P. Growth performance, immune-physiological variables and disease resistance of common carp (Cyprinus carpio) orally subjected to different concentrations of Lactobacillus plantarum. Aquacult. Int. 2017, 25, 1913–1933. [Google Scholar] [CrossRef]
- Miller, E.L.; Bimbo, A.P.; Barlow, S.M.; Sheridan, B. Repeatability and reproducibility of determination of the nitrogen content of fishmeal by the combustion (Dumas) method and comparison with the Kjeldahl method: Interlaboratory study. J. AOAC Int. 2007, 90, 6–20. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Ai, Q.; Mai, K.; Xu, W.; Liufu, Z.; Zhang, Y.; Peng, M. Effects of dietary hydroxyproline on growth performance, body composition, hydroxyproline and collagen concentrations in tissues in relation to prolyl 4-hydroxylase α(I) gene expression of juvenile turbot, Scophthalmus maximus L. fed high plant protein diets. Aquaculture 2013, 404, 77–84. [Google Scholar]
- Torrecillas, S.; Makol, A.; Caballero, M.J.; Montero, D.; Robaina, L.; Real, F.; Sweetman, J.; Tort, L.; Izquierdo, M.S. Immune stimulation and improved infection resistance in European sea bass (Dicentrarchus labrax) fed mannan oligosaccharides. Fish Shellfish. Immunol. 2007, 23, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Chen, N.; Liu, Z.; Gou, S.; Yin, J. Effects of dietary starch sources and levels on liver histology in largemouth bass, Micropterus salmoides. J. Shanghai Ocean. Univ. 2016, 25, 64–73. [Google Scholar]
- Yao, W.; Li, X.; Kabir Chowdhury, M.A.; Wang, J.; Leng, X. Dietary protease, carbohydrase and micro-encapsulated organic acid salts individually or in-combination improved growth, feed utilization and intestinal histology of Pacific white shrimp. Aquaculture 2018, 503, 88–95. [Google Scholar] [CrossRef]
- Tan, X.; Lin, H.; Huang, Z.; Zhou, C.; Wang, A.; Qi, C.; Zhao, S. Effects of dietary leucine on growth performance, feed utilization, non-specific immune responses and gut morphology of juvenile golden pompano Trachinotus ovatus. Aquaculture 2016, 465, 100–107. [Google Scholar] [CrossRef]
- Tan, X.; Sun, Z.; Liu, Q.; Ye, H.; Zou, C.; Ye, C.; Wang, A.; Lin, H. Effects of dietary ginkgo biloba leaf extract on growth performance, plasma biochemical parameters, fish composition, immune responses, liver histology, and immune and apoptosis-related genes expression of hybrid grouper (Epinephelus lanceolatus♂ × Epinephelus fuscoguttatus♀) fed high lipid diets. Fish Shellfish. Immunol. 2018, 72, 399–409. [Google Scholar] [CrossRef]
- Kumar, P.; Jain, K.K.; Sardar, P.; Jayant, M.; Tok, N.C. Effect of dietary synbiotic on growth performance, body composition, digestive enzyme activity and gut microbiota in Cirrhinus mrigala (Ham.) fingerlings. Aquacult. Nutr. 2018, 24, 921–929. [Google Scholar] [CrossRef]
- Ye, J.; Wang, K.; Li, F.; Sun, Y. Single or combined effects of fructo- and mannan oligosaccharide supplements and Bacillus clausii on the growth, feed utilization, body composition, digestive enzyme activity, innate immune response and lipid metabolism of the Japanese flounder Paralichthys olivaceus. Aquacult. Nutr. 2011, 17, 902–911. [Google Scholar] [CrossRef]
