
Tissue Distribution of the Piscine Novirhabdovirus Genotype IVb in Muskellunge (Esox masquinongy)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Culture and Virus Propagation and Quantitation
2.2. Fish and Maintenance
2.3. Experimental Infection of Juvenile Muskellunge
2.4. Tissue Sampling
2.5. Viral Plaque Assay
2.6. VHSV Reisolation and Confirmation
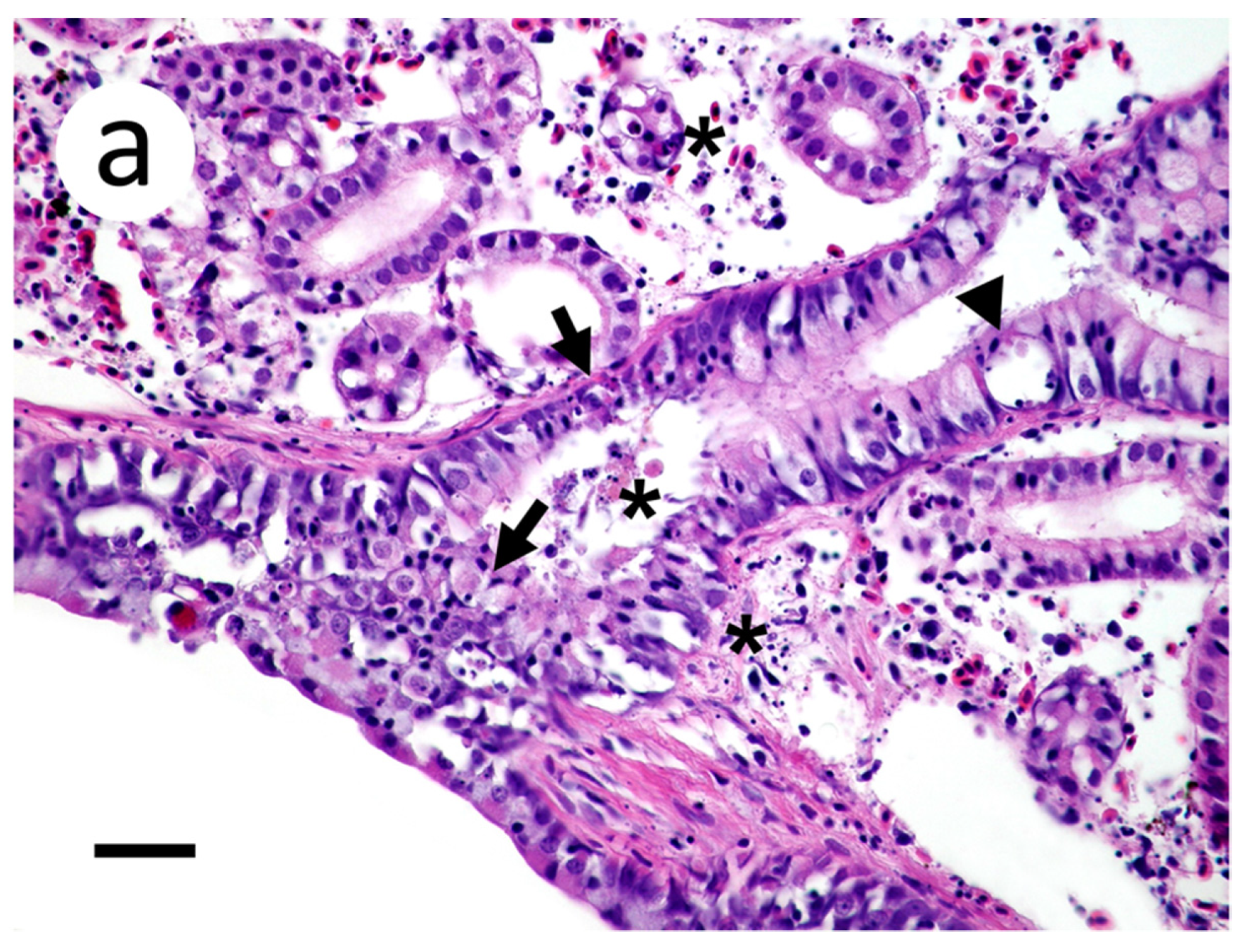
2.7. Histopathology
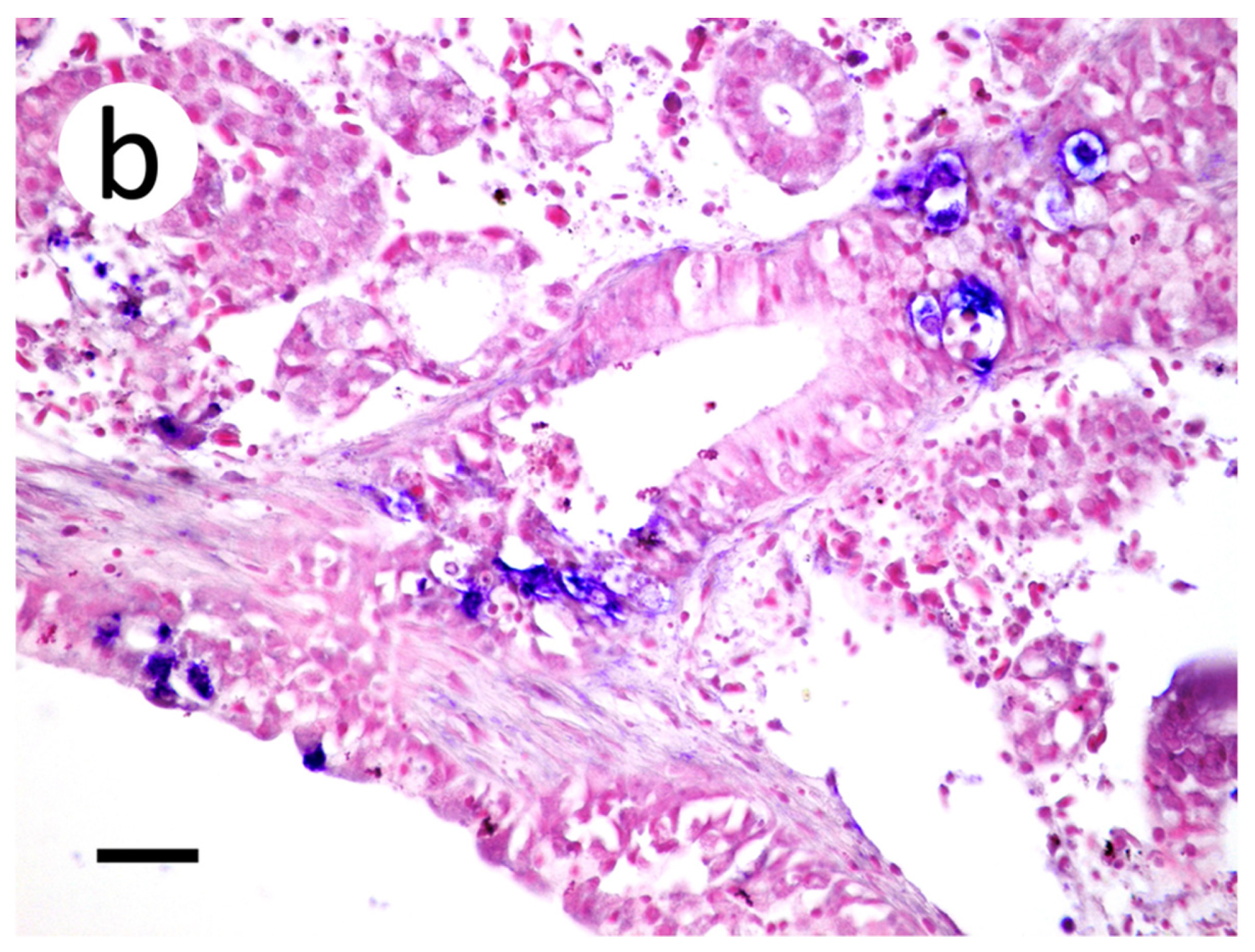
2.8. In Situ Hybridization
2.9. Statistics
3. Results
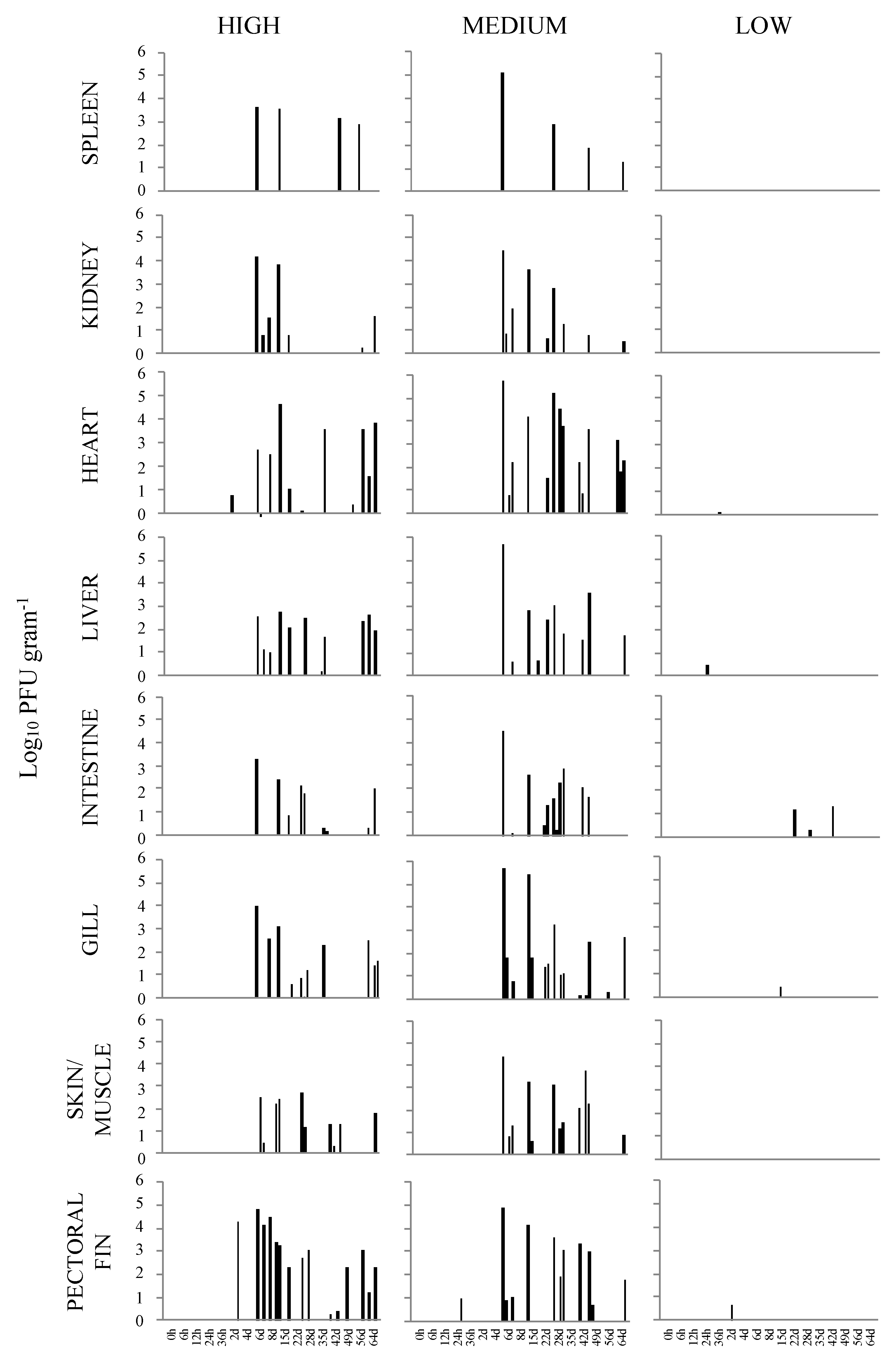
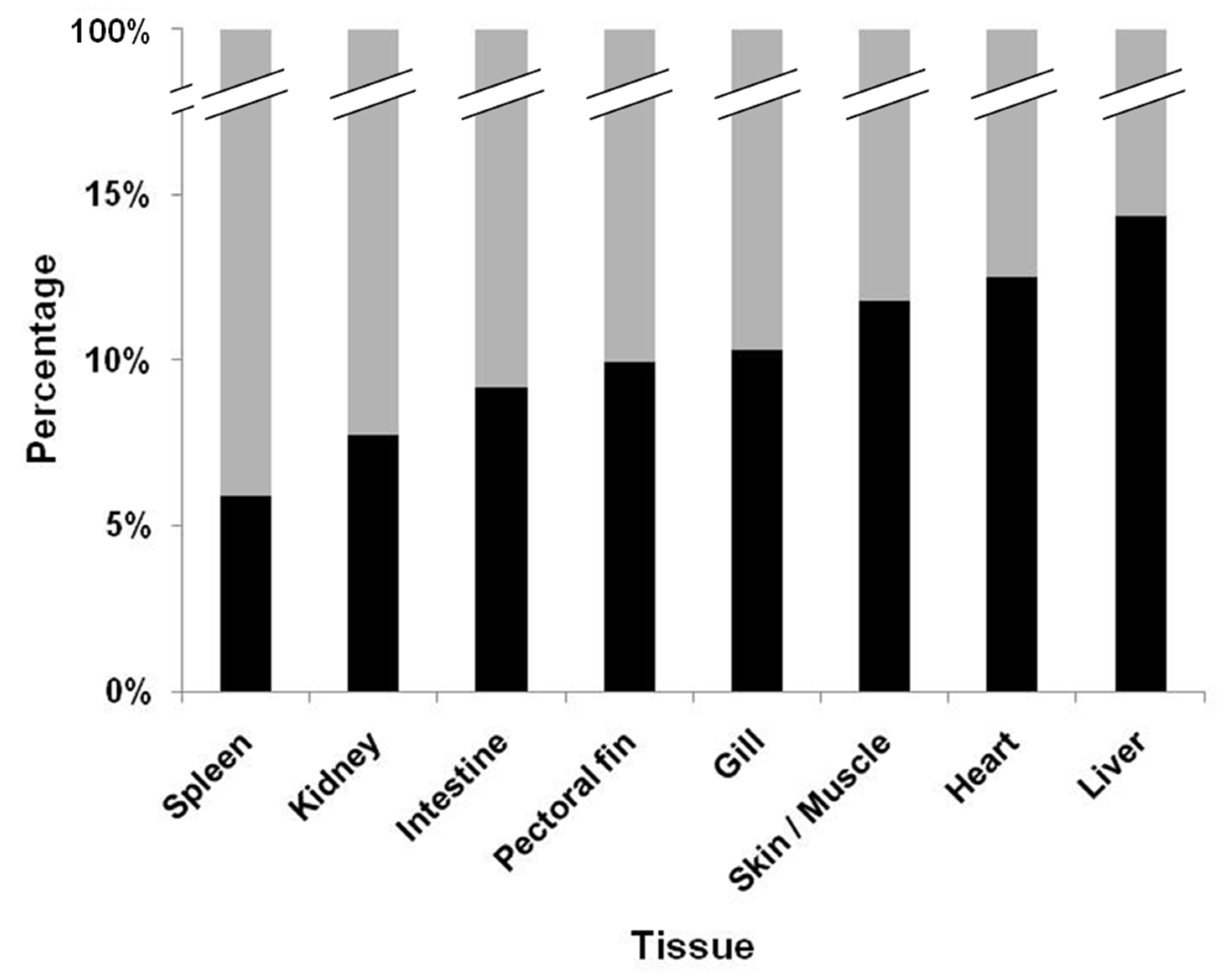
3.1. Virus Reisolation and Infectious Virus Load
3.2. Statistical Analysis
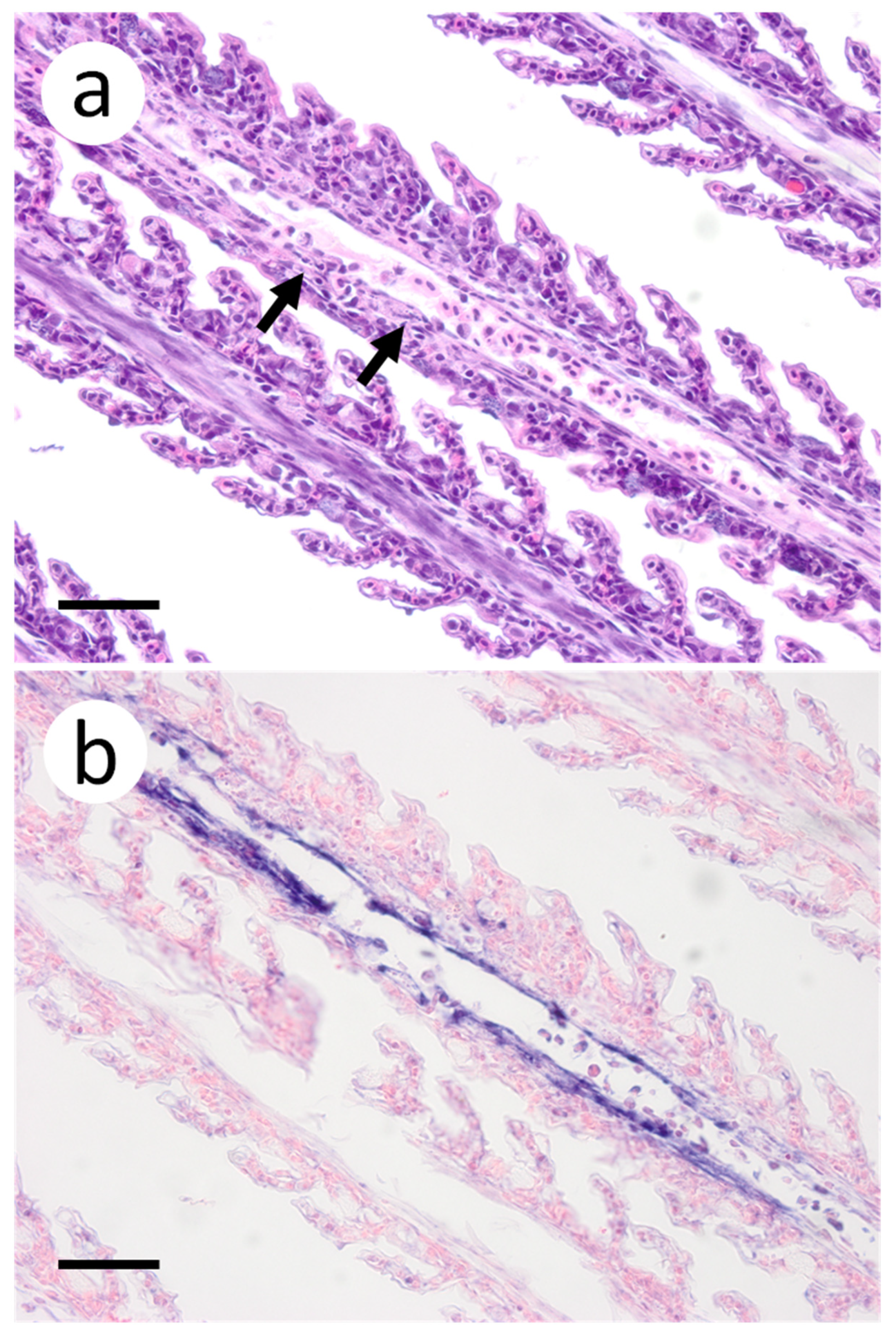
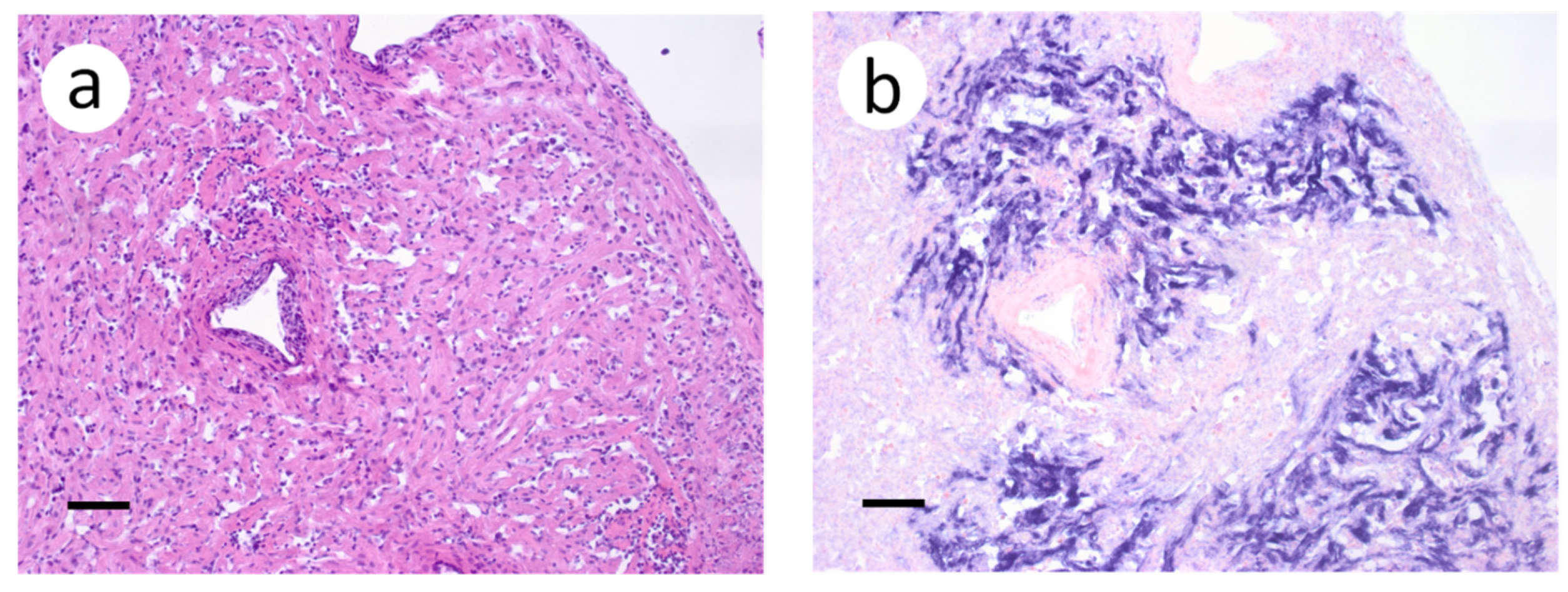
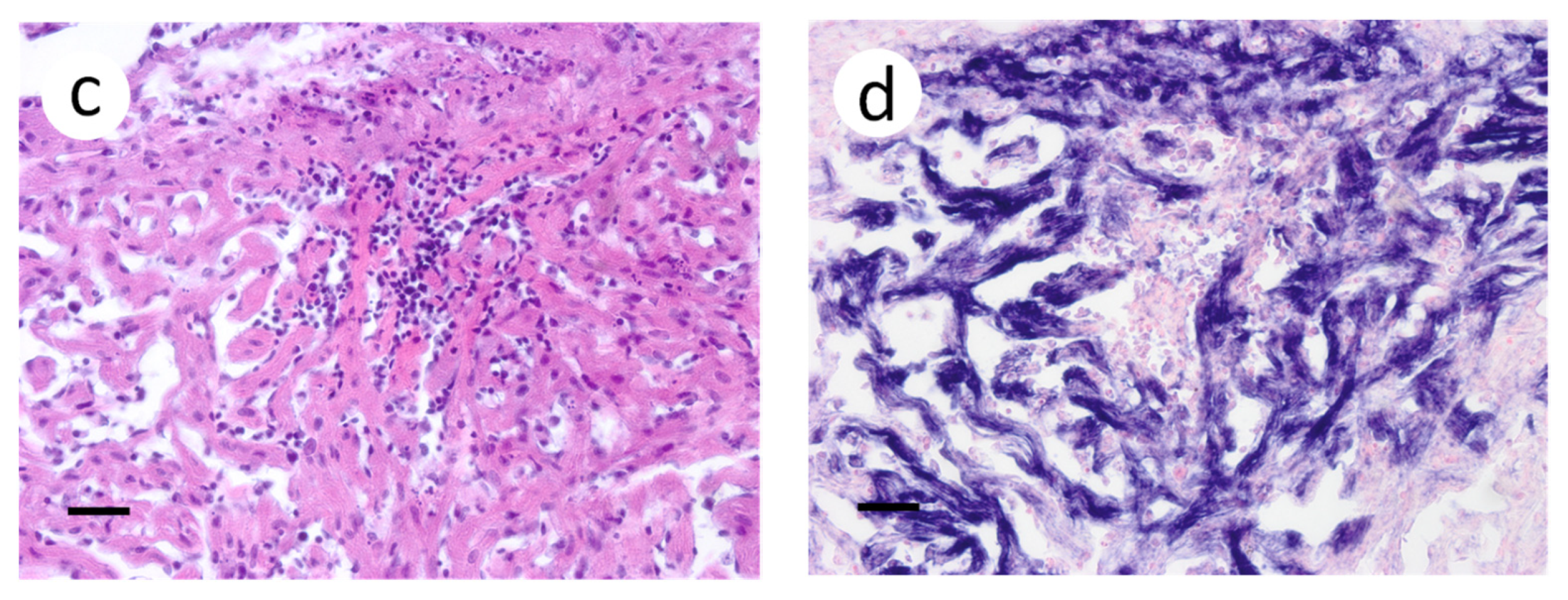
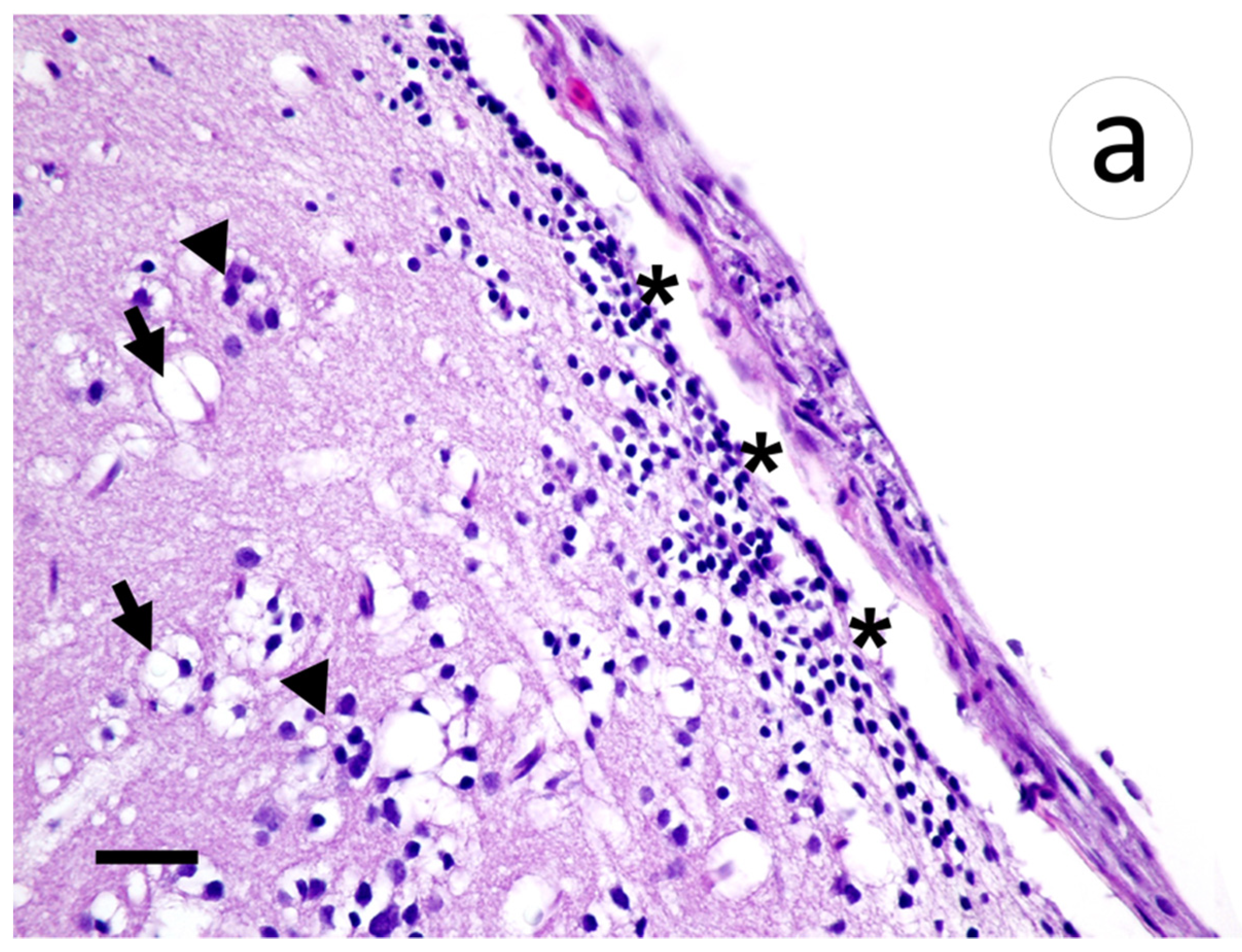
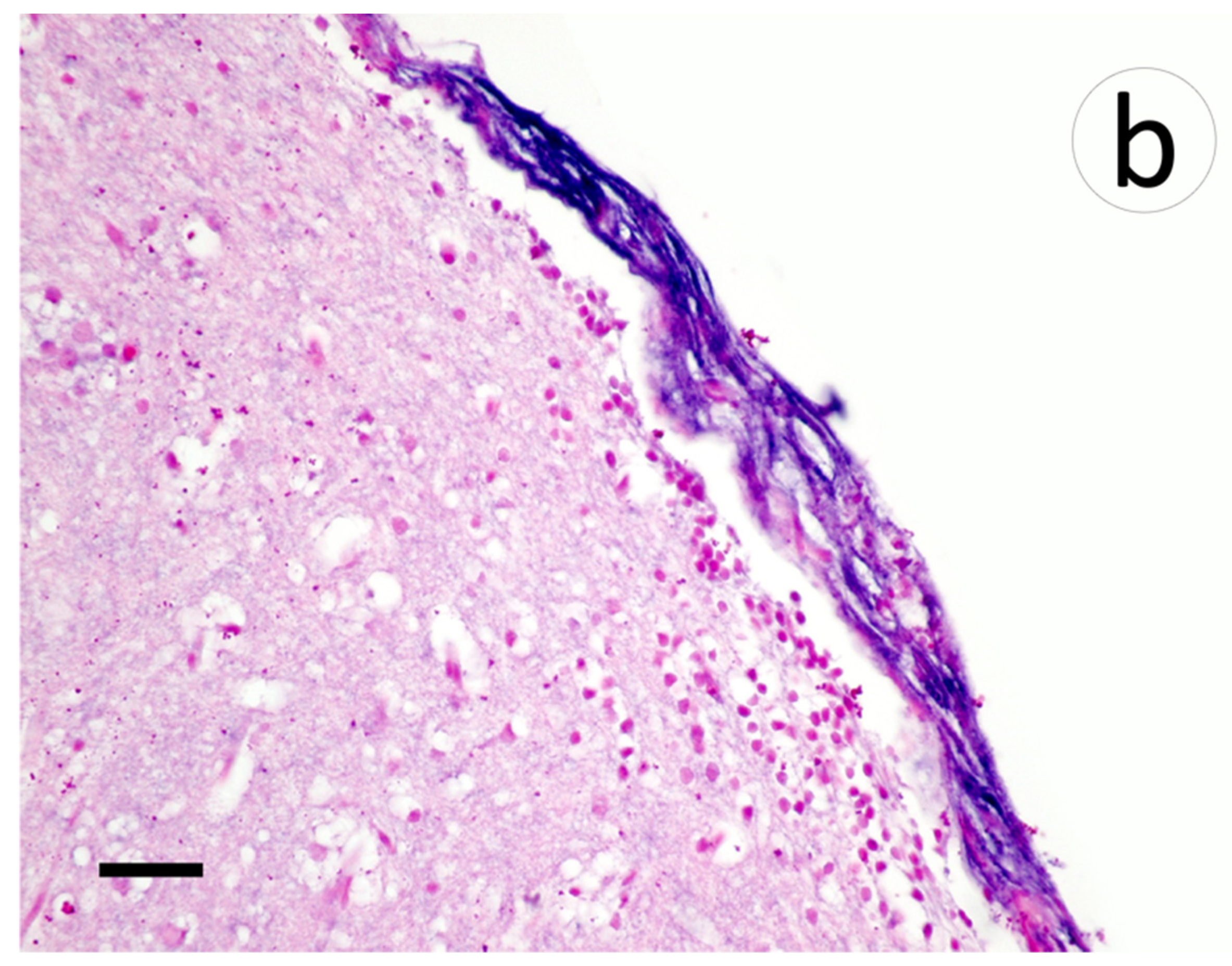
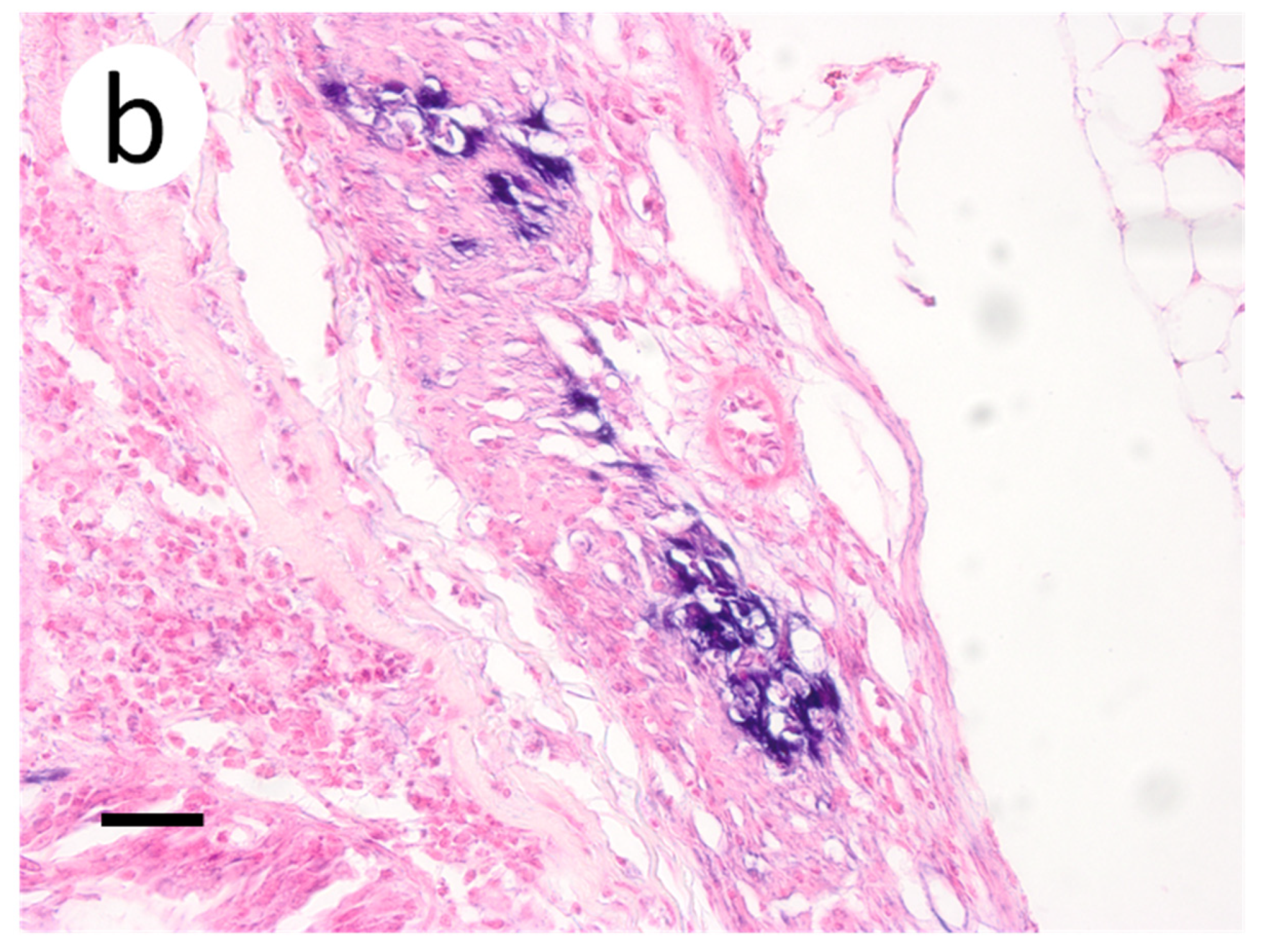
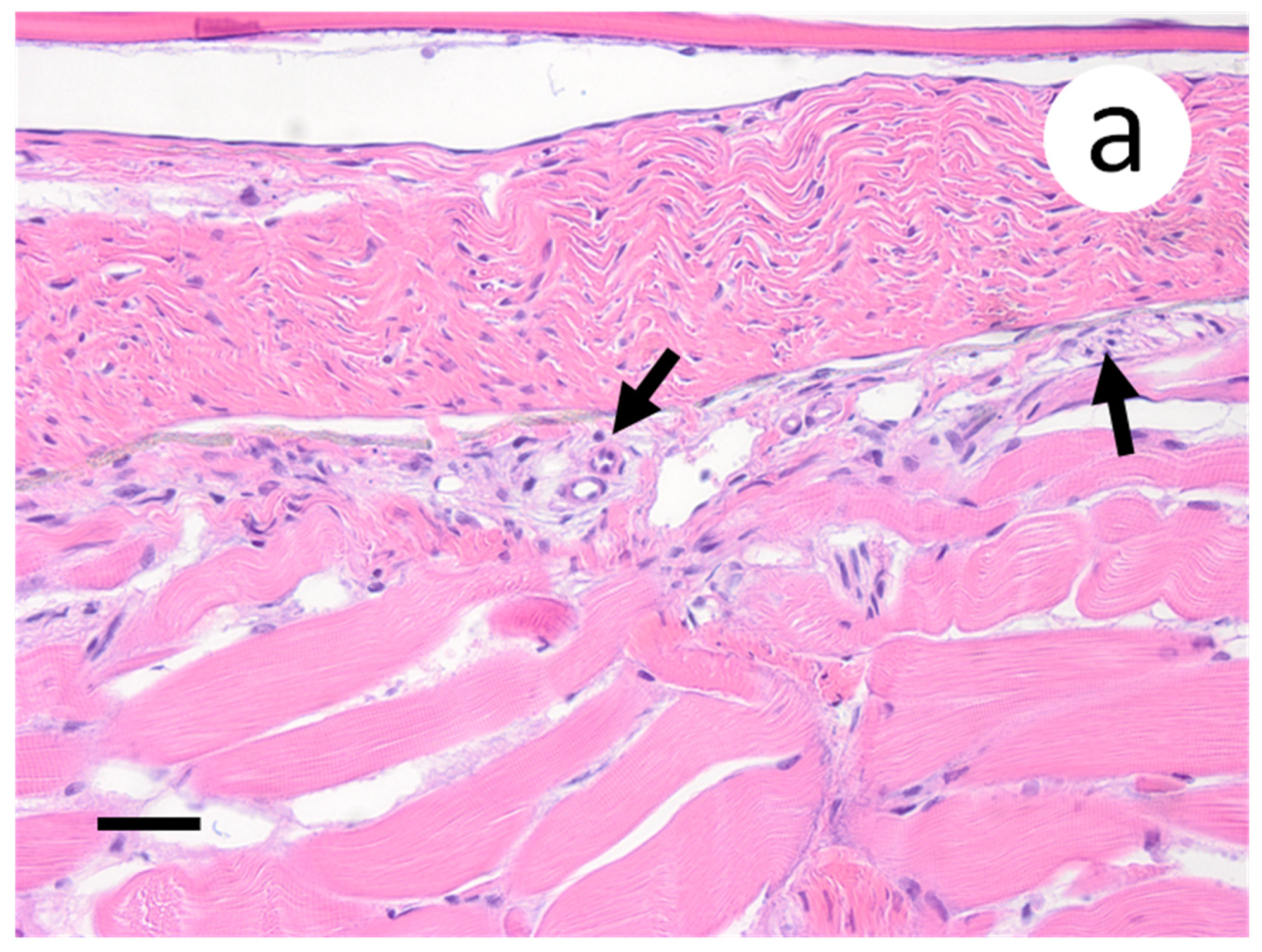
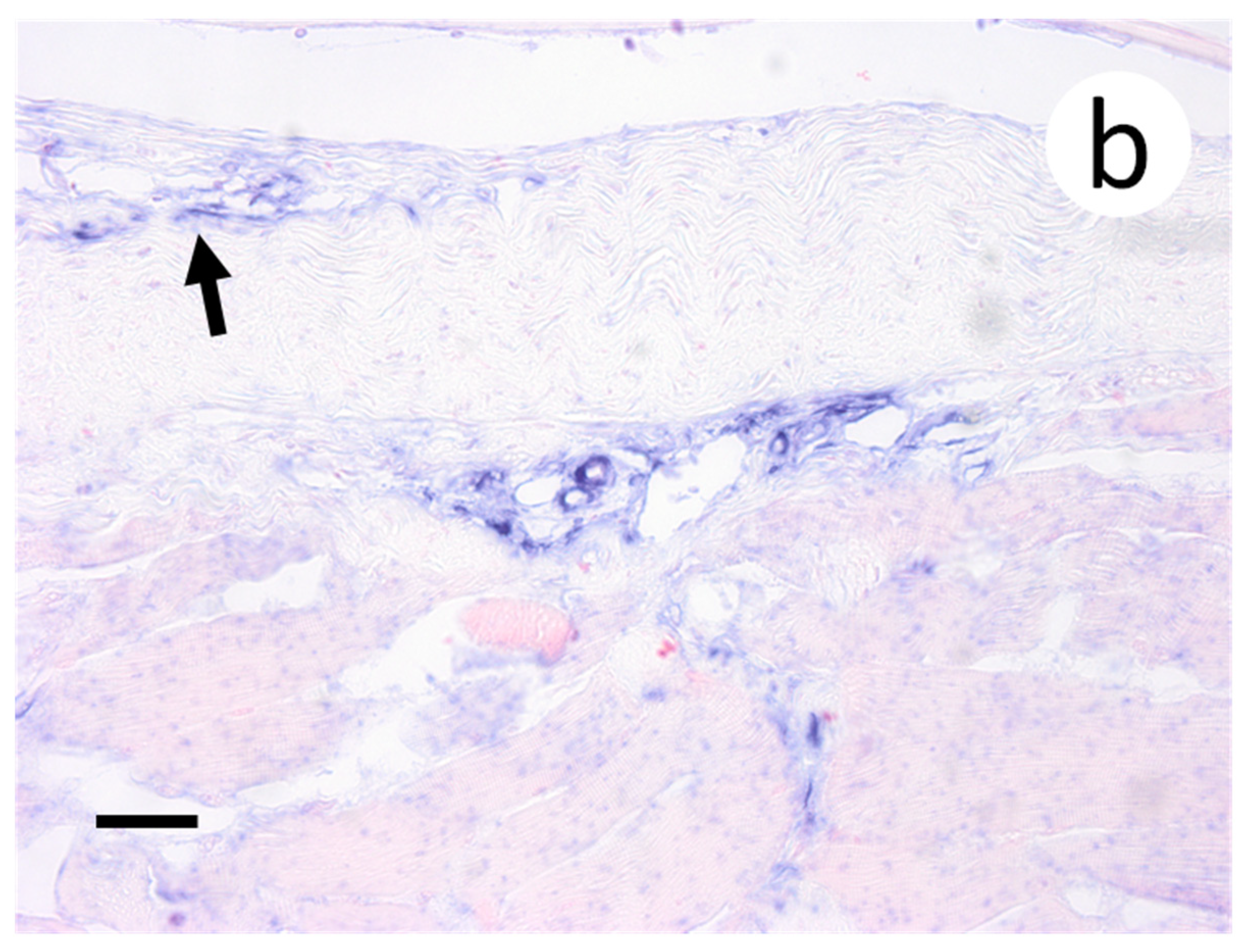
3.3. Visualization of VHSV-IVb in Tissues of Infected Muskellunge
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walker, P.J.; Siddell, S.G.; Lefkowitz, E.J.; Mushegian, A.R.; Adriaenssens, E.M.; Dempsey, D.M.; Dutilh, B.E.; Harrach, B.; Harrison, R.L.; Hendrickson, R.C.; et al. Changes to virus taxonomy and the statutes ratified by the international committee on taxonomy of viruses. Arch. Virol. 2020, 165, 2737–2748. [Google Scholar] [CrossRef]
- Wolf, K. Viral Hemorrhagic Septicemia. In Fish Viruses and Fish Viral Diseases; Wolf, K., Ed.; Comstock Publishing Associates: Cornell University Press, Ithaca, NY, USA and London, UK, 1988; pp. 217–249. ISBN 9780801412592. [Google Scholar]
- Smail, D.A. Viral haemorrhagic septicaemia. In Fish Diseases and Disorders: Viral, Bacterial, and Fungal Infections; Woo, P., Bruno, D.W., Eds.; CABI Publishing: Wallingford, Oxfordshire, UK, 1999; Volume 3, pp. 123–146. ISBN 978-0851991948. [Google Scholar]
- Elsayed, E.; Faisal, M.; Thomas, G.; Whelan, G.; Batts, W.; Winton, J. Isolation of viral haemorrhagic septicaemia virus from muskellunge, Esox masquinongy (Mitchill), in Lake St. Clair, Michigan, USA reveals a new sublineage of the North American genotype. J. Fish Dis. 2006, 29, 611–619. [Google Scholar] [CrossRef]
