
Potential Probiotic Acceptability of a Novel Strain of Paenibacillus konkukensis SK 3146 and Its Dietary Effects on Growth Performance, Intestinal Microbiota, and Meat Quality in Broilers

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. In Vitro Experiments of Probiotic Acceptability
2.1.1. Hemolytic Activity
2.1.2. Antibiotic Susceptibility
2.1.3. Acid and Bile Salt Tolerance
2.1.4. Adhesion Assay
2.2. Broiler Experiment
2.2.1. Preparation of P. konkukensis Bacterial Culture
2.2.2. Experimental Animals, Diets, and Design
2.2.3. Growth Performance
2.2.4. Sample Collection and Measurements
2.3. Statistical Analysis
3. Results
3.1. Hemolytic Activity and Antibiotic Susceptibility
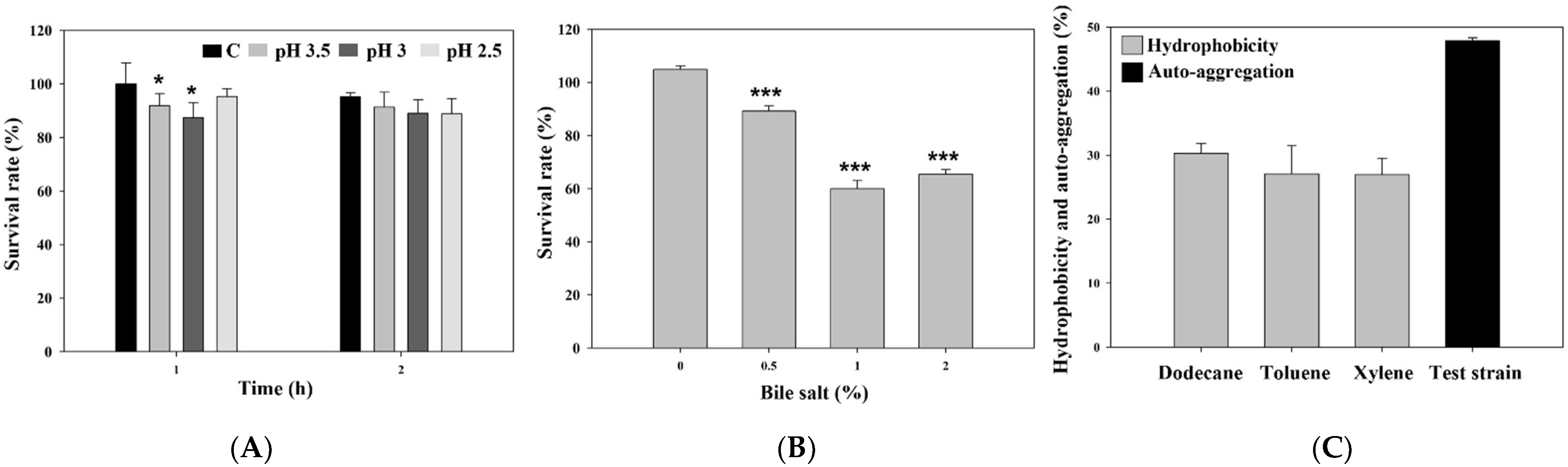
3.2. Acid and Bile Salt Tolerance and Adhesion Properties
3.3. Growth Performance
3.4. Intestinal Weight and Length
3.5. Intestinal Microbiota
3.6. Meat Quality
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moore, P.; Evenson, A.; Luckey, T.; McCoy, E.; Elvehjem, C.; Hart, E. Use of sulfasuxidine, streptothricin, and streptomycin in nutritional studies with the chick. J. Biol. Chem. 1946, 165, 437–441. [Google Scholar] [CrossRef]
- Khalid, F.; Khalid, A.; Fu, Y.; Hu, Q.; Zheng, Y.; Khan, S.; Wang, Z. Potential of Bacillus velezensis as a probiotic in animal feed: A review. J. Microbiol. 2021, 59, 627–633. [Google Scholar] [CrossRef] [PubMed]
- Gadde, U.; Kim, W.; Oh, S.; Lillehoj, H.S. Alternatives to antibiotics for maximizing growth performance and feed efficiency in poultry: A review. Anim. Health Res. Rev. 2017, 18, 26–45. [Google Scholar] [CrossRef] [PubMed]
- Suresh, G.; Das, R.K.; Kaur Brar, S.; Rouissi, T.; Avalos Ramirez, A.; Chorfi, Y.; Godbout, S. Alternatives to antibiotics in poultry feed: Molecular perspectives. Crit. Rev. Microbiol. 2018, 44, 318–335. [Google Scholar] [CrossRef] [PubMed]
- Moon, S.-G.; Lee, S.-K.; Lee, W.-D.; Niu, K.-M.; Hwang, W.-U.; Oh, J.-S.; Kothari, D.; Kim, S.-K. Effect of dietary supplementation of a phytogenic blend containing Schisandra chinensis, Pinus densiflora, and Allium tuberosum on productivity, egg quality, and health parameters in laying hens. Anim. Biosci. 2021, 34, 285. [Google Scholar] [CrossRef]
- Moon, S.-G.; Kothari, D.; Kim, W.-L.; Lee, W.-D.; Kim, K.-I.; Kim, J.-I.; Kim, E.-J.; Kim, S.-K. Feasibility of sodium long chain polyphosphate as a potential growth promoter in broilers. J. Anim. Sci. Technol. 2021, 63, 1286–1300. [Google Scholar] [CrossRef]
- Alagawany, M.; El-Hack, A.; Mohamed, E.; Farag, M.R.; Sachan, S.; Karthik, K.; Dhama, K. The use of probiotics as eco-friendly alternatives for antibiotics in poultry nutrition. Environ. Sci. Pollut. Res. 2018, 25, 10611–10618. [Google Scholar] [CrossRef]
