1. Introduction
Currently, there is a concept to keep hens for extended periods in production, aiming to increase profitability and sustainability. Despite the benefits of doing so, controlling the excess body fat and eggshell quality in old laying hens is reported to be the main concern on poultry farms [
1,
2]. The body fat and egg components are influenced by the feed offered [
3] and eggshell quality reduces as egg weight increases, which may be partially controlled with nutritional strategies [
4,
5]. In addition, the feed offered to hens during the rearing phase may affect the development of reproductive organs [
6], influencing the long-term laying cycle.
The ability of laying hens to overcome a nutritional deficiency or an imbalanced diet is not completely elucidated. Some effort was made to investigate the effects of a previous feed on the laying cycle phase [
7,
8,
9] but little or no knowledge is available in the literature describing how a modern hen may deal with a deficient diet during the growth and its impact in a long-term laying cycle. This information is convenient for poultry nutritionists because they often change the feed formula to improve the economic return and sustainability of egg production farms. In this sense, protein is frequently investigated in poultry nutrition given its importance for growth [
10,
11], egg production [
12,
13], economic return [
14,
15], and sustainability of the farm [
16,
17]. Because amino acids are the basic constituents of proteins and that essential amino acids should be offered in the feed in a proper ratio with lysine [
18], it seems reasonable to investigate the effects of balanced protein in a long-term egg production cycle.
Understanding the dynamics of body and egg components represents a step towards an effective way to improve long-term egg production given feeds should be formulated based on physiological needs and the response of laying hens. In a conventional poultry house, the feed offered is the only source of energy and nutrients for a laying hen. Therefore, a change in voluntary feed intake is the only mechanism that a hen can use to consume a proper amount of all nutrients. If they fail to do so, body and egg components are expected to change [
6,
19] along with the egg production [
20]. This highlights the importance to elucidate the dynamics of body and egg composition over different sets of nutrition scenarios.
The effects of dietary balanced protein for hens in the rearing phase and its cumulated influence in the long-term laying cycle have not been investigated so far. In the context aforementioned, we hypothesized that balanced protein levels affect the body and egg components leading to a shift in the long-term laying cycle; thus, the aim of the present research was to describe how laying hens respond to three levels of dietary balanced protein from 8 to 102 weeks old.
2. Materials and Methods
2.1. Ethics Approval
All procedures described were approved by the Ethical Committee on the Use of Animals of the School of Agrarian and Veterinary Sciences, São Paulo State University (UNESP), Jaboticabal, São Paulo, Brazil (Process 012598/2018; approved on 14 February 2019).
2.2. Bird Husbandry
A total of 600 Lohmann LITE LSL-NA were obtained from a breeding company (Planalto Postura LTDA. Uberlândia, MG. Brazil). The hatchlings were raised in conventional cages from 1 to 7 weeks before being moved to wire rearing cages (375 cm2 per pullet) from 8 to 18 weeks. At 19 weeks, hens were transferred to wire-laying cages (563 cm2 per hen). All cages were equipped with trough feeders and nipple drinkers. Hens received a corn-soybean meal-based diet to meet or exceed breeding company recommendations from 1 to 7 weeks old. A feed program with three feeds was offered from 8 to 18 weeks, for grower (8–11 weeks), developer (12–15 weeks), and pre-laying phases (16–18 weeks). At the laying phase, a feed program with five feeds was formulated according to the breeding company recommendations: layer one (19–26 w-old), layer two (27–46 w-old), layer three (47–66 w-old), layer four (67–82 w-old), and layer five (83–102 w-old). Birds had free access to feed and fresh water throughout the trial. The lighting program adopted was 24 L in the first week, reduced gradually to 12L:12D up to 10-w-old, which was maintained until the pullets achieved 5% of egg production (20-w-old). After the onset of egg production, the lighting program was gradually increased from 12–16 h of light by adding one hour per week and was then kept constant up to 102 weeks of age. In the period of 0–7, 8–18, and 19–102 w-old, the maximum temperatures recorded were 31, 26, and 27 °C, while the minimum were 24, 17, and 19 °C, respectively. The maximum relative humidity of the air was 77, 79, and 84% while the minimum records were 51, 57, and 50%, respectively for the same phases. One laying cycle was considered as 28 consecutive days.
