
Sex Determination in Immature Sierra Nevada Lizard (Timon nevadensis)



Abstract
:Simple Summary
Abstract
1. Introduction
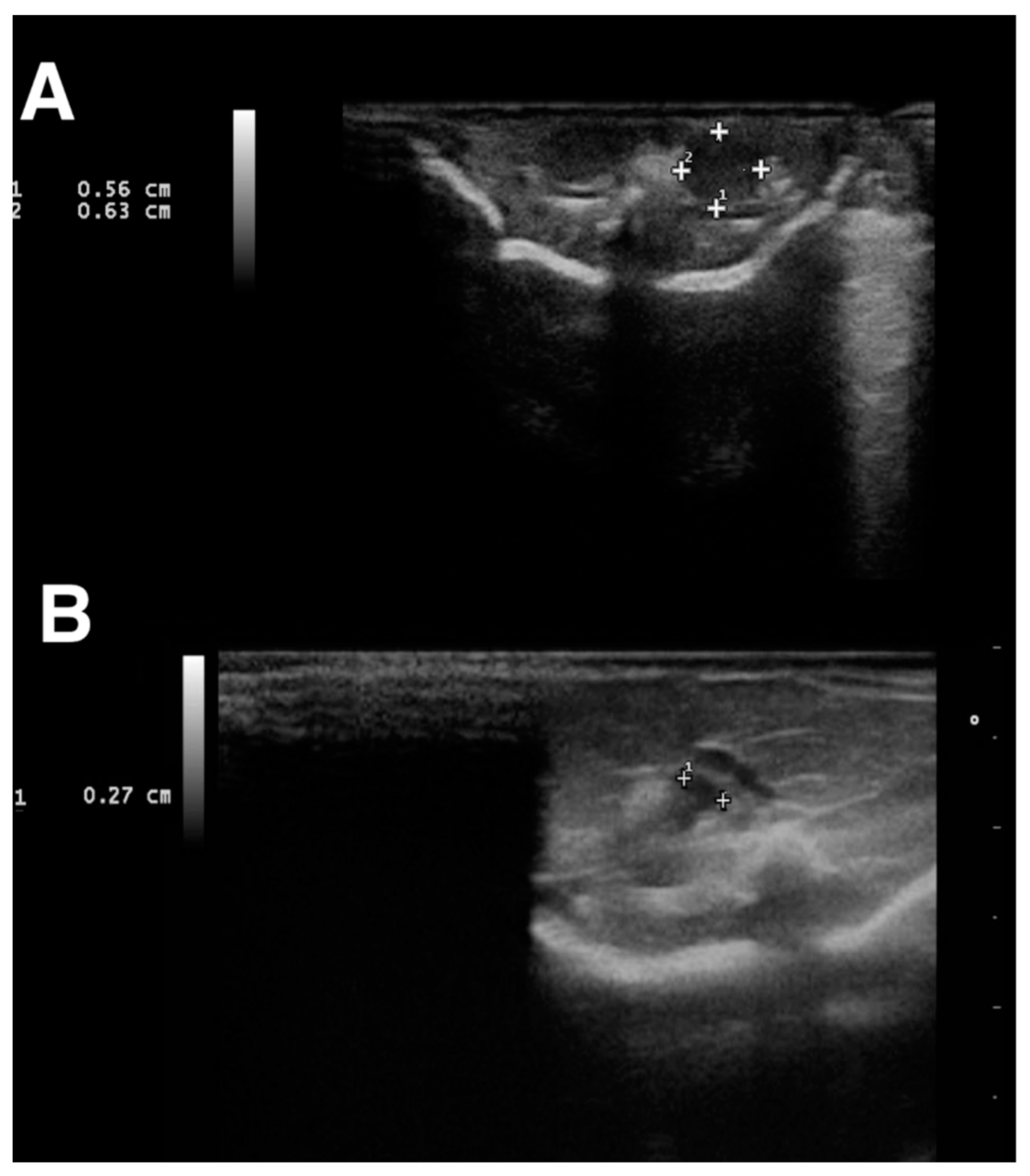
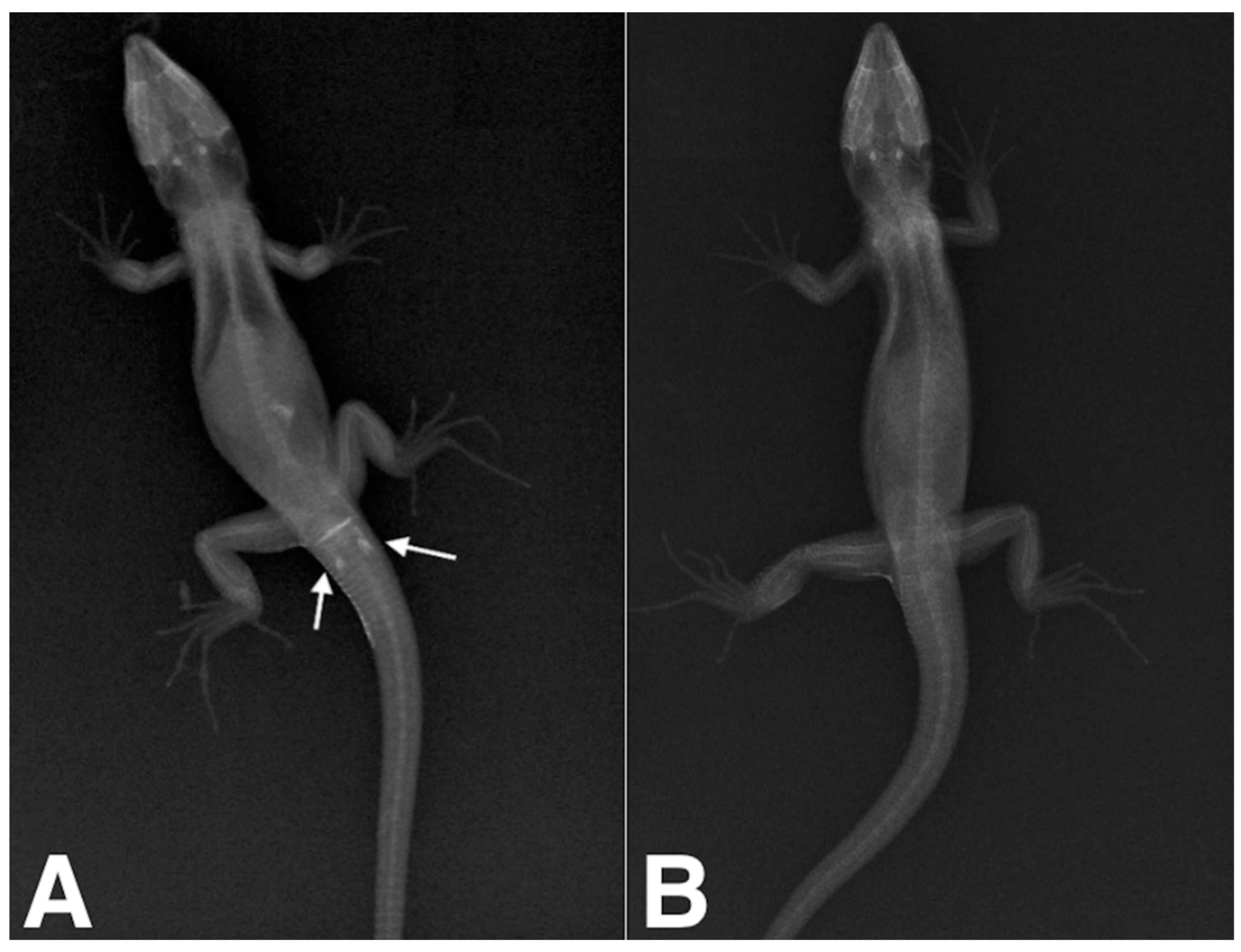
2. Materials and Methods
Statistical Analysis
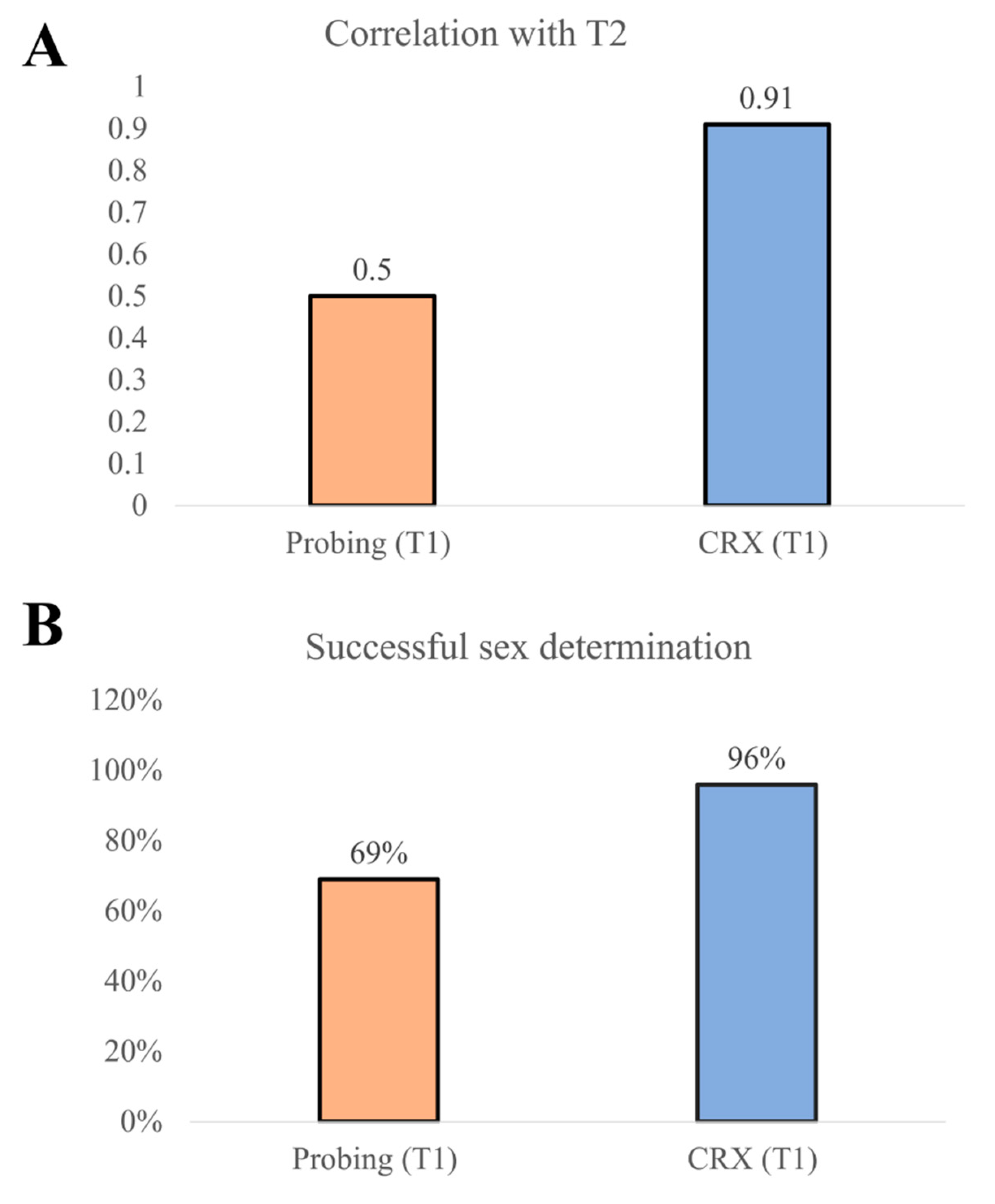
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Abbreviations
| CT | computed tomography |
| EV | endovenous |
| ND | not determined |
| NT | near threatened |
| VD | ventro-dorsal |
References
- Ahmadzadeh, F.; Flecks, M.; Carretero, M.A.; Böhme, W.; Ihlow, F.; Kapli, P.; Rödder, D. Separate histories in both sides of the Mediterranean: Phylogeny and niche evolution of ocellated lizards. J. Biogeogr. 2016, 43, 1242–1253. [Google Scholar] [CrossRef]
- Carretero, M.Á.; Salvador Milla, A. Lagarto Bético–Timon Nevadensis (Buchholz, 1963); Museo Nacional de Ciencias Naturales: Madrid, Spain, 2017. [Google Scholar]
- Braña, F. Sexual dimorphism in lacertid lizards: Male head increase vs female abdomen increase? Oikos 1996, 75, 511–523. [Google Scholar] [CrossRef]
- Mateo, J.A.; Milla, A.S. Lagarto Ocelado—Timon Lepidus (Daudin, 1802); Milla, S., Marco, A., Eds.; Museo Nacional de Ciencias Naturales: Madrid, Spain, 2017. [Google Scholar]
- Pellett, S.; Cope, I. Visual identification of reptiles: Part 2—lizards. Companion Anim. 2013, 18, 399–405. [Google Scholar] [CrossRef]
- Hailey, A.; Willemsen, R.E. Population density and adult sex ratio of the tortoise Testudo hermanni in Greece: Evidence for intrinsic population regulation. J. Zool. 2000, 251, 325–338. [Google Scholar] [CrossRef]
