
Aberrant Lighting Causes Anxiety-like Behavior in Mice but Curcumin Ameliorates the Symptoms

 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Drugs and Biochemical Reagents
2.3. Drug Preparation and Experiment Design
3. Behavioral Studies
3.1. Open Field Test
- (1)
- Percentage of freezing time (percentage of times the animal stands still in one place).
- (2)
- Frequency of grooming (total number of times animals scratch and lick their body without motion).
- (3)
- Frequency of changes in body posture (total number of times animals exhibit elongated body posture).
3.2. Brain Tissue Sample Collection
3.3. Total RNA Extraction and cDNA Synthesis
3.4. Quantitative RT-PCR
3.5. Statistical Analysis
4. Results
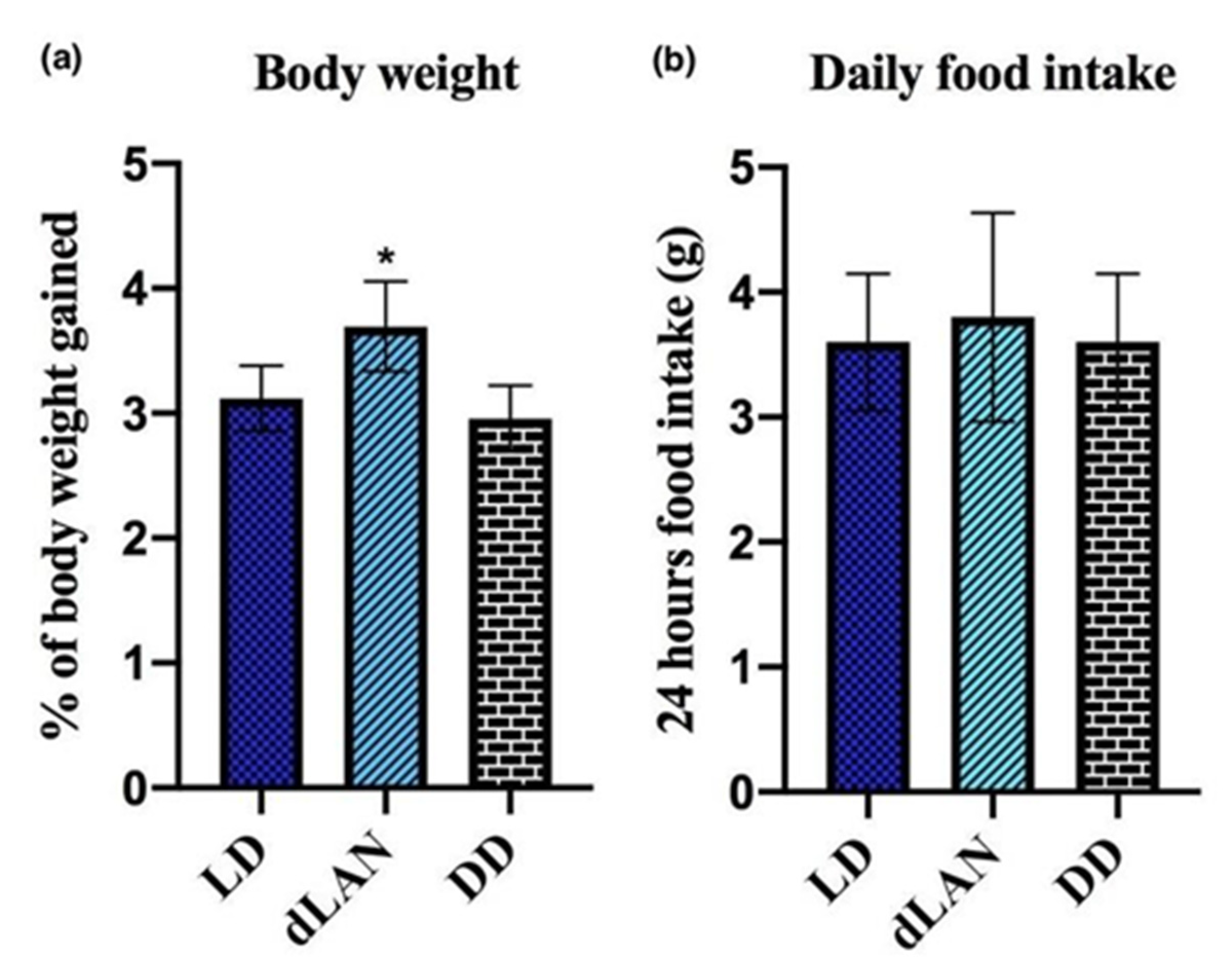
4.1. Exposure to dLAN Increases Body Weight in Mice
4.2. Curcumin Is Ineffective in Controlling Increased Body Weight Induced Due to dLAN Exposure
4.3. Aberrant Lighting (dLAN and DD) Caused Anxiety-like Behavior in Mice
4.4. Curcumin Improves the Anxiety-Like Behavior in Mice Exposed to dLAN and DD
4.5. Influence of Aberrant Lighting (dLAN and DD) on the Hippocampal Sleep Gene PER1
4.6. Curcumin Increases the Expression of the Hippocampal Sleep Gene PER1 in Mice Exposed to dLAN and DD
5. Discussion
6. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nakamaru-Ogiso, E.; Miyamoto, H.; Hamada, K.; Tsukada, K.; Takai, K. Novel biochemical manipulation of brain serotonin reveals a role of serotonin in the circadian rhythm of sleep-wake cycles. Eur. J. Neurosci. 2012, 35, 1762–1770. [Google Scholar] [CrossRef] [PubMed]
- Alibhai, F.; Tsimakouridze, E.V.; Reitz, C.J.; Pyle, G.; Martino, T.A. Consequences of Circadian and Sleep Disturbances for the Cardiovascular System. Can. J. Cardiol. 2015, 31, 860–872. [Google Scholar] [CrossRef] [PubMed]
- Chakir, I.; Dumont, S.; Pévet, P.; Ouarour, A.; Challet, E.; Vuillez, P. Pineal melatonin is a circadian time-giver for leptin rhythm in Syrian hamsters. Front Neurosci. 2015, 9, 190. [Google Scholar] [CrossRef] [Green Version]
- Ben-Hamo, M.; Larson, T.A.; Duge, L.S.; Sikkema, C.; Wilkinson, C.W.; De La Iglesia, H.O.; González, M.M.C. Circadian Forced Desynchrony of the Master Clock Leads to Phenotypic Manifestation of Depression in Rats. Eneuro 2016, 3. [Google Scholar] [CrossRef]
- Bailey, S.M.; Udoh, U.S.; Young, M.E. Circadian regulation of metabolism. J. Endocrinol. 2014, 222, R75–R96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lunn, R.M.; Blask, D.E.; Coogan, A.N.; Figueiro, M.G.; Gorman, M.R.; Hall, J.E.; Hansen, J.; Nelson, R.J.; Panda, S.; Smolensky, M.H.; et al. Health consequences of electric lighting practices in the modern world: A report on the National Toxicology Program’s workshop on shift work at night, artificial light at night, and circadian disruption. Sci. Total Environ. 2017, 607, 1073–1084. [Google Scholar] [CrossRef] [Green Version]