- Adorian, T.J.; Jamali, H.; Farsani, H.G.; Darvishi, P.; Hasanpour, S.; Bagheri, T.; Roozbehfar, R. Effects of probiotic bacteria Bacillus on growth performance, digestive enzyme activity, and hematological parameters of Asian Sea Bass, Lates calcarifer (Bloch). Probiotics Antimicrob. Proteins 2019, 11, 248–255. [Google Scholar] [CrossRef]
- Hooshyar, Y.; Abedian Kenari, A.; Paknejad, H.; Gandomi, H. Effects of Lactobacillus rhamnosus ATCC 7469 on different parameters related to health status of Rainbow Trout (Oncorhynchus mykiss) and the protection against Yersinia ruckeri. Probiotics Antimicrob. Proteins 2020, 12, 1370–1384. [Google Scholar] [CrossRef]
- Allameh, S.K.; Yusoff, F.M.; Ringø, E.; Daud, H.M.; Saad, C.R. Effects of dietary mono- and multiprobiotic strains on growth performance, gut bacteria and body composition of Javanese carp (Puntius gonionotus, Bleeker 1850). Aquacult. Nutr. 2016, 22, 367–373. [Google Scholar] [CrossRef]
- Rodrigez, M.A.S.D.; Daz-Rosales, P.; Chabrilln, M.; Smidt, H.; Moyano, F.J. Effect of dietary administration of probiotics on growth and intestine functionality of juvenile Senegalese sole (Solea senegalensis, Kaup 1858). Aquacult. Nutr. 2010, 15, 177–185. [Google Scholar]
- Gordon, M.K.; Hahn, R.A. Collagens. Cell Tissue Res. 2010, 339, 247–257. [Google Scholar] [CrossRef] [PubMed]
- Hatae, K.; Tobimatsu, A.; Takeyama, M.; Matsumoto, J.J. Contribution of the connective tissues on the texture difference of various fish species. Nippon Suisan Gakkaishi 1986, 52, 2001–2007. [Google Scholar] [CrossRef]
- Johnston, I.A.; Li, X.; Vieira, V.L.A.; Nickell, D.; Dingwall, A.; Alderson, R.; Campbell, P.; Bickerdike, R. Muscle and flesh quality traits in wild and farmed Atlantic salmon. Aquaculture 2006, 256, 323–336. [Google Scholar] [CrossRef]
- Li, X.; Bickerdike, R.; Lindsay, E.; Campbell, P.; Nickell, D.; Dingwall, A.; Johnston, I.A. Hydroxylysyl pyridinoline cross-link concentration affects the textural properties of fresh and smoked Atlantic salmon (Salmo salar L.) flesh. J. Agric. Food Chem. 2005, 53, 6844–6850. [Google Scholar] [CrossRef]
- Sun, W.; Xu, X.; Li, X.; Pan, W.; Leng, X. Effects of dietary geniposidic acid on growth performance, flesh quality and collagen gene expression of grass carp, Ctenopharyngodon idella. Aquacult. Nutr. 2018, 24, 1112–1121. [Google Scholar] [CrossRef]
- Rehfeldt, C.; Pas, T.M.F.W.; Wimmers, K.; Brameld, J.M.; Nissen, P.M. Advances in research on the prenatal development of skeletal muscle in animals in relation to the quality of muscle-based food. I. Regulation of myogenesis and environmental impact (review). Animal 2011, 5, 703–717. [Google Scholar] [CrossRef] [Green Version]
- Bayol, S.; Jones, D.; Goldspink, G.; Stickland, N.C. The influence of undernutrition during gestation on skeletal muscle cellularity and on the expression of genes that control muscle growth. Brit. J. Nutr. 2004, 91, 331–339. [Google Scholar] [CrossRef]
- Monteschio, J.O.; Vargas-Junior, F.M.; Almeida, F.L.A.; Pinto, L.A.M.; Kaneko, I.N.; Almeida, A.A.; Freitas, L.W.; Alves, S.P.A.; Bessa, R.J.B.; Prado, I.N. The effect of encapsulated active principles (eugenol, thymol and vanillin) and clove and rosemary essential oils on the structure, collagen content, chemical composition and fatty acid profile of Nellore heifers muscle. Meat Sci. 2019, 155, 27–35. [Google Scholar] [CrossRef]