2.3. Experimental Design and Feeds
Three treatments were randomly assigned to 30 experimental units of 20 pullets each, totalling ten replicates per treatment, performing a completed randomized design. Treatments consisted of three dietary levels of balanced protein (BP): 1—standard feed (S), formulated to meet or exceed breeding company recommendations; 2—reduction of 20% in dietary balanced protein (L), in reference to the S feed; 3—increase of 20% in dietary balanced protein (H), in reference to the S feed. Dietary balanced protein was defined as a constant ratio of essential amino acids with lysine [
21] and the ratio was the same proposed by the breeding company [
22]. Standardized ileal digestible lysine (SID-Lys) was the reference to produce the three levels of dietary balanced protein (
Table 1).
In the rearing phase, the S-BP group consumed a feed containing 0.80, 0.70, and 0.74 % of SID-Lys for the grower, developer, and pre-layer phase, respectively. The S-BP feeds in the laying phase contained 0.68, 0.66, 0.63, 0.61, and 0.58% of SID-Lys, respectively for each layer phase (
Table 2). The remaining nutrients and energy in the feed were as recommended by the guideline [
22].
2.4. Performance Data and Egg Components Measurement
The number of eggs produced, and mortality were daily recorded. Every week, all eggs produced were weighed and the egg mass (rate of egg produced × egg weight) was calculated. The feed leftovers were weighed fortnightly and adjusted by mortality to calculate the food intake. The cumulative feed intake was expressed on g/bird for the rearing (eight to 18 w-old) and the rearing plus laying phases (8–102 w-old). Hen-housed egg production was calculated based on the number of eggs produced in the entire experiment period per number of housed hens at 19 w-old.
2.5. Body Composition
On the first day of the trial, eight birds per treatment were randomly selected and identified for body composition measurements by dual-energy X-ray absorptiometry (Hologic-QDR
® model 13.4.2., Marlborough, MA, USA). Throughout the experiment, the same hen was scanned on the last day of every feeding phase. Prior to each scan, hens were fasted for five hours, weighed, anesthetized with isoflurane (2%) diluted in 100% of oxygen, and positioned in dorsal decubitus with the wings and legs flexed [
23]. Measures collected were fat mass (g), lean mass (water + protein content, g), bone mineral content (g), and bone mineral density (g/cm²). Data collected were converted to contents of body protein, fat, and ash by applying the equations published by Alves et al. [
23].
2.6. Egg Components
At the end of each laying cycle (28 days), a total of nine eggs per experimental unit were collected (three eggs per three sequential days). On each day, the eggs were broken apart individually to measure the albumen, yolk, and shell weights. Before measurement, the eggshell was washed with tap water and dried using a forced oven at 55 °C for 24 h. The percentages of albumen, yolk, and eggshell were then calculated.
2.7. Statistical Analysis
The collected data were examined for outliers, normality, and homoscedasticity. The data of cumulative feed intake and hen-housed egg production were analysed as One-Way ANOVA with a Tukey test to evaluate the differences between dietary balanced protein levels, using a generalized linear model. Two-factor repeated measure design was employed to determine the effects of dietary balanced protein over time, using a mixed model. One factor is represented by the three groups receiving the different series of dietary balanced protein feeds and the other factor is the age of hens. Each experimental unit was the repeated measures factor. Differences were considered to be significant at a probability of 5%. The Statistical Analysis System (SAS Institute Inc., Cary, NC, USA) was used to perform both a One-Way ANOVA and the Two-factor repeated measure analyses procedures. The data was analysed considering 21 cycles of four weeks each.
To test whether the responses differed between dietary balanced protein levels over time, non-linear regression with groups was used, the groups being the dietary balanced protein [
24]. The average data per replicate were treated as the experimental unit. Two exponential models were applied and that with the lower Akaike information criterion [
25] value was used to describe the response variable. The model used were:
where
A and
C are the y-intercept and slope of the linear segment, respectively,
B is the y-intercept of the exponential segment, and
R is the exponential base.
where
A1 +
B1 is the y-intercept, and
R1 is the exponential base.