- Fonken, L.K.; Nelson, R.J. Illuminating the deleterious effects of light at night. F1000 Med. Rep. 2011, 3, 18. [Google Scholar] [CrossRef]
- Gerstner, J.R. On the Evolution of Memory: A Time for Clocks. Front. Mol. Neurosci. 2012, 5, 23. [Google Scholar] [CrossRef] [Green Version]
- Fonken, L.K.; Workman, J.L.; Walton, J.C.; Weil, Z.M.; Morris, J.S.; Haim, A.; Nelson, R.J. Light at night increases body mass by shifting the time of food intake. Proc. Natl. Acad. Sci. USA 2010, 107, 18664–18669. [Google Scholar] [CrossRef] [Green Version]
- Bedrosian, T.A.; Fonken, L.; Walton, J.; Haim, A.; Nelson, R.J. Dim light at night provokes depression-like behaviors and reduces CA1 dendritic spine density in female hamsters. Psychoneuroendocrinology 2011, 36, 1062–1069. [Google Scholar] [CrossRef]
- Fonken, L.K.; Kitsmiller, E.; Smale, L.; Nelson, R.J. Dim Nighttime Light Impairs Cognition and Provokes Depressive-Like Responses in a Diurnal Rodent. J. Biol. Rhythm. 2012, 27, 319–327. [Google Scholar] [CrossRef]
- LeGates, T.; Altimus, C.M.; Wang, H.; Lee, H.-K.; Yang, S.; Zhao, H.; Kirkwood, A.; Weber, E.T.; Hattar, S. Aberrant light directly impairs mood and learning through melanopsin-expressing neurons. Nature 2012, 491, 594–598. [Google Scholar] [CrossRef] [Green Version]
- Tapia-Osorio, A.; Salgado-Delgado, R.; Angeles-Castellanos, M.; Escobar, C. Disruption of circadian rhythms due to chronic constant light leads to depressive and anxiety-like behaviors in the rat. Behav. Brain Res. 2013, 252, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Fonken, L.K.; Lieberman, R.A.; Weil, Z.M.; Nelson, R.J. Dim Light at Night Exaggerates Weight Gain and Inflammation Associated With a High-Fat Diet in Male Mice. Endocrinology 2013, 154, 3817–3825. [Google Scholar] [CrossRef] [Green Version]
- Moriya, S.; Tahara, Y.; Sasaki, H.; Ishigooka, J.; Shibata, S. Housing under abnormal light–dark cycles attenuates day/night expression rhythms of the clock genes Per1, Per2, and Bmal1 in the amygdala and hippocampus of mice. Neurosci. Res. 2015, 99, 16–21. [Google Scholar] [CrossRef]
- Vogel, M.; Braungardt, T.; Meyer, W.; Schneider, W. The effects of shift work on physical and mental health. J. Neural Transm. 2012, 119, 1121–1132. [Google Scholar] [CrossRef]
- Morrissette, D.A. Twisting the night away: A review of the neurobiology, genetics, diagnosis, and treatment of shift work disorder. CNS Spectr. 2013, 18, 42–54. [Google Scholar] [CrossRef]
