
The Odontocete Ear Canal-Associated Lymphoid Tissue (ECALT) and Lymph Nodes: Morphological and Pathological Description with Immuno-Phenotypic Characterisation
 ,
,  , , ,
, , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Tissue Sampling and Processing
2.3. Immunohistochemistry (IHC)
3. Results
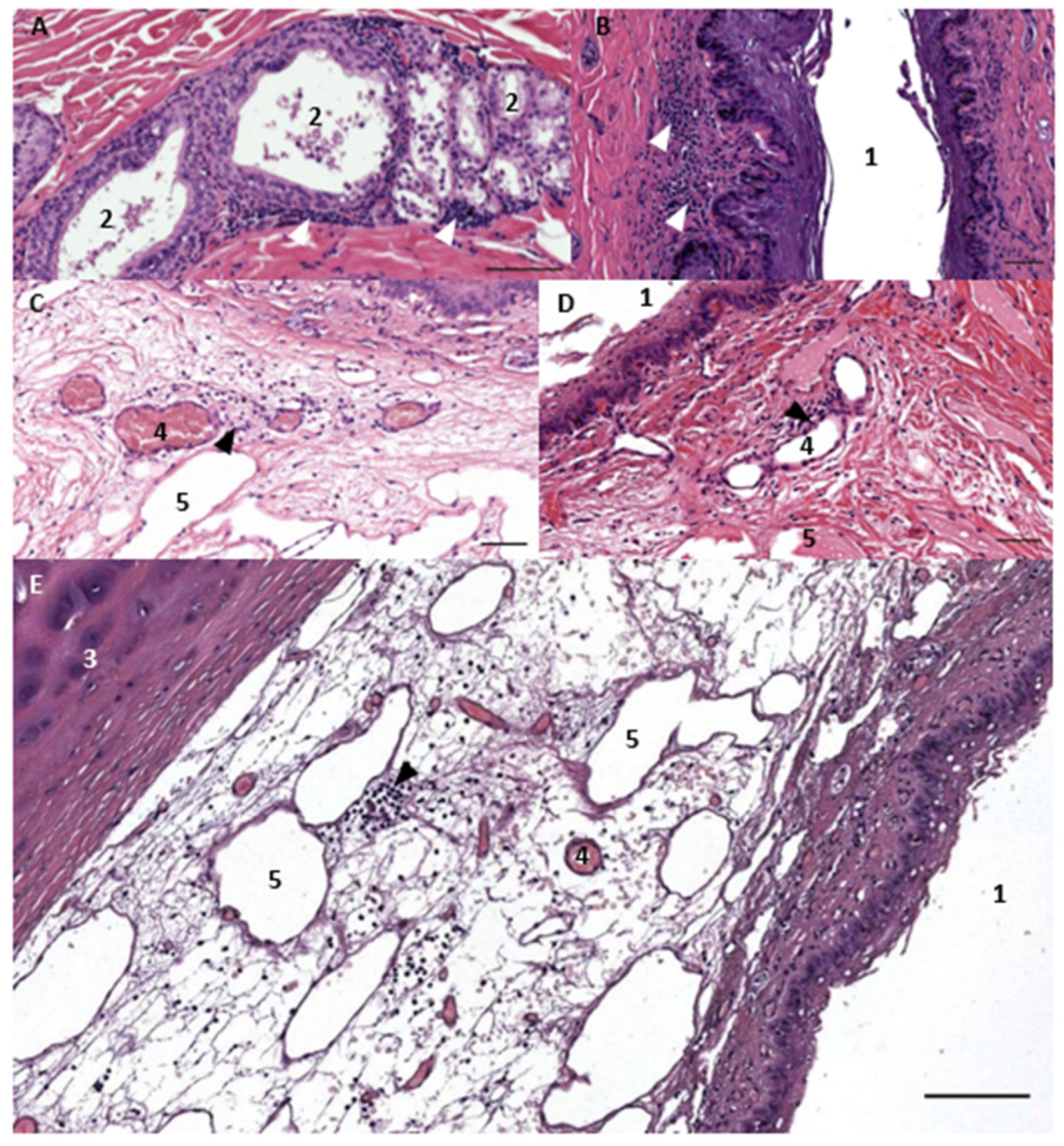
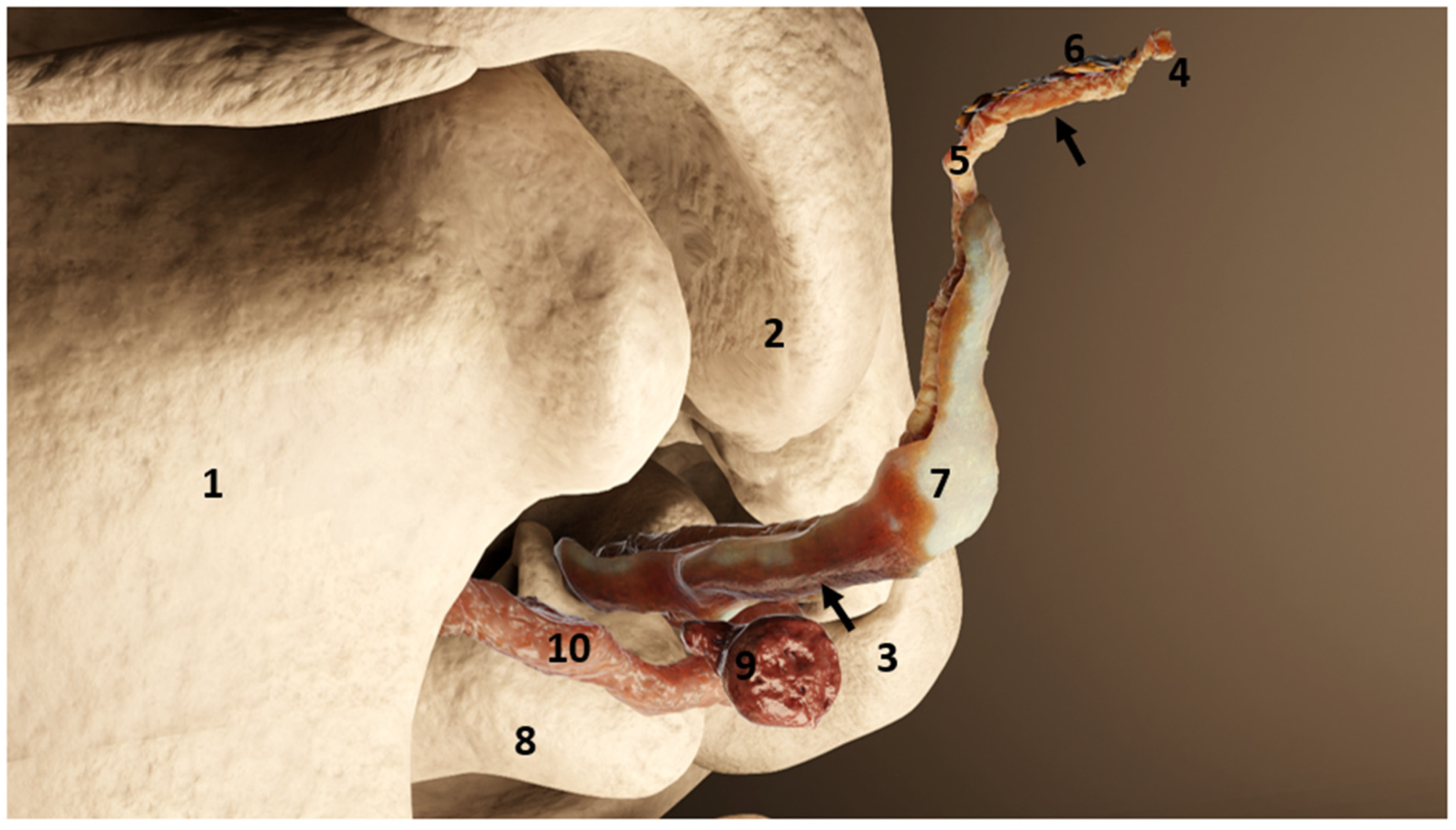
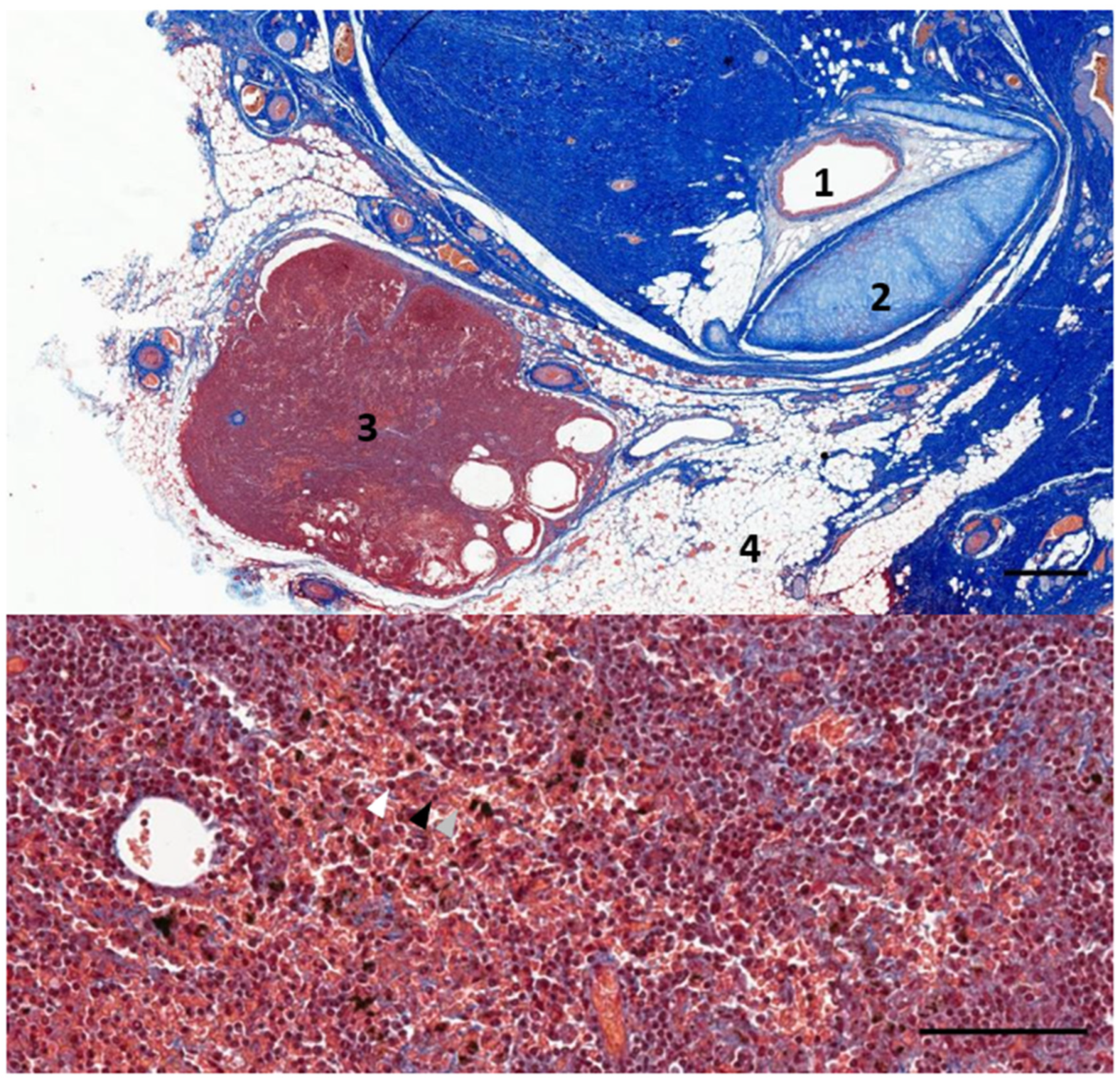
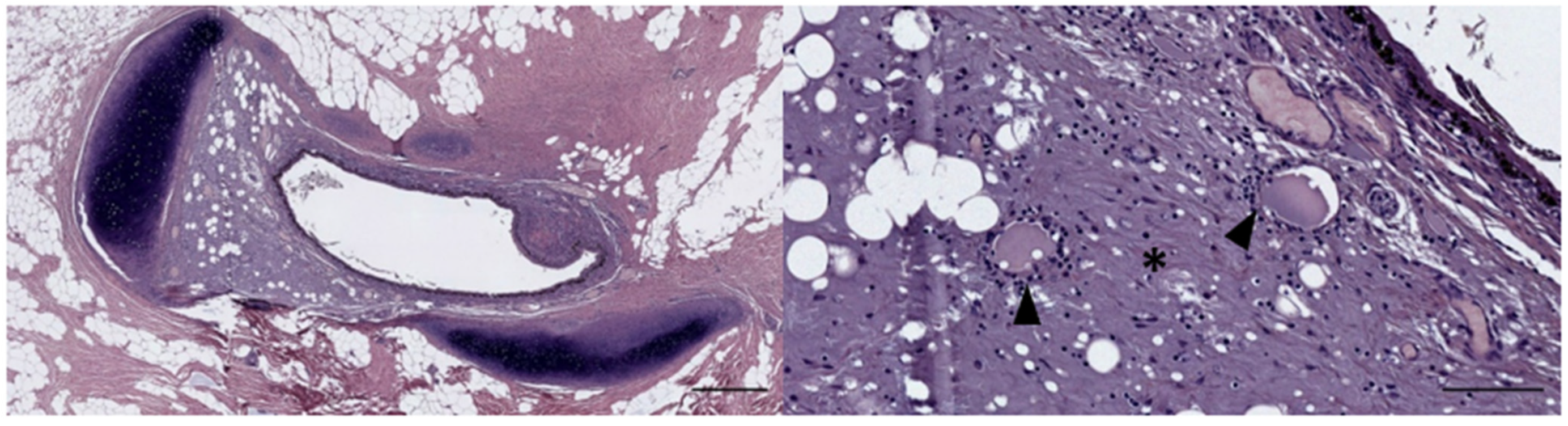
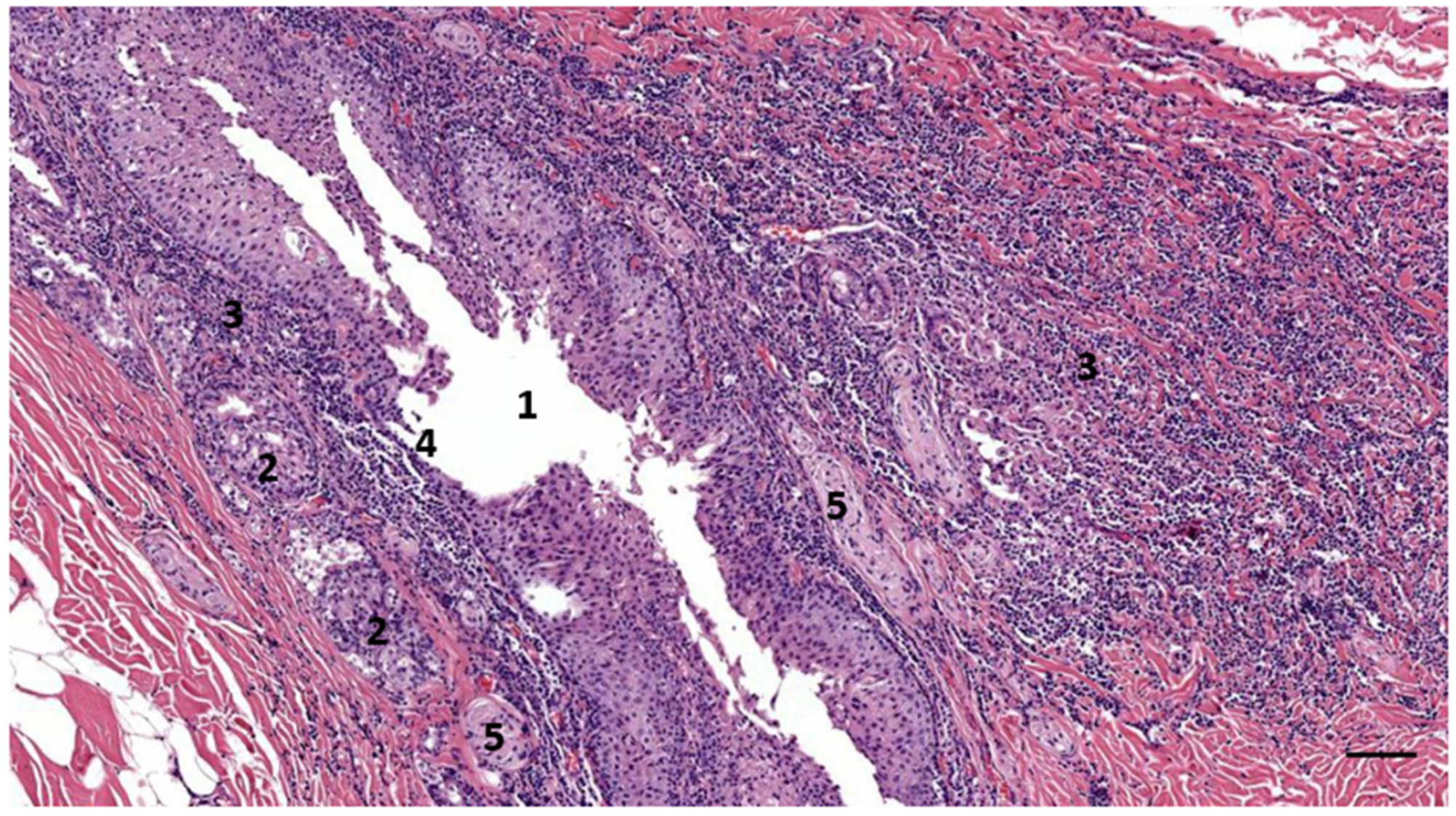
3.1. Ear Canal-Associated Lymphoid Tissue (ECALT)
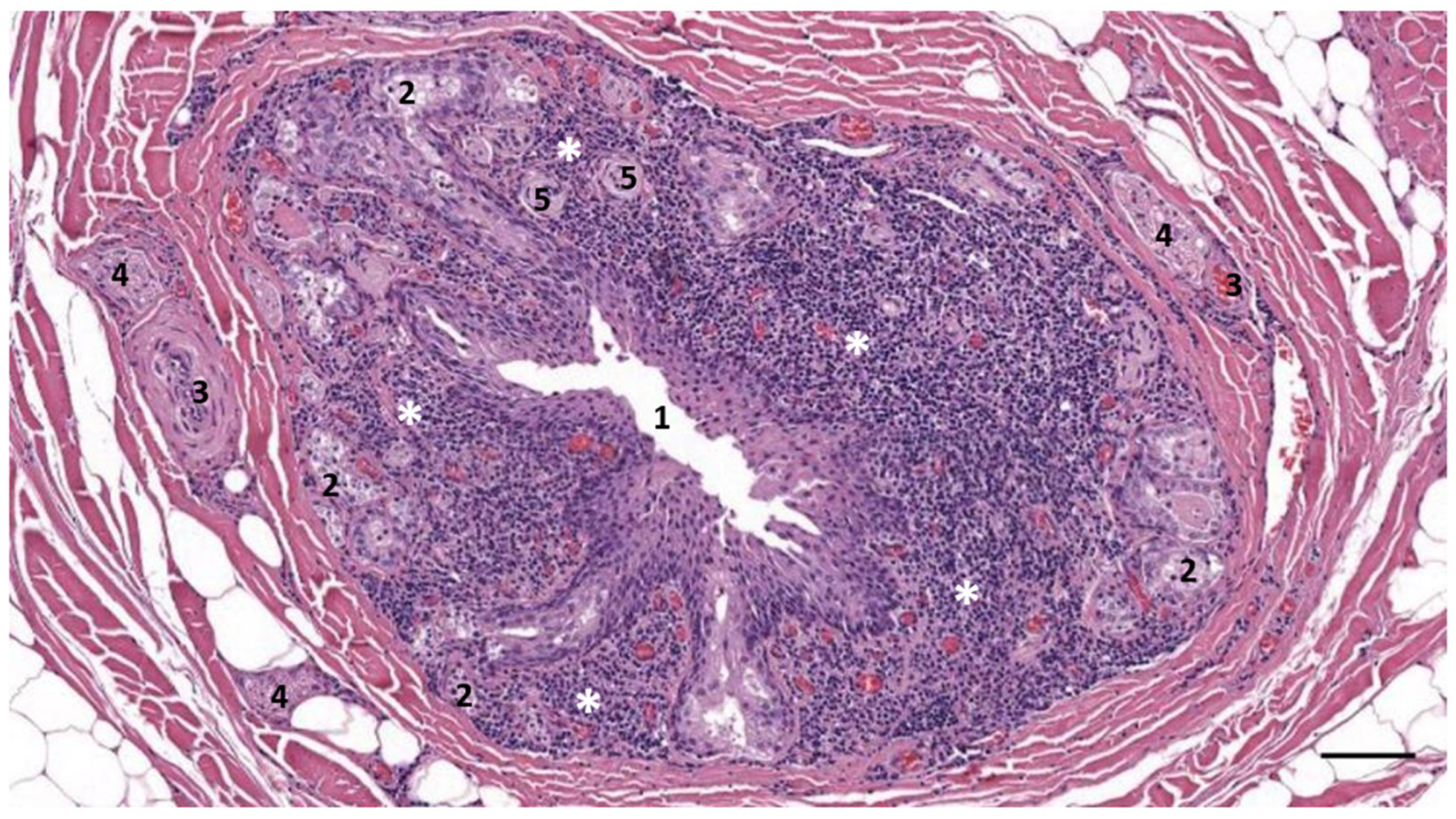
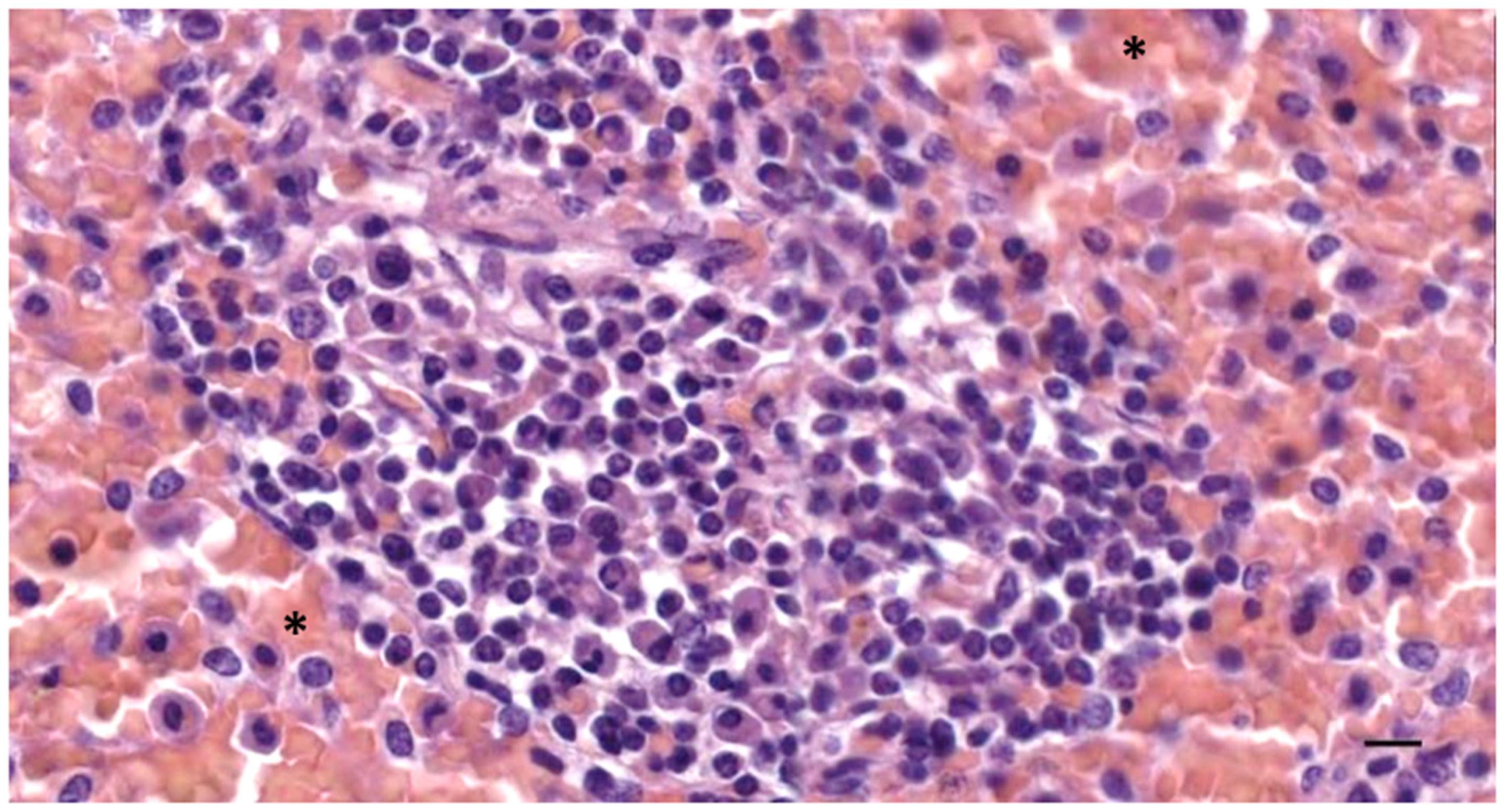
3.2. Nodular Lymphoid Tissue
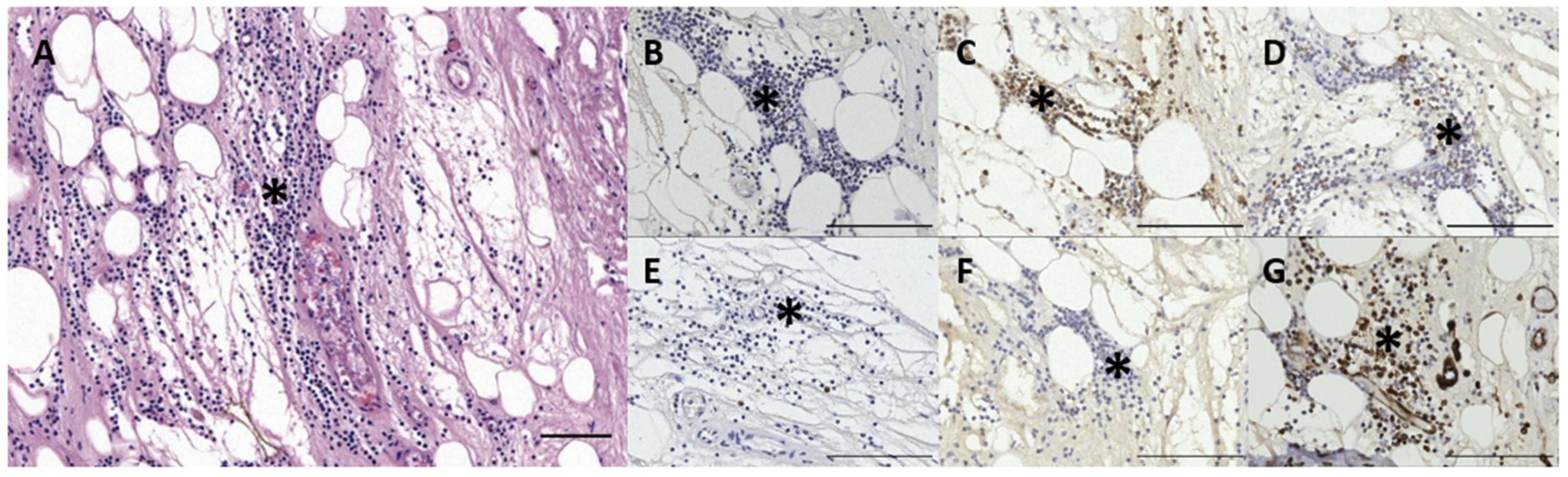
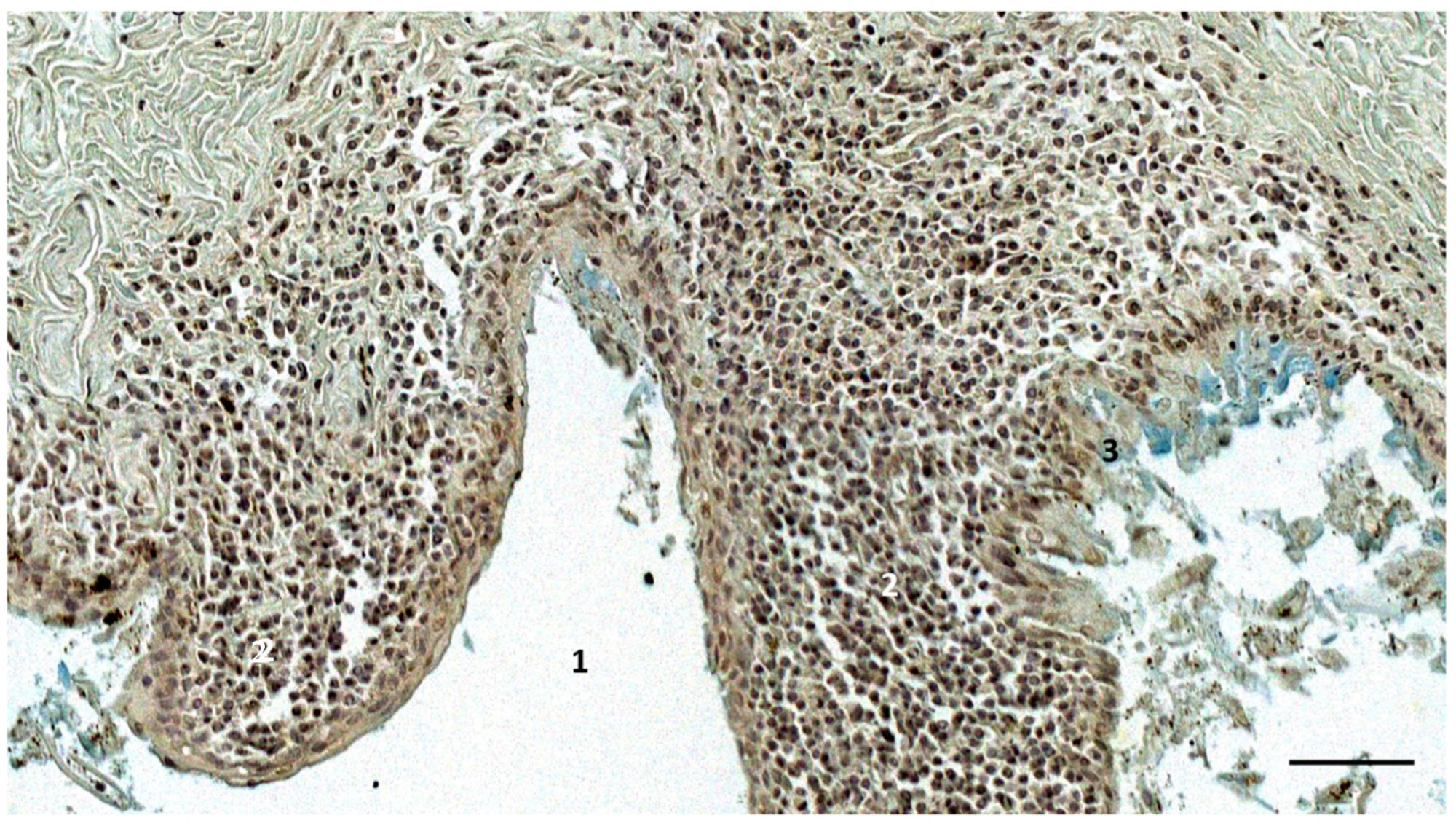
3.3. Immunoreactive Tissue
4. Discussions
4.1. Ear Canal-Associated Lymphoid Tissue
4.2. Ear Canal Lymph Node
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal ID | Species | CC | Origin | Date of Necropsy | Death | Sex | Length (CM) | Weight (KG) | Ear Canal | Lymph Node | Lab Analysis Lymphoid Tissue | |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Left | Right | |||||||||||
| N-00419-16 | S. coeruleoalba | 2 | Port de la Selva, Spain | 16 November 2016 | Spont. | M | 103 | 14 | Y | N | N | HC |
| N-00044-17 | S. coeruleoalba | 2 | Port Ginesta, Spain | 10 February 2017 | Spont. | M | 153 | 37 | Y | Y | N | HC |
| N-00168-17 | S. coeruleoalba | 2 | L’Escala, Spain | 19 April 2017 | Spont. | M | 193 | 79 | Y | N | N | HC |
| N-00169-17 | D. delphis | 2 | L’Escala, Spain | 20 April 2017 | Spont. | M | 179 | 72 | Y | Y | N | HC, IHC |
| N-00488-17 | S. coeruleoalba | 2 | Gavà, Spain | 28 September 2017 | Euth. | F | 198 | 70 | Y | Y | N | HC |
| N-00509-17 | S. coeruleoalba | 2-3 | Tarragona, Spain | 10 October 2017 | Spont. | M | 183 | 87 | Y | Y | N | HC |