- Zhan, H.; Dong, X.; Li, L.; Zheng, Y.; Gong, Y.; Zou, X. Effects of dietary supplementation with Clostridium butyricum on laying performance, egg quality, serum parameters, and cecal microflora of laying hens in the late phase of production. Poult. Sci. 2019, 98, 896–903. [Google Scholar] [CrossRef]
- Hill, C.; Guarner, F.; Reid, G.; Gibson, G.R.; Merenstein, D.J.; Pot, B.; Morelli, L.; Canani, R.B.; Flint, H.J.; Salminen, S. Expert consensus document: The International Scientific Association for Probiotics and Prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 2014, 11, 506–514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Attia, Y.; Ellakany, H.; El-Hamid, A.; Bovera, F.; Ghazaly, S. Control of Salmonella enteritidis infection in male layer chickens by acetic acid and/or prebiotics, probiotics and antibiotics. Arch. Geflügelk 2012, 76, 239–245. [Google Scholar]
- Attia, Y.; Abd El Hamid, E.; Ismaiel, A.; El-Nagar, A. The detoxication of nitrate by two antioxidants or a probiotic, and the effects on blood and seminal plasma profiles and reproductive function of New Zealand White rabbit bucks. Animal 2013, 7, 591–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adel, M.; El-Sayed, A.-F.M.; Yeganeh, S.; Dadar, M.; Giri, S.S. Effect of potential probiotic Lactococcus lactis subsp. lactis on growth performance, intestinal microbiota, digestive enzyme activities, and disease resistance of Litopenaeus vannamei. Probiotics Antimicrob. Proteins 2017, 9, 150–156. [Google Scholar] [CrossRef]
- de Souza, L.F.; Araújo, D.N.; Stefani, L.M.; Giometti, I.C.; Cruz-Polycarpo, V.C.; Polycarpo, G.; Burbarelli, M.F. Probiotics on performance, intestinal morphology and carcass characteristics of broiler chickens raised with lower or higher environmental challenge. Austral. J. Vet. Sci. 2018, 50, 35–41. [Google Scholar] [CrossRef] [Green Version]
- Salah, R.B.; Trabelsi, I.; Mansour, R.B.; Lassoued, S.; Chouayekh, H.; Bejar, S. A new Lactobacillus plantarum strain, TN8, from the gastro intestinal tract of poultry induces high cytokine production. Anaerobe 2012, 18, 436–444. [Google Scholar] [CrossRef]
- Dec, M.; Puchalski, A.; Urban-Chmiel, R.; Wernicki, A. Screening of Lactobacillus strains of domestic goose origin against bacterial poultry pathogens for use as probiotics. Poult. Sci. 2014, 93, 2464–2472. [Google Scholar] [CrossRef]
- Grady, E.N.; MacDonald, J.; Liu, L.; Richman, A.; Yuan, Z.-C. Current knowledge and perspectives of Paenibacillus: A review. Microb. Cell Factories 2016, 15, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Calik, A.; Burcu, E.; Bayraktaroğlu, A.G.; Ergün, A.; Saçakli, P. Effects of dietary probiotic and synbiotic supplementation on broiler growth performance and intestinal histomorphology. Ank. Üniversitesi Vet. Fakültesi Derg. 2017, 64, 183–189. [Google Scholar]