- Caruso, C.C. Negative Impacts of Shiftwork and Long Work Hours. Rehabil. Nurs. 2013, 39, 16–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Panagiotou, M.; Deboer, T. Effects of Chronic Dim-light-at-night Exposure on Sleep in Young and Aged Mice. Neuroscience 2019, 426, 154–167. [Google Scholar] [CrossRef]
- Russart, K.L.; Chbeir, S.A.; Nelson, R.J.; Magalang, U.J. Light at night exacerbates metabolic dysfunction in a polygenic mouse model of type 2 diabetes mellitus. Life Sci. 2019, 231, 116574. [Google Scholar] [CrossRef]
- Namgyal, D.; Chandan, K.; Sultan, A.; Aftab, M.; Ali, S.; Mehta, R.; El-Serehy, H.A.; Al-Misned, F.A.; Sarwat, M. Dim Light at Night Induced Neurodegeneration and Ameliorative Effect of Curcumin. Cells 2020, 9, 2093. [Google Scholar] [CrossRef] [PubMed]
- Mendoza, J.; Albrecht, U.; Challet, E. Behavioural food anticipation in clock genes deficient mice: Confirming old phenotypes, describing new phenotypes. Genes Brain Behav. 2010, 9, 467–477. [Google Scholar] [CrossRef]
- Kim, M.; De La Peña, J.B.; Cheong, J.H.; Kim, H.J. Neurobiological Functions of the Period Circadian Clock 2 Gene, Per2. Biomol. Ther. 2018, 26, 358–367. [Google Scholar] [CrossRef] [PubMed]
- Lavebratt, C.; Sjöholm, L.K.; Partonen, T.; Schalling, M.; Forsell, Y. PER2 variantion is associated with depression vulnerability. Am. J. Med Genet. Part B Neuropsychiatr. Genet. 2009, 153B, 570–581. [Google Scholar] [CrossRef]
- Obayashi, K.; Saeki, K.; Iwamoto, J.; Okamoto, N.; Tomioka, K.; Nezu, S.; Ikada, Y.; Kurumatani, N. Exposure to Light at Night, Nocturnal Urinary Melatonin Excretion, and Obesity/Dyslipidemia in the Elderly: A Cross-Sectional Analysis of the HEIJO-KYO Study. J. Clin. Endocrinol. Metab. 2013, 98, 337–344. [Google Scholar] [CrossRef]
- Namgyal, D.; Sarwat, M. Saffron as a Neuroprotective Agent. In Saffron: The Age-Old Panacea in a New Light; Sarwat, M., Sumaiya, S., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 93–102. [Google Scholar]
- Xu, Y.; Ku, B.; Tie, L.; Yao, H.; Jiang, W.; Ma, X.; Li, X. Curcumin reverses the effects of chronic stress on behavior, the HPA axis, BDNF expression and phosphorylation of CREB. Brain Res. 2006, 1122, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Hamaguchi, T.; Ono, K.; Yamada, M. Curcumin and Alzheimer’s disease. CNS Neurosci. Ther. 2010, 16, 285–297. [Google Scholar] [CrossRef]
- Huang, Z.; Zhong, X.-M.; Li, Z.-Y.; Feng, C.-R.; Pan, A.-J.; Mao, Q.-Q. Curcumin reverses corticosterone-induced depressive-like behavior and decrease in brain BDNF levels in rats. Neurosci. Lett. 2011, 493, 145–148. [Google Scholar] [CrossRef]
- Tiwari, S.K.; Agarwal, S.; Tripathi, A.; Chaturvedi, R.K. Bisphenol-A Mediated Inhibition of Hippocampal Neurogenesis Attenuated by Curcumin via Canonical Wnt Pathway. Mol. Neurobiol. 2015, 53, 3010–3029. [Google Scholar] [CrossRef]