| 12691 | S. coeruleoalba | 2 | Bibona, Livorno, Italy | 19 December 2017 | Spont. | M | 193,5 | 62.4 | Y | Y | N | HC |
| 12693 | S. coeruleoalba | 3 | Livorno Italy | 19 December 2017 | Spont. | F | 160 | 40 | Y | Y | N | HC |
| 12694/429 | Z. cavirostris | 2 | Livorno Italy | 23 December 2017 | Spont. | M | 503 | na | Y | Y | N | HC, IHC |
| 12703/2926 | S. coeruleoalba | 2 | Bagni Buraxen, Imperia, Italy | 13 January 2018 | Spont. | M | 133 | 34.5 | Y | Y | N | HC |
| N-00021-18 | S. coeruleoalba | 2-3 | Calella de palafrugell, Spain | 15 January 2018 | Spont. | M | 182 | 73 | Y | N | N | HC |
| 12708/5386 | S. coeruleoalba | 2 | Genova Italy | 21 January 2018 | Spont. | M | 192 | 69 | Y | Y | N | HC |
| N-00042-18 | S. coeruleoalba | 2 | Viladecans, Spain | 25 January 2018 | Spont. | M | 178 | 62 | Y | N | N | HC |
| N-00077-18 | S. coeruleoalba | 2 | Delta del Ebro, Spain | 19 February 2018 | Spont. | M | 220 | 97.5 | Y | Y | N | HC, IHC |
| 441 | G. melas | 4 | Aglientu, Sardegna, Italy | 20 March 2018 | Spont. | M | 525 | na | Y | Y | N | HC, IHC |
| 444 | T. truncatus | 3 | Pellestrina, Venezia, Italy | 24 March 2018 | Spont. | M | 264 | 260 | Y | Y | N | HC, IHC |
| N-00145-18 | S. coeruleoalba | 2 | Palamós, Spain | 26 March 2018 | Spont. | F | 195 | 91 | Y | N | N | HC |
| N-00232-18 | S. coeruleoalba | 1 | Tarragona, Spain | 24 May 2018 | Spont. | M | 180 | 51.5 | Y | N | N | HC |
| N-00274-18 | S. coeruleoalba | 3 | Delta del Ebro, Spain | 24 June 2018 | Spont. | M | 152 | 38 | Y | Y | Bilateral | HC, IHC |
| N-00292-18 | S. coeruleoalba | 2 | Tarragona, Spain | 6 July 2018 | Spont. | M | 194 | 59.5 | Y | Y | N | HC |
| N-00293-18 | S. coeruleoalba | 3 | Port Ginesta, Spain | 09/07/2018 | Euth. | F | 187 | 85 | Y | Y | N | HC, IHC |
| N-00620-17 | S. coeruleoalba | 2 | Vilanova i la Geltrú, Spain | 24 July 2018 | Spont. | M | 196 | 75.5 | Y | N | N | HC |