- Faisal, M.; Ahne, W. Spring viremia of carp virus (SVCV): Comparison of immunoperoxidase, fluorescent antibody and cell culture isolation techniques for detection of antigen. J. Fish Dis. 1984, 7, 57–64. [Google Scholar] [CrossRef]
- Stepien, C.A.; Niner, M.D. Evolutionary trajectory of fish Piscine Novirhabdovirus (=Viral Hemorrhagic Septicemia Virus) across its Laurentian Great Lakes History: Spatial and temporal diversification. Ecol. Evol. 2020, 10, 9740–9775. [Google Scholar] [CrossRef]
- Niner, M.D.; Stepien, C.A.; Gorgoglione, B.; Leaman, D.W. Genomic and immunogenic changes of Piscine Novirhabdovirus (Viral Hemorrhagic Septicemia Virus) over its evolutionary history in the Laurentian Great Lakes. PLoS ONE 2021, 16, e0232923. [Google Scholar] [CrossRef]
- Faisal, M.; Winters, A.D. Detection of viral hemorrhagic septicemia virus (VHSV) from Diporeia spp. (Pontoporeiidae, Amphipoda) in the Laurentian Great Lakes, USA. Parasit Vectors. 2011, 4, 201. [Google Scholar] [CrossRef] [Green Version]
- Faisal, M.; Schulz, C.A. Detection of viral hemorrhagic septicemia virus (VHSV) from the Leech Myzobdella lugubris Leidy, 1851. Parasit Vectors. 2009, 2, 45. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.; Faisal, M. Viral Hemorrhagic Septicemia Virus in The Great Lakes Basin. In Aquatic Animal Health: A Continuing Dialogue Between Russia and The United States. Proceedings of the Third Bilateral Conference Between the United States and Russia, July 2009; Cipriano, R.C., Bruckner, A., Shchelkunov, I.S., Eds.; Michigan State University Printing: East Lansing, MI, USA, 2011; pp. 113–123. [Google Scholar]
- Faisal, M.; Shavalier, M.; Kim, R.K.; Millard, E.V.; Gunn, M.R.; Winters, A.D.; Schulz, C.A.; Eissa, A.; Thomas, M.V.; Wolgamood, M.; et al. Spread of the emerging viral hemorrhagic septicemia virus strain, genotype IVb, in Michigan, USA. Viruses 2012, 4, 734–760. [Google Scholar] [CrossRef]
- Olesen, N.J.; Garver, K.; Kim, H.J. Viral Haemorrhagic Septicaemia. In Manual of Diagnostic Tests for Aquatic Animals, 7th ed.; Vallat, B., Ed.; World Organization for Animal Health: Paris, France, 2021; Chapter 2.3.10; ISBN 978-92-9044-887-7. [Google Scholar]