- Bassani, T.B.; Turnes, J.M.; Moura, E.L.; Bonato, J.M.; Cóppola-Segovia, V.; Zanata, S.M.; Oliveira, R.M.; Vital, M.A. Effects of curcumin on short-term spatial and recognition memory, adult neurogenesis and neuroinflammation in a streptozotocin-induced rat model of dementia of Alzheimer’s type. Behav. Brain Res. 2017, 335, 41–54. [Google Scholar] [CrossRef]
- Seibenhener, M.L.; Wooten, M.C. Use of the Open Field Maze to Measure Locomotor and Anxiety-like Behavior in Mice. J. Vis. Exp. 2015, e52434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sarwat, M.; Naqvi, A.R. Heterologous expression of rice calnexin (OsCNX) confers drought tolerance in Nicotiana tabacum. Mol. Biol. Rep. 2013, 40, 5451–5464. [Google Scholar] [CrossRef]
- Stevens, R.G. Light-at-night, circadian disruption and breast cancer: Assessment of existing evidence. Int. J. Epidemiol. 2009, 38, 963–970. [Google Scholar] [CrossRef]
- Kohyama, J. A newly proposed disease condition produced by light exposure during night: Asynchronization. Brain Dev. 2009, 31, 255–273. [Google Scholar] [CrossRef]
- Driesen, K.; Jansen, N.W.H.; Kant, I.; Mohren, D.C.L.; Van Amelsvoort, L.G.P.M. Depressed mood in the working population: Associations with work schedules and working hours. Chrono Int. 2010, 27, 1062–1079. [Google Scholar] [CrossRef]
- Ha, M.; Park, J. Shiftwork and Metabolic Risk Factors of Cardiovascular Disease. J. Occup. Health 2005, 47, 89–95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schaap, E.; Sagong, C.; Castelan, A.S.C.; Sayed, J.; Roth, T.; Drake, C.L.; Cheng, P. 0006 The Role of Stress in Sleep in Night Shift Workers: Going Beyond Circadian Misalignment. Sleep 2020, 43, A2–A3. [Google Scholar] [CrossRef]
- Cannizzaro, E.; Cirrincione, L.; Mazzucco, W.; Scorciapino, A.; Catalano, C.; Ramaci, T.; Ledda, C.; Plescia, F. Night-Time Shift Work and Related Stress Responses: A Study on Security Guards. Int. J. Environ. Res. Public Health 2020, 17, 562. [Google Scholar] [CrossRef] [Green Version]
- Lamont, E.W.; Harbour, V.L.; Barry-Shaw, J.; Diaz, L.R.; Robinson, B.; Stewart, J.; Amir, S. Restricted access to food, but not sucrose, saccharine, or salt, synchronizes the expression of Period2 protein in the limbic forebrain. Neuroscience 2006, 144, 402–411. [Google Scholar] [CrossRef] [Green Version]
- Namgyal, D.; Ali, S.; Mehta, R.; Sarwat, M. The neuroprotective effect of curcumin against Cd-induced neurotoxicity and hippocampal neurogenesis promotion through CREB-BDNF signaling pathway. Toxicology 2020, 442, 152542. [Google Scholar] [CrossRef] [PubMed]