| N-00362-18 | S. coeruleoalba | 2 | Riumar, Spain | 12 September 2018 | Euth. | M | 181 | 78 | Y | Y | Left | HC, IHC |
| 109064/449 | S. coeruleoalba | 2 | Groticelle, Calabria, Italy | 13 October 2018 | Spont. | M | 102 | 12.23 | Y | Y | Right | HC, IHC |
| N-00177-19 | Z. cavirostris | 4 | Tossa de Mar, Spain | 20 April 2019 | Spont. | M | 520 | 1200 | Y | Y | N | HC |
| 127565 | S. coeruleoalba | 2 | Calabria, Italy | 28 November 2018 | Spont. | M | 200 | na | Y | Y | N | HC |
| AE565/457 | T. truncatus | 2 | Bibione, Venezia, Italy | 29 January 2019 | Spont. | M | 285 | 260 | Y | Y | N | HC, IHC |
| UT1664 | P. phocoena | 2 | Ouddorp, the NLs | 9 July 2018 | Spont. | F | 92 | 11 | Y | N | N | HC |
| UT1692 | P. phocoena | 1 | Kats, the NLs | 25 July 2018 | Spont. | M | 90 | 12.5 | Y | N | N | HC |
| UT1709 | P. phocoena | 2 | Oostkapelle, the NLs | 19 September 2018 | Spont. | M | 95 | 10.5 | Y | Y | N | HC, IHC |
| UT1711 | P. phocoena | 2 | Petten, the NLs | 25 September 2018 | Spont. | M | 146 | 37.5 | Y | Y | N | HC |
| UT1712 | P. phocoena | 3 | Dishoek, the NLs | 4 October 2018 | Spont. | M | 124.5 | 30.5 | Y | Y | N | HC |
| UT1718 | P. phocoena | 2 | Vlieland, the NLs | 05 January 2019 | Spont. | M | 113 | 24 | Y | N | N | HC |
| UT1727 | P. phocoena | 2 | Ijmuiden Seport Marina, the NLs | 13 January 2019 | Spont. | F | 141 | 37 | Y | Y | N | HC |
| UT1728 | P. phocoena | 1 | Egmond, the NLs | 17 January 2019 | Spont. | F | 155 | 64.5 | Y | Y | N | HC |
| UT1734 | P. phocoena | 1 | Terschelling, the NLs | 27 January 2019 | Spont. | M | 103 | 24.5 | Y | Y | N | HC |
| UT1740 | P. phocoena | 1 | Terschelling West, the NLs | 27 February 2019 | Spont. | M | 102 | 22.2 | Y | Y | N | HC |
Appendix B



References
- Beineke, A.; Siebert, U.; Wohlsein, P.; Baumgärtner, W. Immunology of Whales and Dolphins. Vet. Immunol. Immunopathol. 2010, 133, 81–94. [Google Scholar] [CrossRef] [PubMed]