3. Results
The reduction and increase of dietary balanced protein in the laying feed did not affect the cumulative feed intake (
p > 0.05,
Table 3), being on average 4.44 kg (
p = 0.986) and 67.7 kg of feed per bird (
p = 0.485) in the growth and whole period, respectively. The reduction of dietary balanced protein affected the hen-housed egg production (
p < 0.01), with similar results between hens from the S and H groups (
p > 0.05).
There was an interaction between dietary balanced protein and hens age for feed intake, egg production, and egg mass (
p < 0.05,
Table 4). For feed intake, differences between treatments were observed only at 26 weeks of age (
p < 0.05). For egg production, differences were observed mainly at the beginning (first three laying cycles) and the end (after 74 w-old) of the laying cycle, whereas for egg mass the differences between groups of hens were consistent during the whole experimental period (
Table 4).
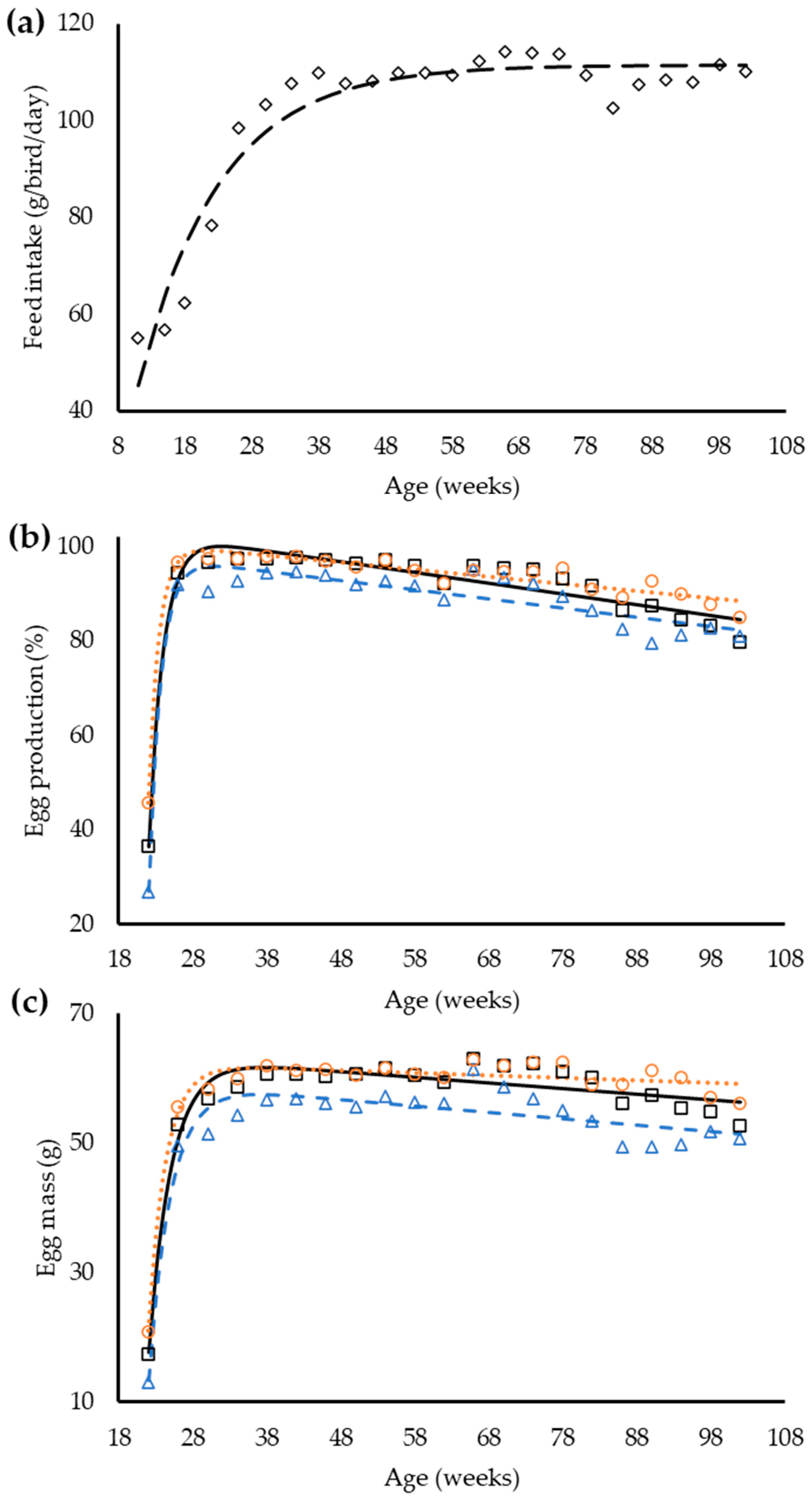
The exponential equation was used to demonstrate the changes in feed intake and the line plus exponential equation had the best fit for egg production and egg mass (
Table 5). The regression with groups identified that a single equation could be used to describe the feed intake between groups, whereas for egg production and egg mass the regression analysis indicates a necessity for different equations for laying hens inside each dietary balanced protein group (
Figure 1).