- Namgyal, D.; Chandan, K.; Mehta, R.; Sarwat, M. Curcumin improves the behaviour and memory in mice by modulating the core circadian genes and their associated micro-RNAs. J. Pharmacol. Pharmacother. 2020, 11, 44–52. [Google Scholar]
- Parkes, K.R. Shift work and age as interactive predictors of body mass index among offshore workers. Scand. J. Work. Environ. Health 2002, 28, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Borniger, J.C.; Maurya, S.K.; Periasamy, M.; Nelson, R.J. Acute dim light at night increases body mass, alters metabolism, and shifts core body temperature circadian rhythms. Chronobiol. Int. 2014, 31, 917–925. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, M.M.C.; Aston-Jones, G. Light deprivation damages monoamine neurons and produces a depressive behavioral phenotype in rats. Proc. Natl. Acad. Sci. USA 2008, 105, 4898–4903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González, M.M.C. Dim Light at Night and Constant Darkness: Two Frequently Used Lighting Conditions That Jeopardize the Health and Well-being of Laboratory Rodents. Front. Neurol. 2018, 9, 609. [Google Scholar] [CrossRef] [Green Version]
- Nelson, R.J.; Chbeir, S. Dark matters: Effects of light at night on metabolism. Proc. Nutr. Soc. 2018, 77, 223–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Escalante-Covarrubias, Q.; Aguilar-Arnal, L. Environmental regulation of metabolism through the circadian clock. Curr. Opin. Toxicol. 2018, 8, 93–101. [Google Scholar] [CrossRef]
- Borniger, J.C.; Weil, Z.; Zhang, N.; Nelson, R.J. Dim Light at Night Does Not Disrupt Timing or Quality of Sleep in Mice. Chrono Int. 2013, 30, 1016–1023. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Zhang, H.-Q.; Liang, X.-Y.; Zhang, H.-F.; Zhang, T.; Liu, F.-E. Melatonin ameliorates cognitive impairment induced by sleep deprivation in rats: Role of oxidative stress, BDNF and CaMKII. Behav. Brain Res. 2013, 256, 72–81. [Google Scholar] [CrossRef]
- Miyazaki, K.; Wakabayashi, M.; Chikahisa, S.; Séi, H.; Ishida, N. PER2 controls circadian periods through nuclear localization in the suprachiasmatic nucleus. Genes Cells 2007, 12, 1225–1234. [Google Scholar] [CrossRef] [Green Version]
- Hampp, G.; Albrecht, U. The circadian clock and mood-related behavior. Commun. Integr. Biol. 2008, 1, 1–3. [Google Scholar] [CrossRef]
- Logan, R.W.; Edgar, N.; Gillman, A.G.; Hoffman, D.; Zhu, X.; McClung, C.A. Chronic Stress Induces Brain Region-Specific Alterations of Molecular Rhythms that Correlate with Depression-like Behavior in Mice. Biol. Psychiatry 2015, 78, 249–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verwey, M.; Khoja, Z.; Stewart, J.; Amir, S. Differential regulation of the expression of Period2 protein in the limbic forebrain and dorsomedial hypothalamus by daily limited access to highly palatable food in food-deprived and free-fed rats. Neuroscience 2007, 147, 277–285. [Google Scholar] [CrossRef] [Green Version]