- Jepson, P.D.; Deaville, R.; Barber, J.L.; Aguilar, À.; Borrell, A.; Murphy, S.; Barry, J.; Brownlow, A.; Barnett, J.; Berrow, S.; et al. PCB Pollution Continues to Impact Populations of Orcas and Other Dolphins in European Waters. Sci. Rep. 2016, 6, 18573. [Google Scholar] [CrossRef] [PubMed]
- Van Bressem, M.-F.; Duignan, P.J.; Banyard, A.; Barbieri, M.; Colegrove, K.M.; De Guise, S.; Di Guardo, G.; Dobson, A.; Domingo, M.; Fauquier, D.; et al. Cetacean Morbillivirus: Current Knowledge and Future Directions. Viruses 2014, 6, 5145–5181. [Google Scholar] [CrossRef]
- Centelleghe, C.; Da Dalt, L.; Marsili, L.; Zanetti, R.; Fernandez, A.; Arbelo, M.; Sierra, E.; Castagnaro, M.; Di Guardo, G.; Mazzariol, S. Insights Into Dolphins’ Immunology: Immuno-Phenotypic Study on Mediterranean and Atlantic Stranded Cetaceans. Front. Immunol. 2019, 10, 888. [Google Scholar] [CrossRef]
- Díaz-Delgado, J.; Groch, K.R.; Ressio, R.; Riskallah, I.P.J.; Sierra, E.; Sacchini, S.; Quesada-Canales, Ó.; Arbelo, M.; Fernández, A.; Santos-Neto, E.; et al. Comparative Immunopathology of Cetacean Morbillivirus Infection in Free-Ranging Dolphins From Western Mediterranean, Northeast-Central, and Southwestern Atlantic. Front. Immunol. 2019, 10, 485. [Google Scholar] [CrossRef]
- Cowan, D.F.; Smith, T.L. Morphology of the Lymphoid Organs of the Bottlenose Dolphin. Tursiops Truncatus. J. Anat. 1999, 194, 505–517. [Google Scholar] [CrossRef]
- Silva, F.M.D.O.E.; Guimarães, J.P.; Vergara-parente, J.E.; Carvalho, V.L.; De Meirelles, A.C.O.; Marmontel, M.; Ferrão, J.S.P.; Miglino, M.A. Morphological Analysis of Lymph Nodes in Odontocetes From North and Northeast Coast of Brazil. Anat. Rec. 2014, 297, 939–948. [Google Scholar] [CrossRef]