- Batts, W.N.; Lovy, J.; Getchell, R.G.; Faisal, M.; Standish, I.; Warg, J.V.; Phelps, N.B.D.; Glenney, G.; Winton, J.R. Viral Hemorrhagic Septicemia. In Afs-Fhs (American Fisheries Society-Fish Health Section). FHS Blue Book: Suggested Procedures for the Detection and Identification of Certain Finfish and Shellfish Pathogens, 2020 Edition. Available online: https://units.fisheries.org/fhs/fishhealth-section-blue-book-2020/ (accessed on 29 November 2021).
- Hope, K.M.; Casey, R.N.; Groocock, G.H.; Getchell, R.G.; Bowser, P.R.; Casey, J.W. Comparison of quantitative RT-PCR with cell culture to detect viral hemorrhagic septicemia virus (VHSV) IVb infections in the Great Lakes. J. Aquat. Anim. Health. 2010, 22, 50–61. [Google Scholar] [CrossRef]
- Brudeseth, B.E.; Raynard, R.S.; King, J.A.; Evensen, Ø. Sequential pathology after experimental infection with marine viral hemorrhagic septicemia virus isolates of low and high virulence in turbot (Scophthalmus maximus L.). Vet. Pathol. 2005, 42, 9–18. [Google Scholar] [CrossRef] [Green Version]
- Evensen, Ø.; Meier, W.; Wahli, T.; Olesen, N.J.; Vestergård Jørgensen, P.E.; Håstein, T. Comparison of immunohistochemistry and virus cultiviation for detection of viral haemorrhagic septicaemia virus in experimentally infected rainbow trout Oncorhynchus mykiss. Dis. Aquat. Organ. 1994, 20, 101–109. [Google Scholar] [CrossRef]
- Kim, R.; Faisal, M. Emergence and resurgence of the viral hemorrhagic septicemia virus (Novirhabdovirus, Rhabdoviridae, Mononegavirales). J. Adv. Res. 2011, 2, 9–23. [Google Scholar] [CrossRef] [Green Version]
- Kim, R.; Faisal, M. Comparative susceptibility of representative Great Lakes Fish Species to the North American viral hemorrhagic septicemia virus sublineage IVb. Dis. Aquat. Organ. 2010, 91, 23–34. [Google Scholar] [CrossRef]
- Weeks, C.; Kim, R.; Wolgamod, M.; Whelan, G.; Faisal, M. Experimental infection studies demonstrate the high susceptibility of the Salmonid, Lake Herring, Coregonus artedi (Le Sueur), to the Great Lakes strain of viral haemorrhagic septicaemia virus (Genotype IVb). J. Fish Dis. 2011, 34, 887–891. [Google Scholar] [CrossRef]