- Ekim, B.; Calik, A.; Ceylan, A.; Saçaklı, P. Effects of Paenibacillus xylanexedens on growth performance, intestinal histomorphology, intestinal microflora, and immune response in broiler chickens challenged with Escherichia coli K88. Poult. Sci. 2020, 99, 214–223. [Google Scholar] [CrossRef]
- Wu, Y.; Wang, B.; Zeng, Z.; Liu, R.; Tang, L.; Gong, L.; Li, W. Effects of probiotics Lactobacillus plantarum 16 and Paenibacillus polymyxa 10 on intestinal barrier function, antioxidative capacity, apoptosis, immune response, and biochemical parameters in broilers. Poult. Sci. 2019, 98, 5028–5039. [Google Scholar] [CrossRef]
- Alagawany, M.; Madkour, M.; El-Saadony, M.T.; Reda, F.M. Paenibacillus polymyxa (LM31) as a new feed additive: Antioxidant and antimicrobial activity and its effects on growth, blood biochemistry, and intestinal bacterial populations of growing Japanese quail. Anim. Feed. Sci. Technol. 2021, 276, 114920. [Google Scholar] [CrossRef]
- Wang, B.; Gong, L.; Zhou, Y.; Tang, L.; Zeng, Z.; Wang, Q.; Zou, P.; Yu, D.; Li, W. Probiotic Paenibacillus polymyxa 10 and Lactobacillus plantarum 16 enhance growth performance of broilers by improving the intestinal health. Anim. Nutr. 2021, 7, 829–840. [Google Scholar] [CrossRef]
- Alshelmani, M.; Loh, T.; Foo, H.; Sazili, A.; Lau, W. Effect of feeding different levels of palm kernel cake fermented by Paenibacillus polymyxa ATCC 842 on broiler growth performance, blood biochemistry, carcass characteristics, and meat quality. Anim. Prod. Sci. 2016, 57, 839–848. [Google Scholar] [CrossRef]
- Im, W.-T.; Yi, K.-J.; Lee, S.-S.; Moon, H.I.; Jeon, C.O.; Kim, D.-W.; Kim, S.-K. Paenibacillus konkukensis sp. nov., isolated from animal feed. Int. J. Syst. Evol. Microbiol. 2017, 67, 2343–2348. [Google Scholar] [CrossRef]
- Jung, H.-I.; Park, S.; Niu, K.-M.; Lee, S.-W.; Kothari, D.; Yi, K.J.; Kim, S.-K. Complete genome sequence of Paenibacillus konkukensis sp. nov. SK3146 as a potential probiotic strain. J. Anim. Sci. Technol. 2021, 63, 666–670. [Google Scholar] [CrossRef] [PubMed]
- Das, D.; Goyal, A. Potential probiotic attributes and antagonistic activity of an indigenous isolate Lactobacillus plantarum DM5 from an ethnic fermented beverage “Marcha” of North Eastern Himalayas. Int. J. Food Sci. Nutr. 2014, 65, 335–344. [Google Scholar] [CrossRef] [PubMed]