- Golubović, A.; Arsovski, D.; Tomović, L.; Bonnet, X. Is sexual brutality maladaptive under high population density? Biol. J. Linn. Soc. 2018, 124, 394–402. [Google Scholar] [CrossRef]
- Vetere, A.; Di Ianni, F.; Bertocchi, M.; Castiglioni, V.; Nardini, G. Unilateral ovarian torsion in a Moroccan eyed lizard (Timon tangitanus). J. Exot. Pet Med. 2022, 41, 46–47. [Google Scholar] [CrossRef]
- Sykes, J.M.I.V. Updates and practical approaches to reproductive disorders in reptiles. Vet. Clin. Exot. Anim. Pract. 2010, 13, 349–373. [Google Scholar] [CrossRef]
- Shea, G.M.; Reddacliff, G.L. Ossifications in the hemipenes of varanids. J. Herpetol. 1986, 20, 566–568. [Google Scholar] [CrossRef]
- Arnold, E.N. The hemipenis of lacertid lizards (Reptilia: Lacertidae): Structure, variation and systematic implications. J. Nat. Hist. 1986, 20, 1221–1257. [Google Scholar] [CrossRef]
- Di Ianni, F.; Volta, A.; Pelizzone, I.; Manfredi, S.; Gnudi, G.; Parmigiani, E. Diagnostic sensitivity of ultrasound, radiography and computed tomography for gender determination in four species of lizards. Vet. Radiol. Ultrasound 2015, 56, 40–45. [Google Scholar] [CrossRef]
- Knotek, Z.; Cermakova, E.; Oliveri, M. Reproductive medicine in lizards. Vet. Clin. Exot. Anim. Pract. 2017, 20, 411–438. [Google Scholar] [CrossRef] [PubMed]
- Denardo, D. Reproductive biology. In Reptile Medicine and Surgery; Mader, D., Ed.; Saunders-Elsiever: St. Louis, MO, USA, 2006; pp. 376–390. [Google Scholar]
