
First Steps into Ruminal Microbiota Robustness


Abstract
:Simple Summary
Abstract
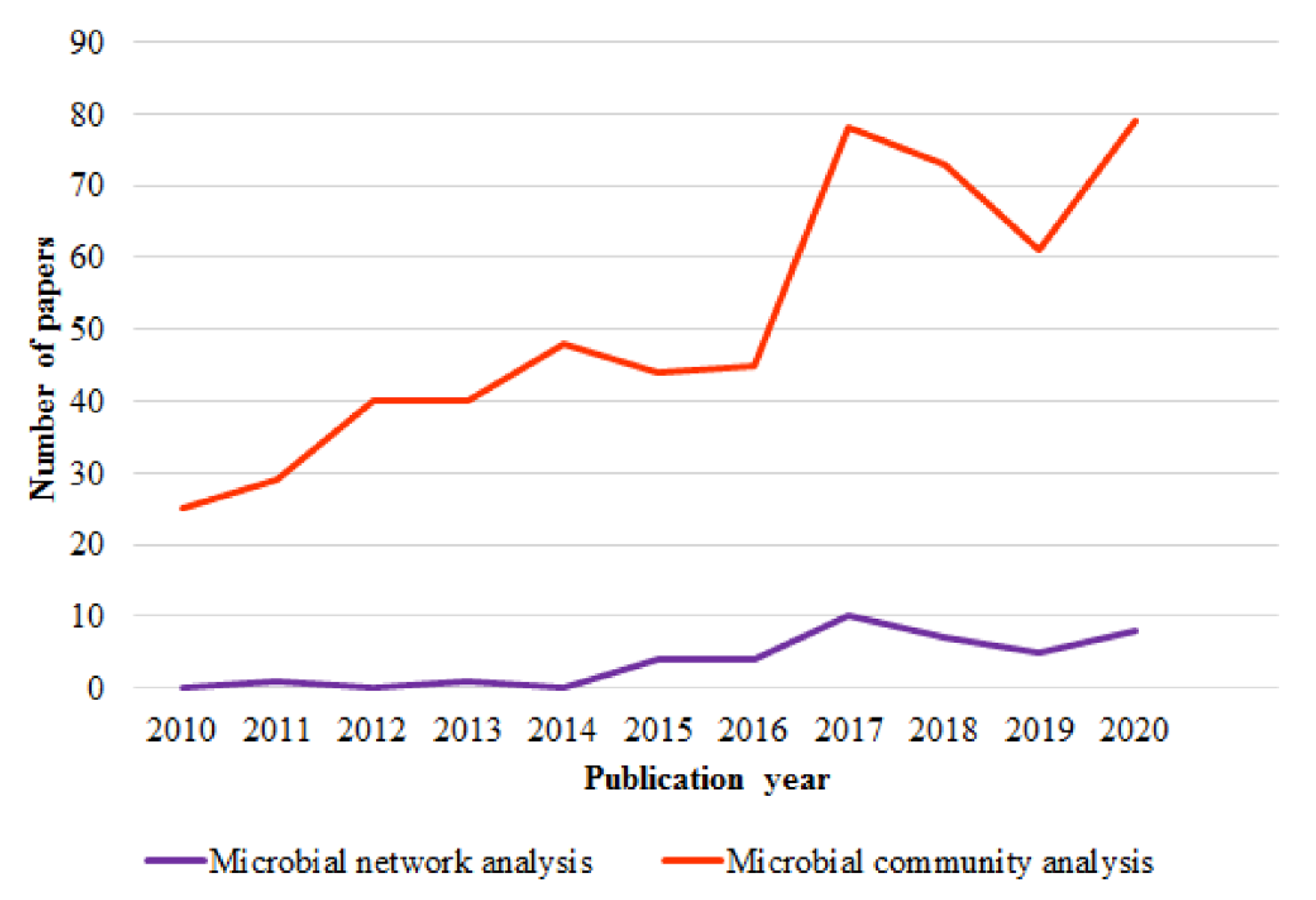
1. Introduction
2. Briefing on the Current Knowledge about Ruminal Microbiota
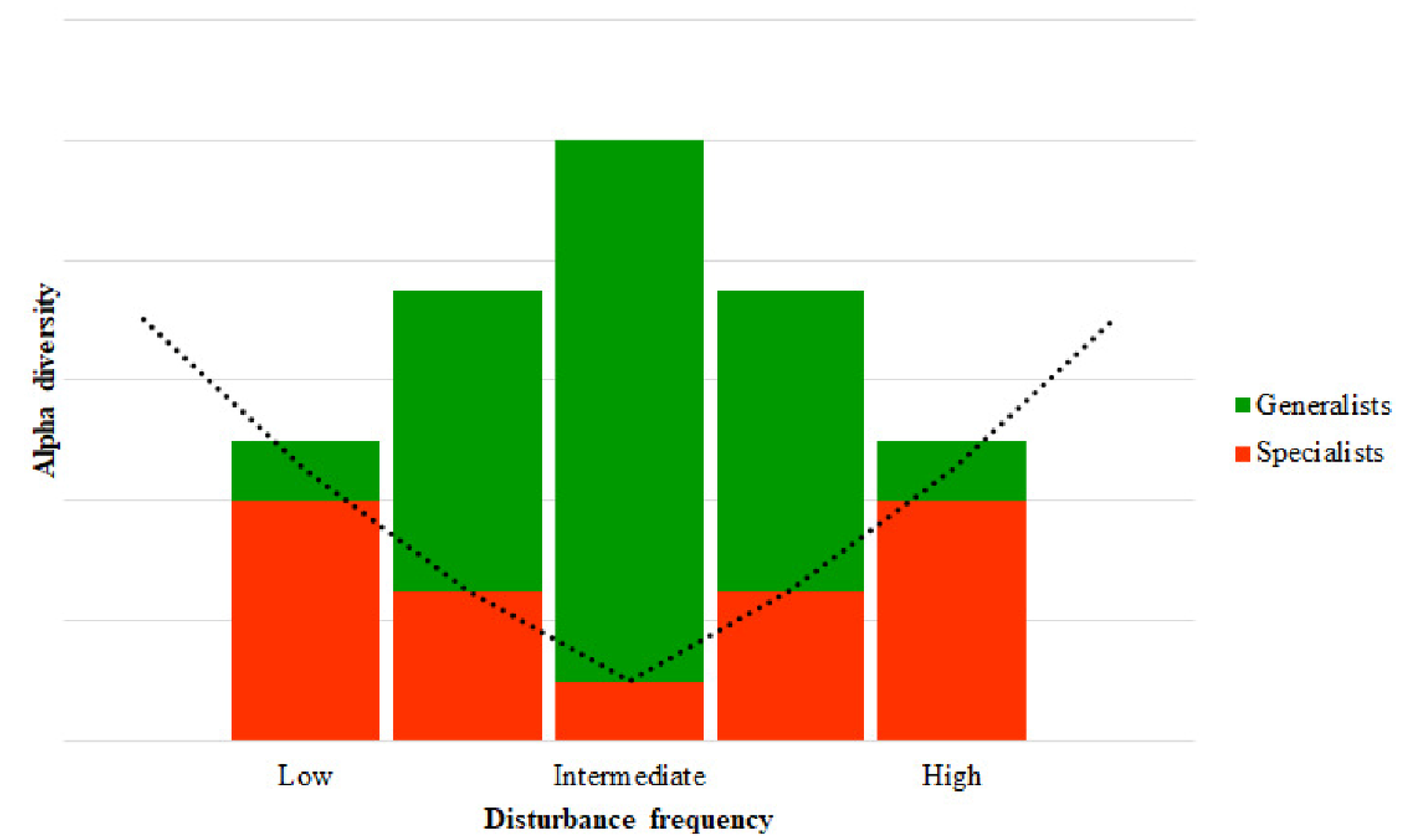
3. Defining Ecosystem Disturbance
4. Defining Microbial Community Robustness
5. Calculating Microbial Community Robustness
6. Drivers of Microbial Community Robustness
6.1. Microbial Alpha Diversity and its Temporal Succession
6.2. Microbial Network Complexity
- The number of nodes is the number of connected taxa within the network;
- The number of edges refers to the number of links established between nodes, and connectance is the number of potential links that are actually realized. An increased number of edges can stabilize the rate of ecosystem processes over time under fluctuating environmental conditions, owing to the ecological redundancy of links, a phenomenon that can also be explained by the previously mentioned insurance hypothesis;
- Nestedness is the tendency of nodes to interact with subsets of the interaction partners of better-connected nodes; in other words, a network is nested when the species interacting with specialists comprise a proper subset of the species interacting with generalists. Nestedness is an important feature of robust communities in that specialists are usually the first species to go extinct from a network; however, if nested, the remaining species will still have generalists to interact with;