- Kwon, H.; Choo, Y.; Choi, Y.; Kim, E.; Kim, H.; Heo, K.; Choi, H.; Lee, S.; Kim, C.; Kim, B. Carcass characteristics and meat quality of Korean native ducks and commercial meat-type ducks raised under same feeding and rearing conditions. Asian Australas. J. Anim. Sci. 2014, 27, 1638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dave, R.; Shah, N. Evaluation of media for selective enumeration of Streptococcus thermophilus, Lactobacillus delbrueckii ssp. Bulgaricus, Lactobacillus acidophilus, and Bifidobacteria. J. Dairy Sci. 1996, 79, 1529–1536. [Google Scholar]
- Alshammari, E.; Patel, M.; Sachidanandan, M.; Kumar, P.; Adnan, M. Potential evaluation and health fostering intrinsic traits of novel probiotic strain Enterococcus durans F3 isolated from the gut of fresh water fish catla catla. Food Sci. Anim. Resour. 2019, 39, 844. [Google Scholar] [CrossRef] [Green Version]
- Giri, S.S.; Sen, S.S.; Saha, S.; Sukumaran, V.; Park, S.C. Use of a potential probiotic, Lactobacillus plantarum L7, for the preparation of a rice-based fermented beverage. Front. Microbiol. 2018, 9, 473. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.D. Diverse origins of tetracycline resistance in the honey bee bacterial pathogen Paenibacillus larvae. J. Invertebr. Pathol. 2003, 83, 46–50. [Google Scholar] [CrossRef]
- Pednekar, P.B.; Jain, R.; Thakur, N.L.; Mahajan, G.B. Isolation of multi-drug resistant Paenibacillus sp. from fertile soil: An imminent menace of spreading resistance. J. Life Sci. 2010, 4, 7–12. [Google Scholar]
- Musikasang, H.; Tani, A.; H-kittikun, A.; Maneerat, S. Probiotic potential of lactic acid bacteria isolated from chicken gastrointestinal digestive tract. World J. Microbiol. Biotechnol. 2009, 25, 1337–1345. [Google Scholar] [CrossRef]
- Shi, Y.; Zhai, M.; Li, J.; Li, B. Evaluation of safety and probiotic properties of a strain of Enterococcus faecium isolated from chicken bile. J. Food Sci. Technol. 2020, 57, 578–587. [Google Scholar] [CrossRef]
- Lin, J.; Sahin, O.; Michel, L.O.; Zhang, Q. Critical role of multidrug efflux pump CmeABC in bile resistance and in vivo colonization of Campylobacter jejuni. Infect. Immun. 2003, 71, 4250–4259. [Google Scholar] [CrossRef] [Green Version]
- García-Hernández, Y.; Pérez-Sánchez, T.; Boucourt, R.; Balcázar, J.L.; Nicoli, J.R.; Moreira-Silva, J.; Rodríguez, Z.; Fuertes, H.; Nuñez, O.; Albelo, N. Isolation, characterization and evaluation of probiotic lactic acid bacteria for potential use in animal production. Res. Vet. Sci. 2016, 108, 125–132. [Google Scholar] [CrossRef]