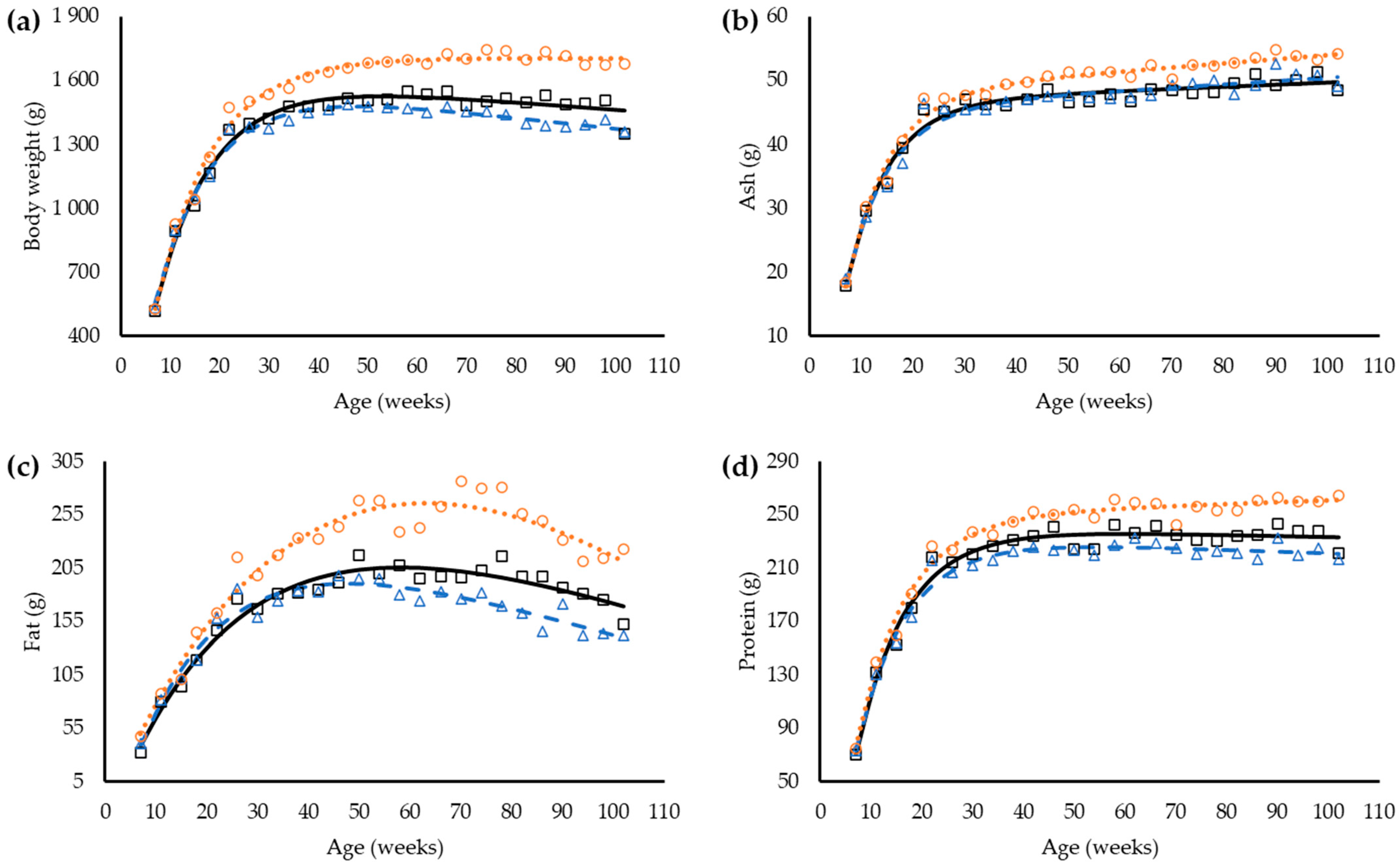
Differences in body weight influenced by dietary balanced protein were observed in 30 w-old hens (
p < 0.05,
Table 6), with heavier hens in the H group, followed by the S and L groups, respectively. Laying hens in the higher dietary balanced protein feed was fatter (
p < 0.05) from 38 w-old and forward (
Table 6).
Body contents of ash and protein increased along time for all groups (
p < 0.05,
Table 7). For body ash, differences were observed from 50 w-old and body protein from 30 w-old, and still relatively constant until the end of the trial.
The linear plus exponential function had the best fit for body weight and body components, and it was used to investigate the differences between hens consuming the different feeds (
Table 8). A common coefficient R-value can be used to describe the changes in growth (
p < 0.05), despite the levels of dietary balanced protein, except for fat (
p < 0.05). However, the coefficients A, B, and C are different between groups (
p < 0.05) and specific values are necessary to properly describe the changes in body weight and body composition between hens consuming the different levels of dietary balanced protein (
Figure 2).
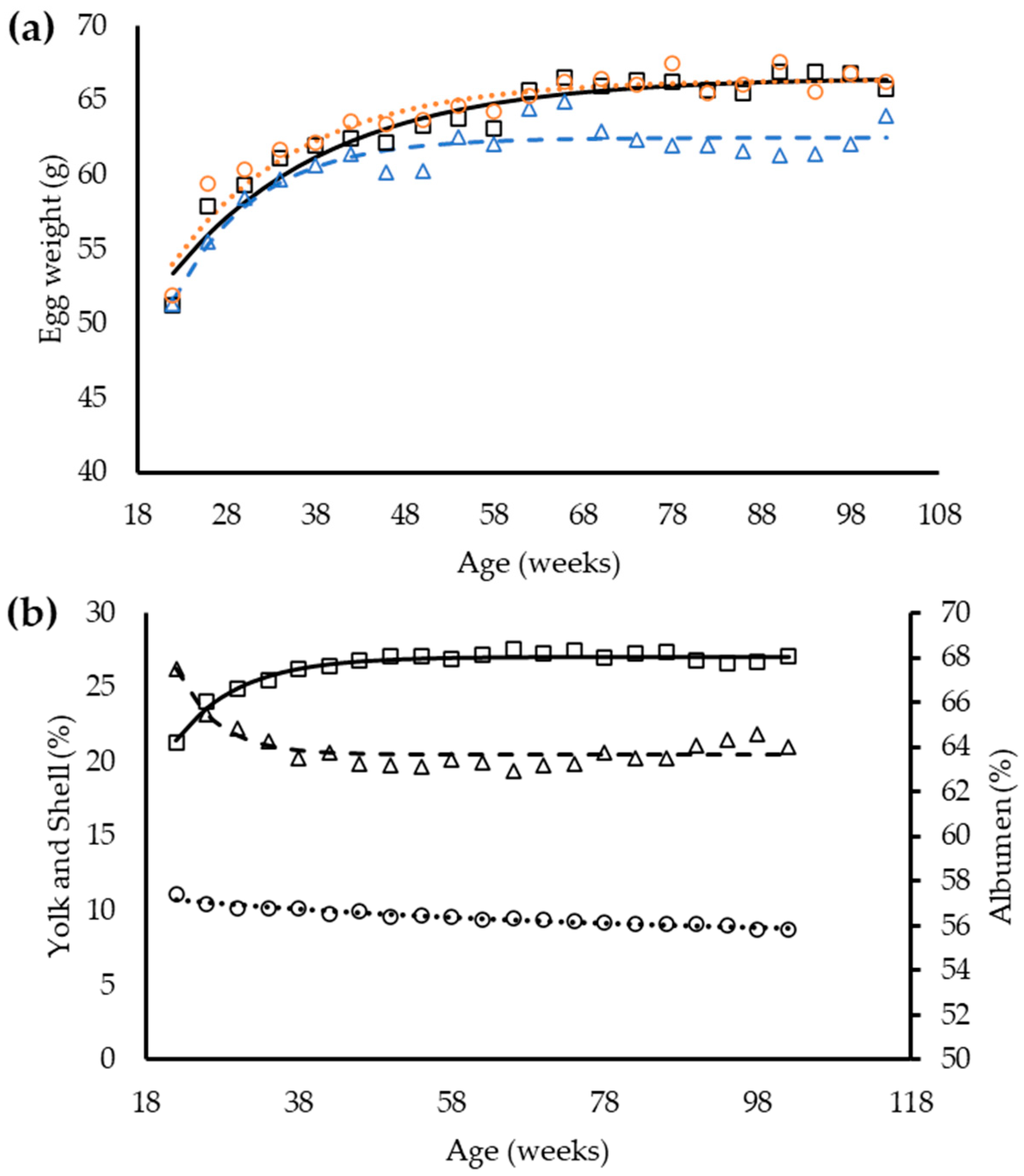
Overall, egg weight of laying hens was affected by dietary balance protein content (
p < 0.05,