- Weatherley, A.H.; Gill, H.S.; Lobo, A.F. Recruitment and maximal diameter of axial muscle fibres in teleosts and their relationship to somatic growth and ultimate size. J. Fish Biol. 2010, 33, 851–859. [Google Scholar] [CrossRef]
- Johnston, I.A.; Alderson, R.; Sandham, C.; Dingwall, A.; Mitchell, D.; Selkirk, C.; Nickell, D.; Baker, R.; Robertson, B.; Whyte, D.; et al. Muscle fibre density in relation to the colour and texture of smoked Atlantic salmon ( Salmo salar L.). Aquaculture 2000, 189, 335–349. [Google Scholar] [CrossRef]
- Modzelewska-Kapituła, M.; Tkacz, K.; Nogalski, Z.; Karpińska-Tymoszczyk, M.; Draszanowska, A.; Pietrzak-Fiećko, R.; Purwin, C.; Lipiński, K. Addition of herbal extracts to the Holstein-Friesian bulls’ diet changes the quality of beef. Meat Sci. 2018, 145, 163–170. [Google Scholar] [CrossRef]
- Falowo, A.B.; Fayemi, P.O.; Muchenje, V. Natural antioxidants against lipid-protein oxidative deterioration in meat and meat products: A review. Food Res. Int. 2014, 64, 171–181. [Google Scholar] [CrossRef]
- Aberle, E.D.; Reeves, E.S.; Judge, M.D.; Hunsley, R.E.; Perry, T.W. Palatability and muscle characteristics of cattle with controlled weight gain: Time on a high energy diet. J. Anim. 1981, 52, 757–763. [Google Scholar] [CrossRef]
- Cheng, C.; Yang, F.; Ling, R.; Liao, S.; Miao, Y.; Ye, C.; Wang, A. Effects of ammonia exposure on apoptosis, oxidative stress and immune response in pufferfish (Takifugu obscurus). Aquat. Toxicol. 2015, 164, 61–71. [Google Scholar] [CrossRef] [PubMed]
- Dawood, M.A.O.; Magouz, F.I.; Salem, M.F.I.; Abdel-Daim, H.A. Modulation of digestive enzyme activity, blood health, oxidative responses and growth-related gene expression in GIFT by heat-killed Lactobacillus plantarum (L-137). Aquaculture 2019, 505, 127–136. [Google Scholar] [CrossRef]
- Zhang, C.; Zhang, J.; Guan, W.; Zhang, X.; Guan, S.; Zeng, Q.; Cheng, G.; Cui, W. Effects of Lactobacillus delbrueckii on immune response, disease resistance against Aeromonas hydrophila, antioxidant capability and growth performance of Cyprinus carpio Huanghe var. Fish Shellfish. Immunol. 2017, 68, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Farideh, E.H.N.; Forouzan, B.; Lakani, B. Effects of biofloc on growth performance, digestive enzyme activities and liver histology of common carp (Cyprinus carpio L.) fingerlings in zero-water exchange system. Fish Physiol. Biochem. 2016, 42, 457–465. [Google Scholar]
- Strüssmann, C.; Takashima, F. Hepatocyte nuclear size and nutritional condition of larval pejerrey, Odontesthes bonariensis (Cuvier et Valenciennes). J. Fish Biol. 2010, 36, 59–65. [Google Scholar] [CrossRef]
- Zhao, D.; Wu, T.; Yi, D.; Wang, L.; Li, P.; Zhang, J.; Hou, Y.; Wu, G. Dietary supplementation with Lactobacillus casei alleviates lipopolysaccharide-induced liver injury in a porcine model. Int. J. Mol. Sci. 2017, 18, 2535. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Du, R.; He, Q.; Li, H. Effect of feeding probiotic Lactobacillus casei Zhang on liver lipid metabolism in rats with high-fat diet. Sci. Agric. Sin. 2012, 45, 943–950. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment Group | Feed |