- Funk, R.S. Lizard reproductive medicine and surgery. Vet. Clin. Exot. Anim. Pract. 2002, 5, 579–613. [Google Scholar] [CrossRef]
- Stahl, S.J. Veterinary management of snake reproduction. Vet. Clin. Exot. Anim. Pract. 2002, 5, 615–636. [Google Scholar] [CrossRef]
- Morris, P.J.; Alberts, A.C. Determination of sex in white-throated monitors (Varanus albigularis), gila monsters (Heloderma suspectum), and beaded lizards (H. horridum) using two-dimensional ultrasound imaging. J. Zoo Wildl. Med. 1996, 27, 371–377. [Google Scholar]
- Gartrell, B.D.; Girling, J.E.; Edwards, A.; Jones, S.M. Comparison of noninvasive methods for the evaluation of female reproductive condition in a large viviparous lizard, Tiliqua nigrolutea. Zoo Biol. 2002, 21, 253–268. [Google Scholar] [CrossRef]
- Gnudi, G.; Volta, A.; Di Ianni, F.; Bonazzi, M.; Manfredi, S.; Bertoni, G. Use of ultrasonography and contrast radiography for snake gender determination. Vet. Radiol. Ultrasound 2009, 50, 309–311. [Google Scholar] [CrossRef]
- Iannucci, A.; Altmanová, M.; Ciofi, C.; Ferguson-Smith, M.; Milan, M.; Pereira, J.C.; Pether, J.; Rehák, I.; Rovatsos, M.; Stanyon, R.; et al. Conserved sex chromosomes and karyotype evolution in monitor lizards (Varanidae). Heredity 2019, 123, 215–227. [Google Scholar] [CrossRef]
- Manukyan, L.; Montandon, S.A.; Fofonjka, A.; Smirnov, S.; Milinkovitch, M.C. A living mesoscopic cellular automaton made of skin scales. Nature 2017, 544, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Urbanová, D.; Halán, M. The use of ultrasonography in diagnostic imaging of reptiles. Folia Vet. 2016, 60, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Berthelet, A.; Bulliot, C. Imaging of reptiles-a sexing technique for squamates by ultrasound. Point Vét. 2014, 45, 6–7. [Google Scholar]