- Shavalier, M.; Faisal, M.; Loch, T.P.; Fitzgerald, S.D.; Thaiwong, T.; Kiupel, M. Disease progression in Lake Trout (Salvelinus Namaycush) experimentally infected with epizootic epitheliotropic disease virus (Salmonid Herpesvirus-3). Vet. Pathol. 2020, 57, 687–699. [Google Scholar] [CrossRef]
- Qadiri, S.S.N.; Kim, S.-J.; Krishnan, R.; Kim, J.-O.; Kim, W.-S.; Oh, M.-J. Development of an in-situ hybridization assay using riboprobes for detection of viral haemorrhagic septicemia virus (VHSV) mRNAs in a cell culture model. J. Virol. Methods. 2019, 264, 1–10. [Google Scholar] [CrossRef]
- Fijan, N.; Sulimanovic, D.; Bearzotti, M.; Muzinic, D.; Zwillenberg, L.O.; Chilmonczyk, S.; Vautherot, J.F.; de Kinkelin, P. Some properties of the Epithelioma papulosum cyprini (EPC) cell line from carp (Cyprinus carpio). Ann. Virol. 1983, 134, 207–220. [Google Scholar] [CrossRef]
- Batts, W.N.; Traxler, G.S.; Winton, J.R. Factors Affecting the efficiency of plating for selected fish rhabdoviruses. In Proceedings of the 2nd International Symposium on Viruses of Lower Vertebrates, Corvallis, OR, USA, 27–31 July 1991; Fryer, J.L., Ed.; Oregon State University: Corvallis, OR, USA, 1991; pp. 17–24. [Google Scholar]
- Kim, R.; Faisal, M. Experimental studies confirm the wide host range of the Great Lakes viral haemorrhagic septicaemia virus genotype IVb. J. Fish Dis. 2010, 33, 83–88. [Google Scholar] [CrossRef]
- Prophet, E.; Mills, B.; Arrington, J.; Sobin, L.H. Laboratory Methods in Histotechnology; Armed Forces Institute of Pathology, American Registry of Pathology: Washington, DC, USA, 1992; ISBN 188104100X 9781881041009. [Google Scholar]
- Menezes, R.C.; Figueiredo, F.B.; Wise, A.G.; Madeira, M.F.; Oliveira, R.V.; Schubach, T.M.; Kiupel, M.; Langohr, I.M. Sensitivity and specificity of in situ hybridization for diagnosis of cutaneous infection by leishmania infantum in dogs. J. Clin. Microbiol. 2013, 51, 206–211. [Google Scholar] [CrossRef] [Green Version]
- He, M.; Yan, X.C.; Liang, Y.; Sun, X.W.; Teng, C.B. Evolution of the viral hemorrhagic septicemia virus: Divergence, selection and origin. Mol. Phylogenet. Evol. 2014, 77, 34–40. [Google Scholar] [CrossRef]
- Kim, R.K.; Faisal, M. The Laurentian Great Lakes strain (MI03) of the viral haemorrhagic septicaemia virus is highly pathogenic for juvenile muskellunge, Esox masquinongy (Mitchill). J. Fish Dis. 2010, 33, 513–527. [Google Scholar] [CrossRef]