|---|---|
| Saline group (SG) | 300 g basic feed + 30 mL 0.85% sterile saline |
| Milk group (MG) | 300 g basic feed + 30 mL 10% skim milk powder |
| Live bacteria (LB) | 300 g basic feed + 30 mL live L. casei K17 108 CFU/mL |
| Live bacteria protected by skim milk powder (MB) | 300 g basic feed+ 30 mL (10% skim milk powder + live L. casei K17 108 CFU/mL) |
| Dead bacteria (DB) | 300 g basic feed + 30 mL dead L. casei K17 108 CFU/mL |
| Fermentation supernatant (FS) | 300 g basic feed + 30 mL fermentation supernatant |
| Group | SG | MG | LB | MB | DB | FS | |
|---|---|---|---|---|---|---|---|
| Hardness (g) | Unadjusted | 490.67 ± 147.03 | 524.67 ± 96.55 | 729.33 ± 119.49 | 945.33 ± 264.13 b | 590.00 ± 95.14 | 718.00 ± 151.25 |
| Adjusted | 456.05 a ± 109.37 | 499.00 a ± 102.27 | 739.66 a ± 94.44 | 972.02 a ± 102.99 | 622.51 a ± 107.55 | 708.76 a ± 94.13 | |
| Stickiness (g.s) | Unadjusted | 5.81 ± 0.81 | 7.37 ± 1.70 | 7.81 ± 2.45 | 9.22 ± 2.36 | 7.46 ± 3.09 | 7.97 ± 3.55 |
| Adjusted | 6.11 a ± 1.76 | 7.59 a ± 1.65 | 7.72 a ± 1.52 | 8.99 a ± 1.66 | 7.18 a ± 1.73 | 8.05 a ± 1.52 | |
| Springiness (mm) | Unadjusted | 0.517 ± 0.068 | 0.457 ± 0.012 | 0.437 ± 0.061 | 0.483 ± 0.058 | 0.437 ± 0.061 | 0.487 ± 0.035 |
| Adjusted | 0.541 a ± 0.035 | 0.475 a ± 0.033 | 0.429 a ± 0.030 | 0.464 a ± 0.033 | 0.413 a ± 0.034 | 0.493 a ± 0.030 | |
| Chewiness (mJ) | Unadjusted | 109.96 ± 33.80 | 120.74 ± 21.92 | 207.34 ± 11.04 | 252.05 ± 40.90 b | 184.54 ± 30.05 | 170.69 ± 24.92 |
| Adjusted | 112.01 a ± 20.35 | 122.26 a ± 19.03 | 206.73 a ± 17.57 | 250.48 a ± 19.16 b | 182.62 a ± 20.01 | 171.24 a ± 17.51 | |
| Gumminess (g) | Unadjusted | 315.12 ± 126.06 | 345.54 ± 90.57 | 489.50 ± 82.82 | 609.30 ± 128.59 b | 374.02 ± 55.57 | 442.65 ± 109.35 |
| Adjusted | 304.47 a ± 72.26 | 337.64 a ± 67.56 | 492.676 a ± 62.39 | 617.51 a ± 68.04 | 384.02 a ± 71.06 | 439.81 a ± 62.19 | |
| Cohesiveness | Unadjusted | 0.65 ± 0.05 | 0.64 ± 0.06 | 0.65 ± 0.06 | 0.61 ± 0.03 | 0.63 ± 0.07 | 0.67 ± 0.03 |
| Adjusted | 0.68 a ± 0.03 | 0.67 a ± 0.03 | 0.64 a ± 0.03 | 0.59 a ± 0.03 | 0.60 a ± 0.03 | 0.68 a ± 0.03 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Zhu, Z.; Tian, S.; Fu, H.; Leng, X.; Chen, L. Dietary Lactobacillus casei K17 Improves Lipid Metabolism, Antioxidant Response, and Fillet Quality of Micropterus salmoides. Animals 2021, 11, 2564. https://doi.org/10.3390/ani11092564
Wang J, Zhu Z, Tian S, Fu H, Leng X, Chen L. Dietary Lactobacillus casei K17 Improves Lipid Metabolism, Antioxidant Response, and Fillet Quality of Micropterus salmoides. Animals. 2021; 11(9):2564. https://doi.org/10.3390/ani11092564
Chicago/Turabian StyleWang, Jinsong, Zhuoying Zhu, Shenghao Tian, Huiyu Fu, Xiangjun Leng, and Lanming Chen. 2021. "Dietary Lactobacillus casei K17 Improves Lipid Metabolism, Antioxidant Response, and Fillet Quality of Micropterus salmoides" Animals 11, no. 9: 2564. https://doi.org/10.3390/ani11092564
APA StyleWang, J., Zhu, Z., Tian, S., Fu, H., Leng, X., & Chen, L. (2021). Dietary Lactobacillus casei K17 Improves Lipid Metabolism, Antioxidant Response, and Fillet Quality of Micropterus salmoides. Animals, 11(9), 2564. https://doi.org/10.3390/ani11092564