Table 9). At 26 w-old, hens consuming the H feed produced heavier eggs, followed by hens from the S and L groups, respectively. Notably, egg weight was similar between groups of hens from 30 to 42 w-old and from 54 to 66 w-old (
p < 0.05). The yolk percentage increased in all feed treatments as the hens aged (
p < 0.05).
Different from those observed for yolk, the albumen and eggshell percentages reduced with time (
p < 0.05,
Table 10). Nevertheless, egg components were similar between groups of hens (
p > 0.05) with a tendency for yolk percentage (
p < 0.06).
The exponential function had the best fit for egg weight and egg components (
Table 11). The analysis indicates that all dietary balanced protein contents used in this study affected the egg weight (
p < 0.05) and all equation coefficients need to be changed to estimate the egg weight of hens according to dietary balanced protein. The range in dietary balanced protein levels applied in this study was not sufficient to change the concentration in egg components (
p < 0.05); therefore, a single exponential equation was used for each egg component. A tendency was observed for yolk percentage (
p = 0.06), suggesting an influence of dietary balanced protein in this egg component. The equation used to describe the albumen percentage had a low R
2 value, mainly because a drop in albumen percentage was observed around 68 w-old and followed by a consecutive increase (
Figure 3), which was poorly predicted with the exponential equation used.
4. Discussion
The aim of the study was to describe how three scenarios of protein levels elicited variations in the growth of laying hens and how such changes might affect long-term egg production and egg components. To our knowledge, this is the first study to investigate the influence of dietary balanced protein in laying hens, from the rearing period (eight w-old) until the end of laying cycle (102 w-old). Currently, there is a growing concept to keep laying hens for longer periods in production [
1]. However, maintaining the egg production and egg quality of a flock of older hens is a challenge. Pieces of evidence demonstrate that body weight of laying hens at the onset of lay may affect the entire egg production [
26] and the egg weight [
27,
28]. Specifically, the body composition of laying hens at the beginning of the laying phase could also affect the peak and persistence of egg production [
29]. The dietary protein content is known to affect the growth of broilers and breeder pullets [
30,
31], and considering the higher cost of dietary protein [
32] and the trend in reducing the nitrogen excretion in poultry farms [
17], it might be convenient to investigate the effects of dietary protein over the growing and laying phase. Assuming that essential amino acids are required in constant ratios with lysine, in this study the concept of balanced protein was used as proposed by Eits et al. [
33].