- Grimaldi, B.; Bellet, M.M.; Katada, S.; Astarita, G.; Hirayama, J.; Amin, R.H.; Granneman, J.G.; Piomelli, D.; Leff, T.; Sassone-Corsi, P. PER2 Controls Lipid Metabolism by Direct Regulation of PPARγ. Cell Metab. 2010, 12, 509–520. [Google Scholar] [CrossRef] [PubMed] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| S. Number | Groups | Experiment Conditions | Treatment | Duration |
|---|---|---|---|---|
| 1 | Light/Dark (LD) control | 12 h:12 h light/dark [~150 lux]/[~0 lux] | 1% CMC | 3 weeks |
| 2 | Dim Light at Night (dLAN) control | 12 h:12 h light/dim light [~150 lux]/[~5 lux] | 1% CMC | 3 weeks |
| 3 | Complete Darkness (DD) control | 12 h:12 h dark/dark [~0 lux]/[~0 lux] | 1% CMC | 3 weeks |
| 4 | dLAN + Cur50 | 12 h:12 h light/dim light [~150 lux]/[~5 lux] | 50 mg/kg curcumin | 3 weeks |
| 5 | dLAN + Cur100 | 12 h:12 h light/dim light [~150 lux]/[~5 lux] | 100 mg/kg curcumin | 3 weeks |
| 6 | dLAN + Cur150 | 12 h:12 h light/dim light [~150 lux]/[~5 lux] | 150 mg/kg curcumin | 3 weeks |
| 7 | DD + Cur50 | 12 h:12 h dark/dark [~0 lux]/[~0 lux] | 50 mg/kg curcumin | 3 weeks |
| 8 | DD + Cur100 | 12 h:12 h dark/dark [~0 lux]/[~0 lux] | 100 mg/kg curcumin | 3 weeks |
| 9 | DD + Cur150 | 12 h:12 h dark/dark [~0 lux]/[~0 lux] | 150 mg/kg curcumin | 3 weeks |
| S. Number | Gene | Forward Primer (5′-3′) | Reverse Primer (5′-3′) |
|---|---|---|---|
| 1 | PER1 | TTGGCAGGCTTCGTGGACTTG | GCGGGAACGCTTTGCTTTAGAT |
| 2 | β-actin | TGGTGGGTATGGGTCAGAAGGACTC | CATGGCTGGGGTGTTGAAGGTCTCA |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Namgyal, D.; Chandan, K.; Ali, S.; Ahmad, A.; Hashim, M.J.; Sarwat, M. Aberrant Lighting Causes Anxiety-like Behavior in Mice but Curcumin Ameliorates the Symptoms. Animals 2021, 11, 2590. https://doi.org/10.3390/ani11092590
Namgyal D, Chandan K, Ali S, Ahmad A, Hashim MJ, Sarwat M. Aberrant Lighting Causes Anxiety-like Behavior in Mice but Curcumin Ameliorates the Symptoms. Animals. 2021; 11(9):2590. https://doi.org/10.3390/ani11092590
Chicago/Turabian StyleNamgyal, Dhondup, Kumari Chandan, Sher Ali, Ajaz Ahmad, Maha J. Hashim, and Maryam Sarwat. 2021. "Aberrant Lighting Causes Anxiety-like Behavior in Mice but Curcumin Ameliorates the Symptoms" Animals 11, no. 9: 2590. https://doi.org/10.3390/ani11092590
APA StyleNamgyal, D., Chandan, K., Ali, S., Ahmad, A., Hashim, M. J., & Sarwat, M. (2021). Aberrant Lighting Causes Anxiety-like Behavior in Mice but Curcumin Ameliorates the Symptoms. Animals, 11(9), 2590. https://doi.org/10.3390/ani11092590