- Cesta, M.F. Normal Structure, Function, and Histology of Mucosa-Associated Lymphoid Tissue. Toxicol. Pathol. 2006, 34, 599–608. [Google Scholar] [CrossRef]
- Romano, T.A.; Felten, S.Y.; Olschowka, J.A.; Felten, D.L. A Microscopic Investigation of the Lymphoid Organs of the Beluga, Delphinapterus leucas. J. Morphol. 1993, 215, 261–287. [Google Scholar] [CrossRef]
- Clark, L.S.; Turner, J.P.; Cowan, D.F. Involution of Lymphoid Organs in Bottlenose Dolphins (Tursiops Truncatus) from the Western Gulf of Mexico: Implications for Life in an Aquatic Environment. Anat. Rec. A Discov. Mol. Cell Evol. Biol. 2005, 282, 67–73. [Google Scholar] [CrossRef]
- Silva, F.M.O.; Guimarães, J.P.; Vergara-Parente, J.E.; Carvalho, V.L.; Carolina, A.; Meirelles, O.; Marmontel, M.; Oliveira, B.S.S.P.; Santos, S.M.; Becegato, E.Z.; et al. Morphology of Mucosa-Associated Lymphoid Tissue in Odontocetes. Microsc. Res. Tech. 2016, 79, 845–855. [Google Scholar] [CrossRef]
- Beineke, A.; Siebert, U.; Wünschmann, A.; Stott, J.L.; Prengel, I.; Kremmer, E.; Baumgärtner, W. Immunohistochemical Investigation of the Cross-Reactivity of Selected Cell Markers from Various Species for Characterization of Lymphatic Tissues in the Harbour Porpoise (Phocoena Phocoena). J. Comp. Pathol. 2001, 125, 311–317. [Google Scholar] [CrossRef] [PubMed]
- Jaber, J.R.; Fernández, A.; Herráez, P.; Espinosa de los Monteros, A.; Ramírez, G.A.; García, P.M.; Fernández, T.; Arbelo, M.; Pérez, J. Cross-Reactivity of Human and Bovine Antibodies in Striped Dolphin Paraffin Wax-Embedded Tissues. Vet. Immunol. Immunopathol. 2003, 96, 65–72. [Google Scholar] [CrossRef]