- Morris, P.J.; Jackintell, L.A.; Alberts, A.C. Predicting the gender of subadult Komodo dragons (Varanus komodoensis) using two-dimensional ultrasound imaging and plasma testosterone concentration. Zoo Biol. 1996, 15, 341–348. [Google Scholar] [CrossRef]
- Wibbels, T.; Lutz, P.L.; Musick, J.A.; Wyneken, J. Critical approaches to sex determination in sea turtles. Biol. Sea Turtles 2003, 2, 103–134. [Google Scholar]
- Cree, A.; Cockrem, J.F.; Brown, M.A.; Watson, P.R.; Guillette, L.J., Jr.; Newman, D.G.; Chambers, G.K. Laparoscopy, radiography, and blood analyses as techniques for identifying the reproductive condition of female tuatara. Herpetologica 1991, 47, 238–249. [Google Scholar]
- Innis, C.J. Endoscopy and endosurgery of the chelonian reproductive tract. Vet. Clin. Exot. Anim. Pract. 2010, 13, 243–254. [Google Scholar] [CrossRef] [PubMed]
- Mannion, P. (Ed.) Diagnostic Ultrasound in Small Animal Practice; John Wiley & Sons: Hoboken, NJ, USA, 2008. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| No. | Probing (T1) | Contrast Radiography [Crx (T1)] | 8 Months Ultrasound Examination (T2) | One Year Visual Check (T3) | Lenght T0, T1 (cm) | Weight (g) (T1) |
|---|---|---|---|---|---|---|
| 1 | M | M | M | M | 10 | 14 |
| 2 | ND | M | M | M | 9 | 11 |
| 3 | ND | F | F | F (Deceased) | 9 | 12 |
| 4 | F | F | F | F | 11 | 15 |