- Kerr, S.J. Distribution and Management of Muskellunge in North America: An Overview; Fisheries Policy Section, Biodiversity Branch, Ontario Ministry of Natural Resources: Peterborough, ON, Canada, 2011. [Google Scholar]
- Meier, W.; Jørgensen, P.E.V. Isolation of VHS virus from pike (Esox lucius L.) with hemorrhagic symptoms. In Fish Diseases. 3rd COPRAQ-Session; Ahne, W., Ed.; Springer-Verlag: Berlin/Heidelberg, Germany, 1980; pp. 8–17. ISBN 9783642678547. [Google Scholar]
- Meier, W.; Wahli, T. Viral haemorrhagic septicaemia (VHS) in grayling (Thyn allusthymallus L.). J. Fish. Dis. 1988, 11, 481–487. [Google Scholar] [CrossRef]
- Kibenge, F.S.; Whyte, S.K.; Hammell, K.L.; Rainnie, D.; Kibenge, M.T.; Martin, C.K. A Dual infection of infectious Salmon Anaemia (ISA) virus and a togavirus-like virus in ISA of Atlantic salmon Salmo salar in New Brunswick, Canada. Dis. Aquat. Organ. 2000, 42, 11–15. [Google Scholar] [CrossRef]
- Plumb, J.A.; Hanson, L.A. Health Maintenance and Principal Microbial Diseases of Cultured Fishes, 3rd ed.; Blackwell Publishing Ltd.: Ames, IA, USA, 2011; ISBN 9780813816937. [Google Scholar]
- Miller, T.A.; Rapp, J.; Wastlhuber, U.; Hoffmann, R.W.; Enzmann, P.J. Rapid and sensitive reverse transcriptase-polymerase chain reaction based detection and differential diagnosis of fish pathogenic rhabdoviruses in organ samples and cultured cells. Dis. Aquat. Organ. 1998, 34, 13–20. [Google Scholar] [CrossRef] [Green Version]
- Oidtmann, B.; Joiner, C.; Stone, D.; Dodge, M.; Reese, R.A.; Dixon, P. viral load of various tissues of rainbow trout challenged with viral haemorrhagic septicaemia virus at various stages of disease. Dis. Aquat. Organ. 2011, 93, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Lovy, J.; Lewis, N.L.; Hershberger, P.K.; Bennett, W.; Meyers, T.R.; Garver, K.A. Viral tropism and pathology associated with viral hemorrhagic septicemia in larval and juvenile pacific herring. Vet. Microbiol. 2012, 161, 66–76. [Google Scholar] [CrossRef]
- Harmache, A.; LeBerre, M.; Droineau, S.; Giovannini, M.; Brémont, M. Bioluminescence Imaging of Live Infected Salmonids Reveals that the Fin Bases are the Major Portal of Entry for Novirhabdovirus. J. Virol. 2006, 80, 3655–3659. [Google Scholar] [CrossRef] [Green Version]