- Reuben, R.C.; Roy, P.C.; Sarkar, S.L.; Alam, R.-U.; Jahid, I.K. Isolation, characterization, and assessment of lactic acid bacteria toward their selection as poultry probiotics. BMC Microbiol. 2019, 19, 253–272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cotter, P.D.; Hill, C. Surviving the acid test: Responses of gram-positive bacteria to low pH. Microbiol. Mol. Biol. Rev. 2003, 67, 429–453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ashraf, R.; Smith, S. Commercial lactic acid bacteria and probiotic strains-tolerance to bile, pepsin and antibiotics. Int. Food Res. J. 2016, 23, 777. [Google Scholar]
- Alshelmani, M.; Loh, T.; Foo, H.; Sazili, A.; Lau, W. Effect of feeding different levels of palm kernel cake fermented by Paenibacillus polymyxa ATCC 842 on nutrient digestibility, intestinal morphology, and gut microflora in broiler chickens. Anim. Feed. Sci. Technol. 2016, 216, 216–224. [Google Scholar] [CrossRef] [Green Version]
- Baldwin, S.; Hughes, R.J.; Hao Van, T.T.; Moore, R.J.; Stanley, D. At-hatch administration of probiotic to chickens can introduce beneficial changes in gut microbiota. PLoS ONE 2018, 13, e0194825. [Google Scholar] [CrossRef] [PubMed]
- Marume, U.; Mokagane, J.; Shole, C.; Hugo, A. Citrullus lanatus essential oils inclusion in diets elicit nutraceutical effects on egg production, egg quality, and physiological characteristics in layer hens. Poult. Sci. 2020, 99, 3038–3046. [Google Scholar] [CrossRef] [PubMed]
- Reis, M.; Fassani, E.; Júnior, A.G.; Rodrigues, P.; Bertechini, A.; Barrett, N.; Persia, M.; Schmidt, C. Effect of Bacillus subtilis (DSM 17299) on performance, digestibility, intestine morphology, and pH in broiler chickens. J. Appl. Poult. Res. 2017, 26, 573–583. [Google Scholar] [CrossRef]
- Molnár, A.; Podmaniczky, B.; Kürti, P.; Tenk, I.; Glávits, R.; Virág, G.; Szabó, Z. Effect of different concentrations of Bacillus subtilis on growth performance, carcase quality, gut microflora and immune response of broiler chickens. Br. Poult. Sci. 2011, 52, 658–665. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ni, X.; Qing, X.; Zeng, D.; Luo, M.; Liu, L.; Li, G.; Pan, K.; Jing, B. Live probiotic Lactobacillus johnsonii BS15 promotes growth performance and lowers fat deposition by improving lipid metabolism, intestinal development, and gut microflora in broilers. Front. Microbiol. 2017, 8, 1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancini, R.; Hunt, M. Current research in meat color. Meat Sci. 2005, 71, 100–121. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Dong, X.; Yao, Z.; Xu, B.; Zhen, S.; Li, C.; Li, X. Effects of prechilling parameters on water-holding capacity of chilled pork and optimization af prechilling parameters using response surface methodology. J. Anim. Sci. 2012, 90, 2836–2841. [Google Scholar] [CrossRef]