- Zabka, T.S.; Romano, T.A. Distribution of MHC II (+) Cells in Skin of the Atlantic Bottlenose Dolphin (Tursiops Truncatus): An Initial Investigation of Dolphin Dendritic Cells. Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2003, 273A, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Kawashima, M.; Nakanishi, M.; Kuwamura, M.; Takeya, M.; Yamate, J. Immunohistochemical Detection of Macrophages in the Short-Finned Pilot Whale (Globicephala Macrorhynchus) and Risso’s Dolphin (Grampus Griseus). J. Comp. Pathol. 2004, 130, 32–40. [Google Scholar] [CrossRef]
- De Vreese, S.; André, M.; Mazzariol, S. Morphology of the External Ear Canal in Toothed Whales. Proc. Mtgs. Acoust. 2019, 37, 010016. [Google Scholar] [CrossRef]
- IJsseldijk, L.L.; Brownlow, A.; Mazzariol, S. Best Practice for Cetacean Post Mortem Investigation and Tissue Sampling; ACCOBAMS-MOP7/2019/Doc 33; ASCOBANS: Bonn, Germany, 2019. [Google Scholar]
- Nakagun, S.; Kobyashi, Y. Histochemical and Immunohistochemical Characterizations of Hepatic Trematodiasis in Odontocetes. Front. Vet. Sci. 2020, 7, 336. [Google Scholar] [CrossRef]
- Holt, W.V.; Waller, J.; Moore, A.; Jepson, P.D.; Deaville, R.; Bennett, P.M. Smooth Muscle Actin and Vimentin as Markers of Testis Development in the Harbour Porpoise (Phocoena Phocoena). J. Anat. 2004, 205, 201–211. [Google Scholar] [CrossRef]
- Suárez-Santana, C.M.; Fernández-Maldonado, C.; Díaz-Delgado, J.; Arbelo, M.; Suárez-Bonnet, A.; Espinosa de los Monteros, A.; Câmara, N.; Sierra, E.; Fernández, A. Pulmonary Carcinoma with Metastasis in a Long-Finned Pilot Whale (Globicephala Melas). BMC Vet. Res. 2016, 12, 229. [Google Scholar] [CrossRef]
- Suárez-Santana, C.M.; Fernández, A.; Sierra, E.; Arbelo, M.; Bernaldo de Quirós, Y.; Andrada, M.; Mompeo, B.; Pérez, L.; Blanco, A.; Méndez, A.; et al. Comparative Morphology, Histology, and Cytology of Odontocete Cetaceans Prostates. Anat. Rec. 2020, 303, 2036–2053. [Google Scholar] [CrossRef]