- Neukirch, M. Uptake, multiplication, and excretion of viral haemorrhagic septicaemia virus in rainbow trout (Salmo gairdneri). In Fish and Shellfish Pathology; Ellis, A.E., Ed.; Academic Press, Inc.: London, UK, 1985; pp. 295–300. ISBN 0122374908 9780122374906. [Google Scholar]
- Kim, S.-J.; Kim, W.-S.; Oh, M.-J. Differences of viral hemorrhagic septicemia virus loads among organs of dead and surviving olive flounder infected by intramuscular injection and immersion challenge. J. Aquat. Anim. Health. 2019, 31, 193–200. [Google Scholar] [CrossRef]
















{kind=link}
{kind=link}
{kind=link}
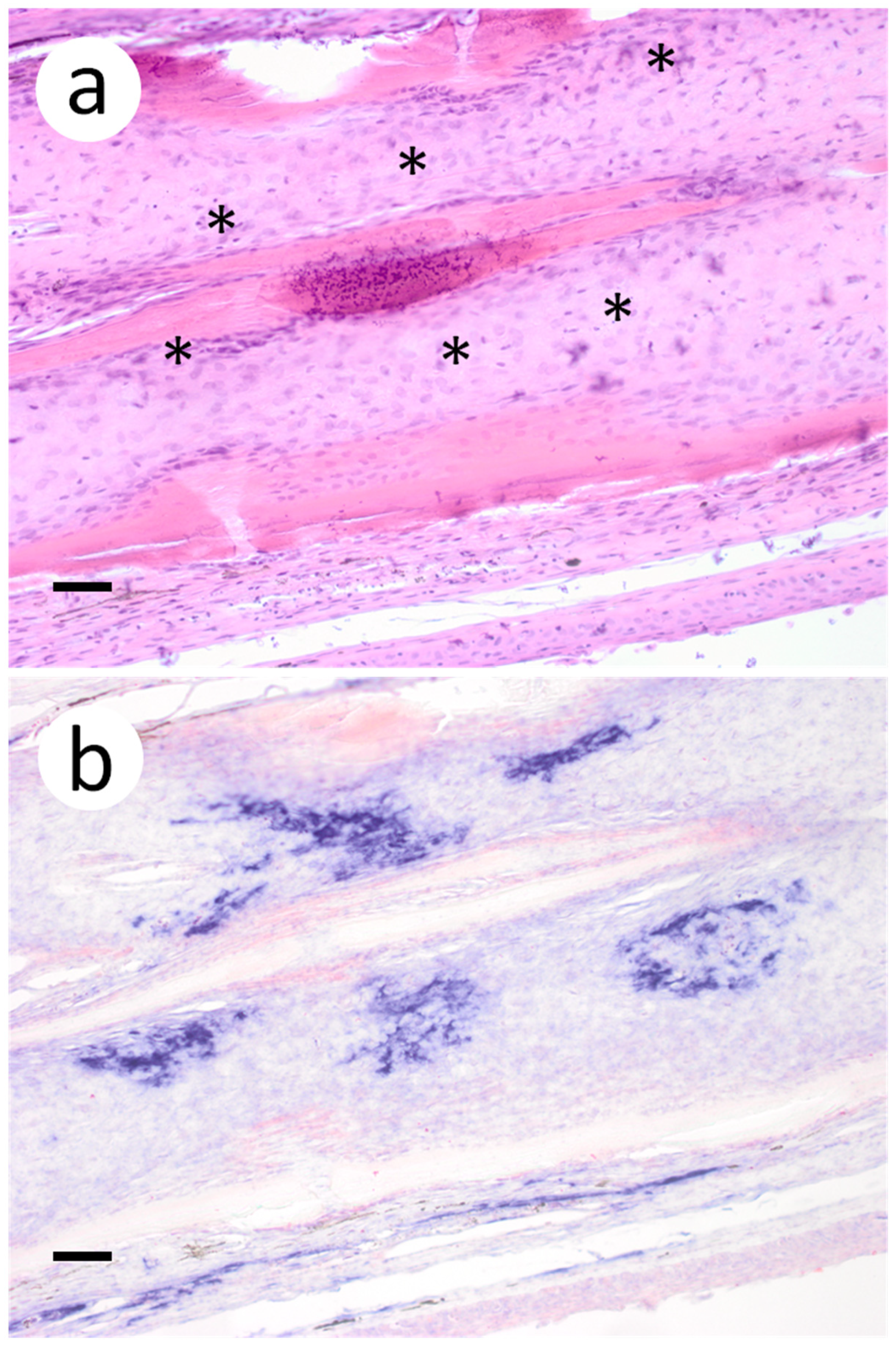{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| VHSV Positive Samples (n= 4 per Sampling Time) | Range of Viral Load In Positive Tissues (PFU g−1) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hours | Days | ||||||||||||||||||
| Tissue | 0 h | 6 h | 12 h | 24 h | 36 h | 2 d | 4 d | 6 d | 8 d | 15 d | 22 d | 29 d | 36 d | 43 d | 50 d | 57 d | 64 d | Total | |
| spleen | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 4 | 1.98 × 102–4.46 × 106 |
| kidney | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 2 | 1 | 0 | 0 | 0 | 0 | 1 | 1 | 8 | 1.20 × 103–1.56 × 107 |
| heart | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | 1 | 2 | 1 | 2 | 1 | 0 | 1 | 2 | 2 | 16 | 2.17 × 102–4.26 × 107 |
| liver | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 | 2 | 3 | 2 | 2 | 0 | 0 | 1 | 2 | 16 | 4.51 × 102–5.95 × 105 |
| intestine | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 1 | 1 | 2 | 0 | 0 | 0 | 2 | 9 | 1.83 × 103–1.95 × 106 |
| gill | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 1 | 1 | 2 | 2 | 1 | 0 | 0 | 1 | 3 | 13 | 5.29 × 102–9.89 × 106 |
| pectoral fin | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 2 | 2 | 2 | 1 | 1 | 0 | 3 | 1 | 1 | 2 | 16 | 1.85 × 102–6.63 × 107 |
| skin/muscle | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 2 | 3 | 1 | 2 | 0 | 3 | 0 | 0 | 1 | 14 | 1.90 × 102–4.81 × 105 |
| Total | - | - | - | - | - | 3 | - | 15 | 8 | 15 | 10 | 10 | 6 | 7 | 2 | 7 | 13 | 96 | |