- Al-Shawi, S.G.; Dang, D.S.; Yousif, A.Y.; Al-Younis, Z.K.; Najm, T.A.; Matarneh, S.K. The potential use of probiotics to improve animal health, efficiency, and meat quality: A Review. Agriculture 2020, 10, 452. [Google Scholar] [CrossRef]
- Hossain, M.E.; Kim, G.M.; Lee, S.K.; Yang, C.J. Growth performance, meat yield, oxidative stability, and fatty acid composition of meat from broilers fed diets supplemented with a medicinal plant and probiotics. Asian-Australas. J. Anim. Sci. 2012, 25, 1159. [Google Scholar] [CrossRef]
- Hussein, E.; Selim, S. Efficacy of yeast and multi-strain probiotic alone or in combination on growth performance, carcass traits, blood biochemical constituents, and meat quality of broiler chickens. Livest. Sci. 2018, 216, 153–159. [Google Scholar] [CrossRef]
- Novak, R.; Bogovič Matijašić, B.; Terčič, D.; Červek, M.; Gorjanc, G.; Holcman, A.; Levart, A.; Rogelj, I. Effects of two probiotic additives containing Bacillus spores on carcass characteristics, blood lipids and cecal volatile fatty acids in meat type chickens. J. Anim. Physiol. Anim. Nutr. 2011, 95, 424–433. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Item | Grower Phase (Day 1 to 21) | Finisher Phase (Day 21 to 35) |
|---|---|---|
| Ingredient composition, % | ||
| Corn | 57.2 | 52.53 |
| Soybean meal (crude protein, 45%) | 27.09 | 26.48 |
| Rice bran | - | 3 |
| Corn gluten meal | 3.38 | 3 |
| Corn dried distillers’ grains with solubles (DDGS) | 3 | 3 |
| Tallow | 3 | 5 |
| Rapeseed meal | 2 | 2 |
| Wheat bran | - | 1.25 |
| Limestone | 1.69 | 1.79 |
| Monocalcium phosphate (MCP) | 1.16 | 0.91 |
| L-lysine HCl 78% | 0.38 | 0.19 |
| DL-methionine 98% | 0.36 | 0.17 |
| Salt | 0.25 | 0.25 |
| Choline-Cl 50% | 0.09 | 0.04 |
| Mineral premix (1) | 0.15 | 0.15 |
| Vitamin premix (2) | 0.12 | 0.11 |
| Phytase optis | 0.05 | 0.05 |
| NaHCO3 | 0.08 | 0.08 |
| Calculated analysis | ||
| Crude protein, % | 20.5 | 20.0 |
| Crude fat, % | 5.84 | 8.23 |
| Crude fiber, % | 2.8 | 2.92 |
| Ash, % | 4.74 | 4.97 |
| Ca, % | 0.80 | 0.90 |
| Available P, % | 0.62 | 0.61 |
| Met+Cys, % | 1.03 | 0.83 |
| AMEn (kcal/kg) | 3100 | 3150 |
| Item | Diameter (mm) | Sensitivity |
|---|---|---|
| Gentamycin | 13 | S |
| Vancomycin | 34 | S |
| Ampicillin | 14 | S |
| Chloramphenicol | 40 | S |
| Clindamycin | 40 | S |
| Erythromycin | 40 | S |
| Oxacillin | ND | R |
| Tetracycline | 20 | S |
| Kanamycin | 15 | S |
| Item | Treatment | p-Value | ||
|---|---|---|---|---|
| NC | PC | PK | ||
| BW (g) | ||||
| Day 7 (initial weight) | 134.46 ± 0.04 | 134.41 ± 0.06 | 134.49 ± 0.03 | 0.39 |
| Day 21 | 664.25 ± 19.61 | 704.44 ± 26.99 | 685.89 ± 18.37 | 0.46 |
| Day 35 | 1536.63 ± 56.16 | 1618.06 ± 54.29 | 1573.13 ± 60.85 | 0.62 |
| BW gain (g/day/bird) | ||||