- The type of interactions between species plays a role in community robustness. Cooperation between species might facilitate colonization but also create dependency and potential mutual downfall, reducing ecological stability. Although competition may drive inefficiencies, it dampens the destabilizing effects of cooperation, increasing overall stability [102];
- The pattern of interaction strength is also believed to affect community stability; in particular, the presence of many weak links within a network serves to limit energy flow in a potentially strong consumer–resource interaction and, therefore, to inhibit runaway consumption that destabilizes the community dynamics [85];
- Modularity compartmentalizes networks into subsets in which species interact frequently with one another but minimally with other species outside the compartment. Modularity increases community robustness because disturbances spread more slowly through a modular network; therefore, compartmentalized communities will deteriorate more gradually than randomly connected communities [103];
- The node degree is the number of interactions established per node, and its distribution is an important parameter determining community robustness; if the interactions are not evenly distributed across nodes within the network and a few well-connected species concentrate most of the existing links, such a community will be robust to random loss of nodes but very fragile to the elimination of the most connected nodes [104].
6.3. Potential Modulation of Robustness through Diet
7. Future Research: The Link between Diet, Ruminal Microbiota Robustness and Host Health
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hungate, R.E. The Rumen and Its Microbes; Academic Press: New York, NY, USA, 1966. [Google Scholar]
- Kingston-Smith, A.H.; Edwards, J.E.; Huws, S.A.; Kim, E.J.; Abberton, M. Plant-based strategies towards minimising livestock’s long shadow. Proc. Nutr. Soc. 2010, 69, 613–620. [Google Scholar] [CrossRef] [PubMed]
- Weimer, P.J. Redundancy, resilience, and host specificity of the ruminal microbiota: Implications for engineering improved ruminal fermentations. Front. Microbiol. 2015, 6, 296. [Google Scholar] [CrossRef] [PubMed]
- Myer, P.R.; Kim, M.S.; Freetly, H.C.; Smith, T.P.L. Evaluation of 16S rRNA amplicon sequencing using two next-generation sequencing technologies for phylogenetic analysis of the rumen bacterial community in steers. J. Microbiol. Methods 2016, 127, 132–140. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Henderson, G.; Sun, X.; Cox, F.; Janssen, P.H.; Guan, L.L. Taxonomic assessment of rumen microbiota using total RNA and targeted amplicon sequencing approaches. Front. Microbiol. 2016, 7, 987. [Google Scholar] [CrossRef]
- Huws, S.A.; Creevey, C.J.; Oyama, L.B.; Mizrahi, I.; Denman, S.E.; Popova, M.; Muñoz-Tamayo, R.; Forano, E.; Waters, S.M.; Hess, M.; et al. Addressing global ruminant agricultural challenges through understanding the rumen microbiome: Past, present, and future. