
Attempted Control of Paratuberculosis in Dairy Calves by Only Changing the Quality of Milk Fed to Calves

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Design
2.2. Study Population
2.2.1. Calves
2.2.2. Nurse Cows
2.3. Milk Treatments
2.3.1. Bacteriological Analyses of Milk and Environmental Samples
2.3.2. Estimation of MAP Load in Milk Samples
2.3.3. Evaluation of the Infection Progression in the Study Animals
Infectious Status Determination
Post-Mortem Analyses
2.3.4. Ancillary Data: Monitoring Potential Copper Toxicosis in Calves
2.3.5. Statistical Analysis
3. Results
3.1. Descriptive
3.2. Detection of MAP in Milk and Environmental Samples
3.3. Detection of MAP Infection in Calves
3.4. Analytical
3.5. Ancillary Data: Monitoring Copper Toxicosis in Calves Fed Copper-Treated Milk
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| AST | Aspartate aminotransferase |
| ATCC | American Type Culture Collection |
| Bce | Bacterial cell equivalent |
| ELISA | Enzyme-linked immunoabsorbent assay |
| GD | Glutamate dehydrogenase |
| GGT | Gamma-glutamyl transferase |
| MAP | Mycobacterium avium subsp. paratuberculosis |
| MGIT | Mycobacterial growth indicator tube |
| PANTA | Polymyxin B, amphotericin B, nalidixic acid, trimethoprim, and azlocillin |
| PBS | Phosphate buffered saline |
| qPCR | Quantitative polymerase chain reaction |
| UHT | Ultra-high temperature |
| ZIPMM | Zero inflated Poisson mixed model |
References
- Harris, N.B.; Barletta, R.G. Mycobacterium avium subsp. paratuberculosis in Veterinary Medicine. Clin. Microbiol. Rev. 2001, 14, 489–512. [Google Scholar] [CrossRef] [Green Version]
- Windsor, P.A.; Whittington, R.J. Evidence for age susceptibility of cattle to Johne’s disease. Vet. J. 2010, 184, 37–44. [Google Scholar] [CrossRef]
- Sweeney, R.W. Pathogenesis of Paratuberculosis. Vet. Clin. Food Anim. 2011, 27, 537–546. [Google Scholar] [CrossRef]
- Van Metre, D.C.; Tennant, B.C.; Whitlock, R.H. Infectious diseases of the gastrointestinal tract. In Rebhun’s Diseases of Dairy Cattle; Divers, T.J., Peek, S.F., Eds.; Elsevier: St-Louis, MI, USA, 2008; pp. 200–294. [Google Scholar]
- Fecteau, M.; Whitlock, R. Paratuberculosis in cattle. In Paratuberculosis: Organism, Disease, Control; Behr, M., Collins, D.M., Eds.; CABI: Wallingford, UK, 2010; pp. 144–153. [Google Scholar]
- Manning, E.; Collins, M. Epidemiology of Paratuberculosis. In Paratuberculosis: Organism, Disease, Control; Behr, M., Collins, D.M., Eds.; CABI: Wallingford, UK, 2010; pp. 22–26. [Google Scholar]
- Lombard, J.E. Epidemiology and Economics of Paratuberculosis. Vet. Clin. Food Anim. 2011, 27, 525–535. [Google Scholar] [CrossRef]
- Whittington, R.; Donat, K.; Weber, M.F.; Kelton, D.; Nielsen, S.S.; Eisenberg, S.; Arrigoni, N.; Juste, R.; Sáez, J.L.; Dhand, N.; et al. Control of paratuberculosis: Who, why and how. A review of 48 countries. BMC Vet. Res. 2019, 15, 198. [Google Scholar] [CrossRef] [Green Version]
- Stehman, S.M. Paratuberculosis in small ruminants, deer, and South American camelids. Vet. Clin. Food Anim. 1996, 12, 441–455. [Google Scholar] [CrossRef]
- Clarke, C.J. The pathology and pathogenesis of paratuberculosis in ruminants and other species. J. Comp. Pathol. 1997, 116, 217–261. [Google Scholar] [CrossRef]
- Sweeney, R.W. Transmission of paratuberculosis. Vet. Clin. Food Anim. 1996, 12, 305–312. [Google Scholar] [CrossRef]
- Taylor, T.K.; Wilks, C.R.; McQueen, D.S. Isolation of Mycobacterium paratuberculosis from the milk of a cow with Johne’s disease. Vet. Rec. 1981, 109, 532–533. [Google Scholar] [CrossRef]
- Streeter, R.N.; Hoffsis, G.F.; Bech-Nielsen, S.; Shulaw, W.P.; Ring, M. Isolation of Mycobacterium paratuberculosis from colostrum and milk of subclinically infected cows. Am. J. Vet. Res. 1995, 56, 1322–1324. [Google Scholar]
- Grant, I.R.; Foddai, A.; Tarrant, J.; Kunkel, B.; Hartmann, F.A.; McGuirk, S.; Hansen, C.; Talaat, A.M.; Collins, M.T. Viable Mycobacterium avium ssp. paratuberculosis isolated from calf milk replacer. J. Dairy Sci. 2017, 100, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Grant, I.R.; Hitchings, E.I.; McCartney, A.; Ferguson, F.; Rowe, M.T. Effect of commercial scale high temperature, short-time pasteurization on the viability of Mycobacterium paratuberculosis in naturally infected cows’ milk. Appl. Environ. Microbiol. 2002, 68, 602–607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grant, I.R.; Williams, A.G.; Rowe, M.T.; Muir, D.D. Efficacy of various pasteurization time temperature conditions in combination with homogenization on inactivation of Mycobacterium avium subsp. paratuberculosis in milk. Appl. Environ. Microbiol. 2005, 71, 2853–2861. [Google Scholar] [CrossRef] [Green Version]
- Steuer, P.; Avilez, C.; Tejeda, C.; González, N.; Ramírez-Revecco, A.; Ulloa, F.; Mella, A.; Grant, I.R.; Collins, M.T.; Salgado, M. In vitro inactivation of Mycobacterium avium subsp. paratuberculosis (MAP) by use of copper ions. BMC Microbiol. 2018, 18, 172. [Google Scholar] [CrossRef]
- Steuer, P.; Tejeda, C.; Martinez, O.; Ramirez-Reveco, A.; González, N.; Grant, I.R.; Foddai, A.C.G.; Collins, M.T.; Salgado, M. Effectiveness of copper ions against Mycobacterium avium subsp. paratuberculosis and bacterial communities in naturally contaminated raw cow’s milk. J. Appl. Microbiol. 2020. [Google Scholar] [CrossRef] [PubMed]
- Albright, J.L.; Fulwider, W.K. Dairy cattle behaviour, facilities, handling, transportation, automation and well-being. In Livestock Handling and Transport, 3rd ed.; Grandin, T., Ed.; CABI: Wallingford, UK, 2007; pp. 109–133. [Google Scholar]
- Stull, C.; Reynolds, J. Calf Welfare. Vet. Clin. Food Anim. 2008, 24, 191–203. [Google Scholar] [CrossRef]
- Dundee, L.; Grant, I.R.; Ball, H.J.; Rowe, M.T. Comparative evaluation of four decontamination protocols for the isolation of Mycobacterium avium subsp. paratuberculosis from milk. Lett. Appl. Microbiol. 2001, 33, 173–177. [Google Scholar] [CrossRef]
- Salgado, M.; Verdugo, C.; Heuer, C.; Castillo, P.; Zamorano, P. A novel low-cost method for Mycobacterium avium SUBSP. paratuberculosis DNA extraction from an automated broth culture system for a real time PCR confirmation. J. Vet. Sci. 2014, 15, 233–239. [Google Scholar] [CrossRef] [Green Version]
- Steuer, P.; Collado, B.; Avilez, C.; Tejeda, C.; Soto, J.P.; Salgado, M. Is the transmission of Mycobacterium avium subspecies paratuberculosis (MAP) infection through milk intended to feed calves an overlooked item in paratuberculosis control programs? Trop. Anim. Health Prod. 2019, 52, 89–94. [Google Scholar] [CrossRef]
- Dzieciol, M.; Volgger, P.; Khol, J.; Baumgartner, W.; Wagner, M.; Hein, I. A novel real-time PCR assay for specific detection and quantification of Mycobacterium avium subsp. paratuberculosis in milk with the inherent possibility of differentiation between viable and dead cells. BMC Res. Notes 2010, 3, 251. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, E.L.; Meier, P. Nonparametric Estimation from Incomplete Observations. J. Am. Stat. Assoc. 1958, 53, 457–481. [Google Scholar] [CrossRef]
- Zuur, A.F.; Savaliev, A.A.; Ieno, E.N. Zero Inflated Models and Generalized Linear Mixed Model with R; Imprint; Highland Statistics Ltd.: Newburgh, UK, 2012. [Google Scholar]
- Therneau, T. A Package for Survival Analysis in R. R Package Version 3.1-12. 2020. Available online: https://CRAN.R-project.org/package=survival (accessed on 27 April 2020).
- Rizopoulos, D. GLMMadaptive: Generalized Linear Mixed Models Using Adaptive Gaussian Quadrature. 2020. Available online: https://drizopoulos.github.io/GLMMadaptive/ (accessed on 27 April 2020).
- McGuirk, S. Forage Feeding and Biosecurity Issues for Cattle. 2002. Available online: https://fyi.extension.wisc.edu/forage/files/2014/01/7321973.pdf (accessed on 10 August 2020).
- Jorgensen, J.B. Survival of Mycobacterium paratuberculosis in slurry. Nord. Vet. Med. 1977, 29, 267–270. [Google Scholar]
- Katayama, N.; Tanaka, C.; Fujita, T.; Takumi, S.; Watanabe, S.; Suzuki, S. Effects of silage fermentation and ammonia treatment on activity of Mycobacterium avium subsp. paratuberculosis. Grass Sci. 2001, 47, 296–299. [Google Scholar]
- Corbett, C.; De Buck, J.; Orsel, K.; Barkema, H.W. Fecal shedding and tissue infections demonstrate transmission of Mycobacterium avium subsp. paratuberculosis in group housed dairy calves. Vet. Res. 2017, 48, 27. [Google Scholar] [CrossRef]
- Mitchell, R.M.; Medley, G.F.; Collins, M.T.; Schukken, Y.H. A meta-analysis of the effect of dose and age at exposure on shedding of Mycobacterium avium subspecies paratuberculosis (MAP) in experimentally infected calves and cows. Epidemiol. Infect. 2012, 140, 231–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corbett, C.; de Jong, M.; Orsel, K.; De Buck, J.; Barkema, H.W. Quantifying transmission of Mycobacterium avium subsp. paratuberculosis among group-housed dairy calves. Vet. Res. 2019, 50, 60. [Google Scholar] [CrossRef] [Green Version]
- Dargatz, D.; Byrum, B.; Barber, L.; Sweeney, R.; Whitlock, R.; Shulaw, W.P.; Jacobson, R.H.; Stabel, J.R. Evaluation of a commercial ELISA for diagnosis of paratuberculosis in cattle. J. Am. Vet. Med. A 2001, 218, 1163–1166. [Google Scholar] [CrossRef] [Green Version]
- Grant, I.R.; Rowe, M.T.; Dundee, L.; Hitchings, E. Mycobacterium avium ssp. paratuberculosis: Its incidence, heat resistance and detection in milk and dairy products. Int. J. Dairy Technol. 2001, 54, 2–13. [Google Scholar] [CrossRef]
- Mortier, R.; Barkema, H.; Bystrom, J.; Illanes, O.; Orsel, K.; Wolf, R.; Atkins, G.; De Buck, J. Evaluation of age-dependent susceptibility in calves infected with two doses of Mycobacterium avium subspecies paratuberculosis using pathology and tissue culture. Vet. Res. 2013, 44, 94. [Google Scholar] [CrossRef] [Green Version]
- Koets, A.; Eda, S.; Sreevatsan, S. The within host dynamics of Mycobacterium avium ssp. paratuberculosis infection in cattle: Where time and place matter. Vet. Res. 2015, 46, 61. [Google Scholar] [CrossRef] [Green Version]
- Mortier, R.; Barkema, H.; Orsel, K.; Wolf, R.; De Buck, J. Shedding patterns of dairy calves experimentally infected with Mycobacterium avium subsp. paratuberculosis. Vet. Res. 2014, 45, 71. [Google Scholar] [CrossRef] [Green Version]


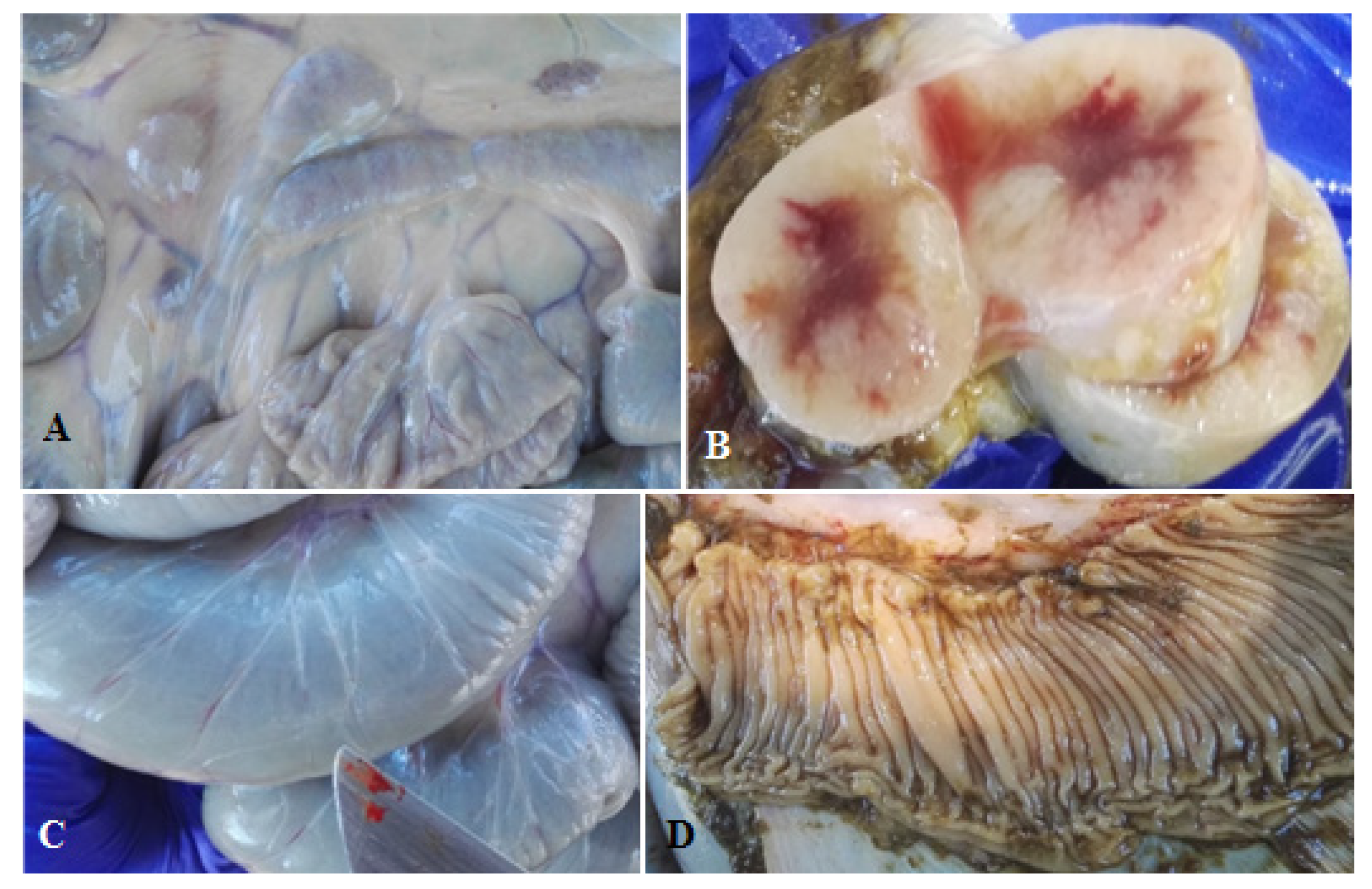{kind=link}
| Naturally Contaminated Milk (MAP bce/mL) | Copper-Treated Milk (MAP bce/mL) | |
|---|---|---|
| 7.8 × 104 | 5.7 × 104 | |
| 3.3 × 104 | ND * | |
| 5.66 × 104 | 1.86 × 104 | |
| 1.65 × 104 | ND * | |
| 6.46 × 104 | ND * | |
| 5.63 × 104 | ND * | |
| 3.80 × 104 | ND * | |
| 4.83 × 104 | ND * | |
| 2.59 × 104 | ND * | |
| 4.31 × 104 | ND * | |
| 6.70 × 102 | ND * | |
| 1.42 × 103 | ND * | |
| Mean | 3.85 × 104 | 6.3 × 103 |
| 7.8 × 104 | 5.7 × 104 | |
| 3.3 × 104 | ND * | |
| 5.66 × 104 | 1.86 × 104 | |
| 1.65 × 104 | ND * | |
| 6.46 × 104 | ND * | |
| 5.63 × 104 | ND * | |
| 3.80 × 104 | ND * | |
| 4.83 × 104 | ND * | |
| 2.59 × 104 | ND * | |
| 4.31 × 104 | ND * | |
| 6.70 × 102 | ND * | |
| 1.42 × 103 | ND * | |
| Mean | 3.85 × 104 | 6.3 × 103 |
| Experimental Group | Calf ID | 1 | 2 | 3 | 4 * | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Contaminated milk (A) | 270817 | ||||||||||||
| Contaminated milk (A) | 50917 | ||||||||||||
| Contaminated milk (A) | 300917 | ||||||||||||
| Contaminated milk (A) | 11017 | ||||||||||||
| Contaminated milk (A) | 21017 | ||||||||||||
| Copper-treated milk (B) | 311017 | ||||||||||||
| Copper-treated milk (B) | 11117 | ||||||||||||
| Copper-treated milk (B) | 21117 | ||||||||||||
| Copper-treated milk (B) | 31117 | ||||||||||||
| Copper-treated milk (B) | 41117 | ||||||||||||
| Milk replacer (C) | 31017 | ||||||||||||
| Milk replacer (C) | 41017 | ||||||||||||
| Milk replacer (C) | 51017 | ||||||||||||
| Milk replacer (C) | 61017 | ||||||||||||
| UHT-milk (D) | 121017 | ||||||||||||
| UHT-milk (D) | 131017 | ||||||||||||
| UHT-milk (D) | 141017 |
| Groups | ||||
|---|---|---|---|---|
| Blood Analyte | Contaminated Milk | Copper-Treated Milk | Milk Replacer | UHT Milk |
| GGT (3–39 U/L) | 21 ± 2.6 | 21.3 ± 1.8 | 24.9 ± 3.7 | 27.0 ± 4.4 |
| GD (2–28 U/L) | 34 ± 4.3 | 35.4 ± 23 | 38.0 ± 14.3 | 58 ± 19.6 |
| AST (25–125 U/L) | 94.5 ± 12.1 | 91.6 ± 6.3 | 101.9 ± 14.1 | 104.0 ± 2.5 |
| Plasma Cu (10–22 µmol/L) | 9 ± 1.5 | 7.5 ± 1.8 | 8 ± 1.4 | 8.7 ± 1.5 |
| Hepatic Cu (25–100 ppm) | 25.4 ± 8.2 | 43.8 ± 17 | 45 ± 9.5 | 46.3 ± 20.4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Steuer, P.; Tejeda, C.; Moroni, M.; Verdugo, C.; Collins, M.T.; Salgado, M. Attempted Control of Paratuberculosis in Dairy Calves by Only Changing the Quality of Milk Fed to Calves. Animals 2021, 11, 2569. https://doi.org/10.3390/ani11092569
Steuer P, Tejeda C, Moroni M, Verdugo C, Collins MT, Salgado M. Attempted Control of Paratuberculosis in Dairy Calves by Only Changing the Quality of Milk Fed to Calves. Animals. 2021; 11(9):2569. https://doi.org/10.3390/ani11092569
Chicago/Turabian StyleSteuer, Pamela, Carlos Tejeda, Manuel Moroni, Cristobal Verdugo, Michael Thomas Collins, and Miguel Salgado. 2021. "Attempted Control of Paratuberculosis in Dairy Calves by Only Changing the Quality of Milk Fed to Calves" Animals 11, no. 9: 2569. https://doi.org/10.3390/ani11092569
APA StyleSteuer, P., Tejeda, C., Moroni, M., Verdugo, C., Collins, M. T., & Salgado, M. (2021). Attempted Control of Paratuberculosis in Dairy Calves by Only Changing the Quality of Milk Fed to Calves. Animals, 11(9), 2569. https://doi.org/10.3390/ani11092569