| Grower phase | 529.79 ± 19.59 | 570.03 ± 26.95 | 551.40 ± 18.37 | 0.46 |
| Finisher phase | 872.38 ± 39.18 | 913.63 ± 31.12 | 887.24 ± 44.22 | 0.75 |
| Total phase | 1402.16 ± 56.114 | 1483.66 ± 54.27 | 1438.64 ± 60.85 | 0.62 |
| ADG (g/day/bird) | ||||
| Grower phase | 37.84 ± 1.40 | 40.72 ± 1.93 | 39.39 ± 1.31 | 0.46 |
| Finisher phase | 62.31 ± 2.80 | 65.26 ± 2.22 | 63.37 ± 3.16 | 0.75 |
| Total phase | 50.08 ± 2.dd00 | 52.99 ± 1.94 | 51.38 ± 2.17 | 0.62 |
| ADFI (g/day/bird) | ||||
| Grower phase | 63.13 ± 1.10 | 62.75 ± 2.64 | 64.58 ± 3.07 | 0.85 |
| Finisher phase | 125.28 ± 2.97 | 129.94 ± 3.10 | 128.35 ± 4.09 | 0.63 |
| Total phase | 94.20 ± 1.69 | 96.34 ± 2.52 | 96.47 ± 3.57 | 0.81 |
| FCR | ||||
| Grower phase | 1.67 ± 0.04 | 1.54 ± 0.03 | 1.65 ± 0.12 | 0.45 |
| Finisher phase | 2.02 ± 0.05 | 1.99 ± 0.02 | 2.04 ± 0.13 | 0.91 |
| Total phase | 1.89 ± 0.04 | 1.82 ± 0.02 | 1.89 ± 0.12 | 0.77 |
| Item | Treatment | p-Value | ||
|---|---|---|---|---|
| NC | PC | PK | ||
| Relative weight, g/100 g BW | ||||
| Duodenum | 0.58 ab ± 0.04 | 0.45 b ± 0.03 | 0.65 a ± 0.04 | <0.01 |
| Jejunum | 0.95 ab ± 0.07 | 0.71 b ± 0.03 | 1.04 a ± 0.09 | 0.01 |
| Ileum | 0.92 ab ± 0.08 | 0.70 b ± 0.04 | 0.94 a ± 0.08 | 0.03 |
| Cecum | 0.19 ± 0.04 | 0.17 ± 0.01 | 0.16 ± 0.02 | 0.64 |
| Relative length, cm/100 g BW | ||||
| Duodenum | 2.10 a ± 0.10 | 1.78 b ± 0.05 | 1.83 b ± 0.06 | 0.01 |
| Jejunum | 5.03 a ± 0.25 | 4.13 b ± 0.18 | 4.16 b ± 0.20 | 0.01 |
| Ileum | 5.57 a ± 0.24 | 4.55 b ± 0.21 | 4.55 b ± 0.24 | <0.01 |
| Cecum | 1.13 ± 0.04 | 1.37 ± 0.26 | 1.07 ± 0.06 | 0.36 |
| Weight/length, g/cm | ||||
| Duodenum | 0.28 ab ± 0.03 | 0.25 b ± 0.02 | 0.35 a ± 0.02 | 0.02 |
| Jejunum | 0.19 b ± 0.02 | 0.17 b ± 0.01 | 0.25 a ± 0.02 | <0.01 |
| Ileum | 0.17 ab ± 0.02 | 0.16 b ± 0.01 | 0.21 a ± 0.02 | 0.05 |
| Cecum | 0.17 ± 0.03 | 0.14 ± 0.02 | 0.15 ± 0.01 | 0.61 |
| Type | Treatment (Log CFU/g) | p-Value | ||
|---|---|---|---|---|
| NC | PC | PK | ||
| Jejunum | ||||
| Coliform and lactose-negative enterobacteria | 6.52 b ± 0.27 | 7.36 a ± 0.17 | 7.16 ab ± 0.19 | 0.04 |
| Lactobacilli | 8.41 a ± 0.13 | 7.60 b ± 0.23 | 8.86 a ± 0.12 | <0.01 |
| Total aerobes | 8.05 a ± 0.15 | 6.87 b ± 0.23 | 7.77 a ± 0.18 | <0.01 |
| ST | 7.03 b ± 0.31 | 6.81 b ± 0.16 | 8.41 a ± 0.18 | <0.01 |
| Ileum | ||||
| Coliform and lactose-negative enterobacteria | 6.70 ± 0.34 | 7.58 ± 0.41 | 7.22 ± 0.20 | 0.20 |
| Lactobacilli | 8.95 ± 0.13 | 8.65 ± 0.20 | 8.87 ± 0.12 | 0.40 |
| Total aerobes | 7.51 ± 0.31 | 8.32 ± 0.23 | 8.38 ± 0.20 | 0.05 |
| ST | 7.88 ± 0.34 | 8.26 ± 0.11 | 8.30 ± 0.15 | 0.37 |
| Cecum | ||||
| Coliform and lactose-negative enterobacteria | 7.60 ± 0.21 | 7.56 ± 0.38 | 6.94 ± 0.16 | 0.13 |