We observed that dietary balanced protein levels used in this study, poorly affected the daily feed intake of laying hens. A general theory for feed intake regulation was developed over the years [
34,
35,
36], suggesting that feed intake is regulated by the first limiting component in the feed, being energy or essential amino acids. Evidence demonstrates that feed intake of growing broiler chicken and pullet of broiler breeder is affected by dietary protein [
30,
31]. For laying birds, the feed intake regulation seems to be more complex because the consumed nutrients are also used for egg production. A model proposed by Fisher et al. [
37] and recently reviewed by Sakomura et al. [
38] accommodated this problem, splitting the amino acid requirement for maintenance and egg mass, which was called the Reading model. Those authors introduced a methodology to predict the requirements of essential amino acids (mg/hen/day), highlighting the importance to understand the mechanisms related to feed intake regulation. In the present study, the higher level of dietary balanced protein elicited an increase in egg mass. According to the cited model, egg mass will affect the requirement of essential amino acids, which may explain why feed intake did not reduce for laying hens in the H group. On the contrary, laying hens in the L group reduced the egg mass, therefore, a lower amino acid was needed for egg production, which may have impacted the feed intake. An interesting behaviour of feed intake was observed after 74 weeks. Laying hens from all groups reduced their intake of feed. For egg production, comparing the values in the peak and that observed at week 74th, egg production reduced by about 2.6, 2.5, and 3.2 units, respectively for hens consuming the standard, low, and high dietary balanced protein feeds, which may be related to a reduction in laying hens needs, consequently, the feed intake.
On the other hand, when the cumulated feed intake was calculated per unit of egg produced, it was evidenced that laying hens in the L group consumed 130 g of feed per unit of egg produced, whereas the S and H groups consumed 122 and 120 g of feed per egg produced. The feed intake per unit of body weight was 50.3, 49.7, and 44.2 kg of feed per kg of body weight for the L, S, and H groups, respectively. Those results suggest that hens attempted to regulate their feed intake to compensate for the reduction in dietary balanced protein when the feed is deficient and reduce the feed intake when the dietary balanced protein is in excess. Recently, Kumar et al. [
39] described a quadratic response of feed intake in function of the dietary balanced protein. Different from the study reported herein, laying hens (Lohmann-LSL) received a standard feed in the rearing phase. The laying hen’s current status, regarding body weight and body composition, seems to be an important factor that modulates their response and needs more attention in future studies.
Even though the reduction in dietary balanced protein may reduce the feed cost and nitrogen excretion [
16,
40], the feed intake per hen-housed egg increased. In this study, the number of eggs produced per hen-housed reduced 31 units for hens in the L group compared with the S group. Therefore, feeding cost (feed price x feed intake), revenue, and viability of hens should be accounted for to properly calculate the economic return. Viability observed in the L group was 85%, whereas for the S and H groups were 90 and 93%, respectively. Laying hens consuming the L feed demonstrated an acute reduction in body fat after 54 w-old. The ovulation cycle was demonstrated to be dependent on plasma-free fatty acids and the body lipid seems to be the main blood source of fatty acids [
41]. We hypothesized that the ovulation cycle was affected by a reduction in body fat content, which reduces egg production in the L group. Eventually, a severe reduction of body fat might drastically affect the ovulation cycle and may stop egg production, reducing the viability of hens in the L group.
Laying hens from the H group had a body weight close to the recommendations in the guideline (Lohmann Tierzucht GmbH, Cuxhaven, Germany), whereas the hens from the S and L groups were 200 g lighter. The variation in body weight was mostly due to body fat, since after sexual maturity there is a reduction in body protein deposition, and the change in body weight is given by variations in body fat deposition [
2,
42]. The observations on body fat and egg production suggest that the hens in the H group did not have an excess of body lipid, as the laying performance was not affected. On the contrary, the persistence of egg production indicates that body fat in the H group was favourable. Milisits et al. [
29] observed that laying hens with high body fat content at the onset of lay reduced the egg production in about 11 to 13 eggs when compared with hens with lower body fat content. There is a discussion about the importance of energy reserves as body fat; however, there is a lack of information on the desired body fat content that benefits longer-term egg production.