| VHSV Positive Samples (n= 4 per Sampling Time) | Range of Viral Load In Positive Tissues (PFU g−1) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hours | Days | ||||||||||||||||||
| Tissue | 0 h | 6 h | 12 h | 24 h | 36 h | 2 d | 4 d | 6 d | 8 d | 15 d | 22 d | 29 d | 36 d | 43 d | 50 d | 57 d | 64 d | Total | |
| spleen | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 | 0 | 2 | 0 | 2 | 0 | 0 | 1 | 7 | 1.85 × 104–1.30 × 108 |
| kidney | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 1 | 1 | 2 | 0 | 2 | 0 | 0 | 1 | 10 | 5.96 × 102–2.63 × 107 |
| heart | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 1 | 2 | 3 | 0 | 3 | 0 | 0 | 3 | 15 | 5.72 × 103–5.93 × 108 |
| liver | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 2 | 1 | 2 | 0 | 2 | 0 | 0 | 1 | 10 | 8.21 × 102–5.20 × 108 |
| intestine | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 1 | 2 | 4 | 0 | 2 | 0 | 0 | 0 | 11 | 1.23 × 103–3.17 × 107 |
| gill | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 0 | 2 | 2 | 3 | 0 | 2 | 0 | 1 | 1 | 14 | 1.49 × 103–5.04 × 108 |
| pectoral fin | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 3 | 0 | 1 | 0 | 3 | 0 | 2 | 1 | 0 | 1 | 12 | 5.15 × 103–8.16 × 107 |
| skin/muscle | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1 | 3 | 0 | 3 | 0 | 3 | 1 | 0 | 1 | 15 | 1.31 × 102–2.82 × 107 |
| Total | - | - | - | 1 | - | - | - | 20 | 1 | 12 | 8 | 22 | - | 18 | 2 | 1 | 9 | 94 | |
| VHSV Positive Samples (n= 4 per Sampling Time) | Range of Viral Load In Positive Tissues (PFU g−1) | ||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Hours | Days | ||||||||||||||||||
| Tissue | 0 h | 6 h | 12 h | 24 h | 36 h | 2 d | 4 d | 6 d | 8 d | 15 d | 22 d | 29 d | 36 d | 43 d | 50 d | 57 d | 64 d | Total | |
| spleen | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| kidney | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| heart | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1.58 × 102 |
| liver | 0 | 0 | 0 | 1 | 0 | 0 | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 3 | 1.08 × 103–2.76 × 103 |
| intestine | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 1 | 0 | 0 | 0 | 3 | 1.78 × 102–1.97 × 104 |
| gill | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2.74 × 103 |