| 5 | M | M | M | M (Deceased) | 6 | 10 |
| 6 | ND | M | M | M | 10 | 14 |
| 7 | M | M | M | M | 9 | 12 |
| 8 | F | F | F | F | 11 | 16 |
| 9 | ND | M | M | M | 10 | 15 |
| 10 | M | M | M | M | 10 | 16 |
| 11 | F | F | F | F | 11 | 17 |
| 12 | F | F | F | F | 10 | 15 |
| 13 | M | M | M | M | 9 | 11 |
| 14 | M | M | M | M | 9 | 12 |
| 15 | M | M | M | M | 9 | 12 |
| 16 | F | F | M | M | 12 | 18 |
| 17 | F | F | F | F | 11 | 15 |
| 18 | M | M | M | M | 11 | 16 |
| 19 | F | M | M | M | 8 | 12 |
| 20 | M | M | M | ND (Deceased) | 11 | 15 |
| 21 | F | F | F | F | 9 | 10 |
| 22 | M | M | M | M | 10 | 13 |
| 23 | M | F | F | F | 11 | 15 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vetere, A.; Ablondi, M.; Bigliardi, E.; Rizzi, M.; Di Ianni, F. Sex Determination in Immature Sierra Nevada Lizard (Timon nevadensis). Animals 2022, 12, 2144. https://doi.org/10.3390/ani12162144
Vetere A, Ablondi M, Bigliardi E, Rizzi M, Di Ianni F. Sex Determination in Immature Sierra Nevada Lizard (Timon nevadensis). Animals. 2022; 12(16):2144. https://doi.org/10.3390/ani12162144
Chicago/Turabian StyleVetere, Alessandro, Michela Ablondi, Enrico Bigliardi, Matteo Rizzi, and Francesco Di Ianni. 2022. "Sex Determination in Immature Sierra Nevada Lizard (Timon nevadensis)" Animals 12, no. 16: 2144. https://doi.org/10.3390/ani12162144
APA StyleVetere, A., Ablondi, M., Bigliardi, E., Rizzi, M., & Di Ianni, F. (2022). Sex Determination in Immature Sierra Nevada Lizard (Timon nevadensis). Animals, 12(16), 2144. https://doi.org/10.3390/ani12162144