- Mompeó, B.; Pérez, L.; Fernández, A.; Saavedra, P.; Rivero, M.; Arbelo, M.; Arregui, M.; Suárez-Santana, C.; Bernaldo-de-Quiros, Y. Morphological Structure of the Aortic Wall in Three Delphinid Species with Shallow or Intermediate Diving Habits: Evidence for Diving Adaptation. J. Morphol. 2020, 281, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Sierra, E.; Ramírez, T.; Bernaldo de Quirós, Y.; Arregui, M.; Mompeó, B.; Rivero, M.A.; Fernández, A. Pulmonary and Systemic Skeletal Muscle Embolism in a Beaked Whale with a Massive Trauma of Unknown Aetiology. Animals 2022, 12, 508. [Google Scholar] [CrossRef]
- Yamada, M. Contribution to the Anatomy of the Organ of Hearing of Whales. Sci. Rep. Whales Res. Inst. 1953, 8, 79. [Google Scholar]
- Degollada, E. Functional Anatomy and Histology of the Nasal Sac System in Odontocetes (Superfamily Delphinoidea); Autonomous University of Barcelona: Barcelona, Spain, 1998. [Google Scholar]
- Smith, T.L.; Turnbull, B.S.; Cowan, D.F. Morphology of the Complex Laryngeal Gland in the Atlantic Bottlenose Dolphin, Tursiops Truncatus. Anat. Rec. 1999, 254, 98–106. [Google Scholar] [CrossRef]
- Cowan, D.F.; Smith, T.L. Morphology of Complex Lymphoepithelial Organs of the Anal Canal (“Anal Tonsil”) in the Bottlenose Dolphin. Tursiops Truncatus. J. Morphol. 1995, 223, 263–268. [Google Scholar] [CrossRef]
- Ridgway, S.H.; McCormick, J.G.; Wever, E.G. Surgical Approach to the Dolphin’s Ear. J. Exp. Zool. 1974, 188, 265–276. [Google Scholar] [CrossRef] [PubMed]
- Simpson, J.G.; Gardner, M.B. Comparative Microscopic Anatomy of Selected Marine Mammals. In Mammals of the Sea: Biology and Medicine; Ridgway, S.H., Ed.; Charles C. Thomas: Springfield, IL, USA, 1972; pp. 298–418. [Google Scholar]
- Baum, H. The Lymphatic System of the Dog; University of Saskatchewan: Saskatoon, SK, Canada, 1918; p. 219. Available online: https://sites.usask.ca/lymphaticsystemofthedog/wp-content/uploads/sites/417/2022/05/Lymphatic-System-of-the-Dog-Baum.pdf (accessed on 1 July 2022).