Using the first derivative of the linear plus exponential equation, the results demonstrated an increase in body fat content until 48 (L), 58(S), and 63 (H) w-old, followed by a linear reduction until 102 w-old. As cited before, few differences were observed between groups, where laying hens from the H group seem to have a delay in body fat mobilization, regarding the age. After the cited ages, the laying hens mobilize body fat, possible to maintain egg production, especially after 82 w-old, when the dietary metabolizable energy was reduced, as recommended by the guideline. Nonis and Gous [
43] demonstrated that broiler breeders produce energy from body lipid if they are allowed to do it so, even though the concentration of dietary energy is above requirement, by regulating their feed intake. Similarly, Caldas et al. [
44] observed a reduction in broiler breeders fat at the end of egg production phase. In line with our findings, Kumar et al. [
17] observed a linear increase in body fat for laying hens consuming a feed with a crescent level of digestible lysine (ranging from 560 to 858 mg/hen/day). However, the authors investigated the effect of balanced protein in Lohmann-LSL Lite NA only until 66 w-old.
The body ash content of laying hens suggested that laying hens did not use mineral reserves to produce an egg, as the body ash increased until the end of the laying cycle. The major portion of minerals used for egg formation is due to calcium carbonate necessary for eggshell formation given approximately 80% of eggshell is formed by this mineral [
45]. Around 99% of total body calcium is found in the bone ash [
46]. Evidence demonstrates that in a flock of older hens, there are individuals with a tendency to develop osteoporosis [
47], and there are an increased number of eggs with thinner eggshells due to lower ability to uptake calcium and phosphorus from the intestinal lumen [
48]. In this study, dietary balanced protein seems to have a low or no effect over the dynamics of body ash contents. Apparently, the advanced ages of laying hens used in this study was not sufficient to elicit a negative consequence in the bone structure; however, in all treatments, the eggshell percentage reduced with age, probably due to the increase in egg size with the age of laying hens [
49].
The dynamic of egg components observed over time was similar to that reported by Bendezu et al. [
5] for white laying hens from 18 to 60 w-old. As the laying hens aged, the yolk percentage increased, and the albumen and eggshell reduced, which was consistent with other reported literature [
50,
51]. The dietary balanced protein tended to influence yolk percentage. Compared with the L group, the hens consuming the H feed produced eggs with more percentage of yolk. The contribution of dietary protein to yolk formation is probably related to phosvitin as this is the major protein molecule found in egg yolk. Around 56% of the amino acid found in the phosvitin is serine phosphorylated [
52,
53]. According to Huang et al. [
54], it is believed that the role of phosvitin in the egg is related to embryo development, which reinforces the importance of such a constituent in egg yolk. In the present study, the tendency of a lower percentage of yolk observed in eggs of the L group may be related to the lower amount of dietary serine, which is necessary to produce the phosvitin in the egg. With lower sources of dietary serine, essential amino acids might be used to overcome this deficiency.
Among the responses observed in this study, the body fat content between laying hens consuming the different levels of balanced protein was unexpected. Most reports in the literature demonstrate that growing birds would increase body lipid content when they are offered a low balanced protein feed [
30,
31]. The opposite result is reported when a growing bird consumes a high balanced protein feed. The regulation of body fat content in laying birds seems to be more complex and the prediction of body fat in laying hens should be done with caution. The increase in egg yolk percentage may contribute to a higher value of body fat content as 34% of egg yolk is constituted of lipid [
55]. Hocking [
56] investigated the effect of body weight and feed intake on the ovarium follicular dynamics and found that feed-restricted broiler breeders reduced the number of yellow follicles. We did not find a similar study for laying hens, which would contribute to a better understanding of the results observed, but we hypothesize that a pullet raised with high level of dietary balanced protein feed may increase the number and the weight of yellow follicles in the ovarium, resulting in more body fat content in laying hens.


 ,
, 



{kind=link}
{kind=link}
{kind=link}