| Lactobacilli | 9.27 a ± 0.10 | 8.50 b ± 0.31 | 9.20 ab ± 0.16 | 0.02 |
| Total aerobes | 7.80 ± 0.23 | 8.16 ± 0.35 | 7.65 ± 0.19 | 0.35 |
| ST | 9.00 ± 0.24 | 8.57 ± 0.34 | 9.08 ± 0.23 | 0.33 |
| Type | Treatment (Log CFU/g) | p-Value | |||
|---|---|---|---|---|---|
| NC | PC | PK | |||
| Cooking loss, % | |||||
| Breast | 68.00 ± 1.48 | 69.31 ± 1.10 | 70.11 ± 1.18 | 0.50 | |
| Thigh | 68.57 ± 1.04 | 68.83 ± 1.01 | 70.68 ± 1.59 | 0.44 | |
| Meat color | |||||
| Breast | L* | 57.75 ± 1.06 | 56.26 ± 1.77 | 59.44 ± 1.39 | 0.31 |
| a* | 2.58 ± 0.38 | 2.48 ± 0.40 | 2.28 ± 0.54 | 0.89 | |
| b* | 12.17 ± 0.51 | 13.31 ± 0.53 | 13.59 ± 0.68 | 0.21 | |
| Thigh | L* | 55.14 ± 0.61 | 56.52 ± 0.64 | 56.72 ± 0.34 | 0.11 |
| a* | 14.46 a ± 0.28 | 12.07 b ± 0.45 | 13.60 a ± 0.41 | <0.01 | |
| b* | 6.30 ± 0.43 | 7.43 ± 0.33 | 6.19 ± 0.35 | 0.06 | |
| pH | |||||
| Breast | 5.59 ± 0.04 | 5.66 ± 0.03 | 5.54 ± 0.05 | 0.16 | |
| Thigh | 6.10 ± 0.10 | 6.26 ± 0.04 | 6.15 ± 0.04 | 0.20 | |
| Meat weight, g/100 g BW | |||||
| Breast | 6.43 b ± 0.13 | 6.51 b ± 0.09 | 7.37 a ± 0.27 | <0.01 | |
| Thigh | 6.43 ± 0.13 | 6.51 ± 0.09 | 6.60 ± 0.15 | 0.62 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Moon, S.-G.; Kothari, D.; Lee, W.-D.; Kim, J.-I.; Kim, K.-I.; Kim, Y.-G.; Ga, G.-W.; Kim, E.-J.; Kim, S.-K. Potential Probiotic Acceptability of a Novel Strain of Paenibacillus konkukensis SK 3146 and Its Dietary Effects on Growth Performance, Intestinal Microbiota, and Meat Quality in Broilers. Animals 2022, 12, 1471. https://doi.org/10.3390/ani12111471
Moon S-G, Kothari D, Lee W-D, Kim J-I, Kim K-I, Kim Y-G, Ga G-W, Kim E-J, Kim S-K. Potential Probiotic Acceptability of a Novel Strain of Paenibacillus konkukensis SK 3146 and Its Dietary Effects on Growth Performance, Intestinal Microbiota, and Meat Quality in Broilers. Animals. 2022; 12(11):1471. https://doi.org/10.3390/ani12111471
Chicago/Turabian StyleMoon, Seung-Gyu, Damini Kothari, Woo-Do Lee, Jong-Il Kim, Kyung-Il Kim, Yong-Gi Kim, Gun-Whi Ga, Eun-Jip Kim, and Soo-Ki Kim. 2022. "Potential Probiotic Acceptability of a Novel Strain of Paenibacillus konkukensis SK 3146 and Its Dietary Effects on Growth Performance, Intestinal Microbiota, and Meat Quality in Broilers" Animals 12, no. 11: 1471. https://doi.org/10.3390/ani12111471
APA StyleMoon, S. -G., Kothari, D., Lee, W. -D., Kim, J. -I., Kim, K. -I., Kim, Y. -G., Ga, G. -W., Kim, E. -J., & Kim, S. -K. (2022). Potential Probiotic Acceptability of a Novel Strain of Paenibacillus konkukensis SK 3146 and Its Dietary Effects on Growth Performance, Intestinal Microbiota, and Meat Quality in Broilers. Animals, 12(11), 1471. https://doi.org/10.3390/ani12111471