- Vukovic, S.; Lucić, H.; Gomercić, H.; Duras Gomercić, M.; Gomercić, T.; Skrtić, D.; Curković, S. Morphology of the Lymph Nodes in Bottlenose Dolphin (Tursiops Truncatus) and Striped Dolphin (Stenella Coeruleoalba) from the Adriatic Sea. Acta Vet. Hung. 2005, 53, 1–11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moskov, M.; Schiwatschewa, T.; Boney, S. Comparative Histological Study of Lymph Nodes in Mammals. Lymph Nodes of the Dolphin. Anat. Anz. 1969, 124, 49–67. [Google Scholar]
- Hogg, C.M.; Reid, O.; Scothorne, R.J. Studies on Hemolymph Nodes. III. Renal Lymph as a Major Source of Erythrocytes in the Renal Hemolymph Node of Rats. J. Anat. 1982, 135, 291–299. [Google Scholar]
- Slijper, E.J. Walvissen (Amsterdam—Hilversum—Den Helder); Den Helder: Amsterdam, The Netherlands, 1958; p. 524. [Google Scholar]














| Antibody | Manufacturer/Code | Target | Cellular Staining | References | Mono/Polyclonal + Host | Antigen Origin | Dilution | Blocking | Positive Control |
|---|---|---|---|---|---|---|---|---|---|
| anti-CD3 * | Dako M7254 | (Mature) T-lymphocytes | Cytoplasm and/or cell membrane | [5,18] | Monoclonal Mouse | Human | 1:50 | No | Canine + bottlenose dolphin lymph node |
| anti-CD20 * | Thermo Scientific #RB-9013-P | Mature B-lymphocytes | Cytoplasm and cell membrane | [4,5,18] | Monoclonal Rabbit | Human | 1:800 | No | |
| anti-Iba-1 * | Wako #019-19741 | Macrophages and microglia | Cell membrane | [5,18] | Polyclonal Rabbit | Human | 1:80 | No | |
| anti-HLA-DR * | Dako M0746 | B-cells, activated T-cells, macrophages, APC * | Cell-surface membrane (also cytoplasm) | [4,5] | Monoclonal mouse | Human | 1:50 | Yes | |
| anti-Vimentin | Dako M0725 | Mesenchymal cells | Cytoplasm | [19,20,21] | Monoclonal mouse | Porcine | 1:150 | No | Internal: interstitial cells |
| anti-Cytokeratin (PanCK *) | Dako M3515 | Epithelium | Cytoplasm | [20] | Monoclonal Mouse | Human | 1:100 | No | Internal: epithelium of external ear canal |
| anti-vWf * | Dako A0082 | Endothelial cells | Cytoplasm | [22,23] | Monoclonal Rabbit | Human | 1:300 | No | Internal: blood vessel endothelium |
| Pathology | Species | Positive | Total Animals | Percentage |
|---|---|---|---|---|
| Otitis externa | Tt | 1 | 2 | 50% |
| Sc | 3 | 21 | 15% | |
| Gm | 1 | 1 | 100% | |
| Zc | 1 | 2 | 50% | |
| Pp | 0 | 10 | 0.00% | |
| TOTAL | 6 | 36 | 16.67% | |
| Haemorrhage | Sc | 3 | 21 | 14.29% |
| Erythrocytes in external ear opening | Sc | 2 | 21 | 9.52% |
| Pp | 1 | 10 | 10.00% | |
| Panniculitis/dermatitis | Sc | 3 | 21 | 14.29% |
| Gm | 1 | 1 | 100.00% | |
| Vasculitis | Sc | 1 | 21 | 4.76% |
| Muscle atrophy/degeneration | Sc | 8 | 21 | 38.10% |
| Tt | 1 | 2 | 50.00% | |
| Pp | 1 | 10 | 10.00% | |
| Peripheral nerve pathology | Sc | 3 | 21 | 14.29% |
| Epithelial cyst/Cholesteatoma/Keratoma | Sc | 2 | 21 | 9.52% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
De Vreese, S.; Centelleghe, C.; Graïc, J.-M.; Corrazola, G.; IJsseldijk, L.L.; André, M.; Mazzariol, S. The Odontocete Ear Canal-Associated Lymphoid Tissue (ECALT) and Lymph Nodes: Morphological and Pathological Description with Immuno-Phenotypic Characterisation. Animals 2022, 12, 2235. https://doi.org/10.3390/ani12172235
De Vreese S, Centelleghe C, Graïc J-M, Corrazola G, IJsseldijk LL, André M, Mazzariol S. The Odontocete Ear Canal-Associated Lymphoid Tissue (ECALT) and Lymph Nodes: Morphological and Pathological Description with Immuno-Phenotypic Characterisation. Animals. 2022; 12(17):2235. https://doi.org/10.3390/ani12172235
Chicago/Turabian StyleDe Vreese, Steffen, Cinzia Centelleghe, Jean-Marie Graïc, Giorgia Corrazola, Lonneke L. IJsseldijk, Michel André, and Sandro Mazzariol. 2022. "The Odontocete Ear Canal-Associated Lymphoid Tissue (ECALT) and Lymph Nodes: Morphological and Pathological Description with Immuno-Phenotypic Characterisation" Animals 12, no. 17: 2235. https://doi.org/10.3390/ani12172235
APA StyleDe Vreese, S., Centelleghe, C., Graïc, J. -M., Corrazola, G., IJsseldijk, L. L., André, M., & Mazzariol, S. (2022). The Odontocete Ear Canal-Associated Lymphoid Tissue (ECALT) and Lymph Nodes: Morphological and Pathological Description with Immuno-Phenotypic Characterisation. Animals, 12(17), 2235. https://doi.org/10.3390/ani12172235