| pectoral fin | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 4.47 × 103 |
| skin/muscle | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | - |
| Total | - | - | - | 1 | 1 | 1 | 1 | - | - | 1 | 2 | 1 | - | 1 | - | - | - | 9 | |
| Tissues | Estimate β | OR of Tissue vs. Kidney (eβ) | 95% Confidence Limit for OR | |
|---|---|---|---|---|
| Liver | 0.7847 | 2.1917 | 1.3860 | 3.4660 |
| Heart | 0.6025 | 1.8267 | 1.2411 | 2.6885 |
| Skin/muscle | 0.5197 | 1.6815 | 1.1073 | 2.5536 |
| Gill | 0.3586 | 1.4313 | 1.0096 | 2.0293 |
| Pectoral fin | 0.3056 | 1.3574 | 0.9945 | 1.8528 |
| Intestine | 0.2151 | 1.2400 | 0.7857 | 1.9570 |
| Kidney | 0.0000 | 1.0000 | - | - |
| Spleen | −0.3215 | 0.7251 | 0.4377 | 1.2009 |
| Kidney | Heart | Liver | Intestine | Gill | Pectoral Fin | Skin/Muscle | |
|---|---|---|---|---|---|---|---|
| spleen | 0.2339 | <0.0001 | <0.0001 | 0.0638 | 0.0030 | 0.0014 | 0.0012 |
| kidney | - | 0.0003 | 0.0005 | 0.3489 | 0.0150 | 0.0023 | 0.0150 |
| heart | - | - | 0.6331 | 0.0138 | 0.1435 | 0.3299 | 0.4112 |
| liver | - | - | - | 0.0042 | 0.0673 | 0.0673 | 0.2354 |
| intestine | - | - | - | - | 0.2838 | 0.2484 | 0.2075 |
| gill | - | - | - | - | - | 0.7493 | 0.7340 |
| pectoral fin | - | - | - | - | - | - | 0.9424 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kim, R.K.; Fitzgerald, S.D.; Kiupel, M.; Faisal, M. Tissue Distribution of the Piscine Novirhabdovirus Genotype IVb in Muskellunge (Esox masquinongy). Animals 2022, 12, 1624. https://doi.org/10.3390/ani12131624
Kim RK, Fitzgerald SD, Kiupel M, Faisal M. Tissue Distribution of the Piscine Novirhabdovirus Genotype IVb in Muskellunge (Esox masquinongy). Animals. 2022; 12(13):1624. https://doi.org/10.3390/ani12131624
Chicago/Turabian StyleKim, Robert K., Scott D. Fitzgerald, Matti Kiupel, and Mohamed Faisal. 2022. "Tissue Distribution of the Piscine Novirhabdovirus Genotype IVb in Muskellunge (Esox masquinongy)" Animals 12, no. 13: 1624. https://doi.org/10.3390/ani12131624
APA StyleKim, R. K., Fitzgerald, S. D., Kiupel, M., & Faisal, M. (2022). Tissue Distribution of the Piscine Novirhabdovirus Genotype IVb in Muskellunge (Esox masquinongy). Animals, 12(13), 1624. https://doi.org/10.3390/ani12131624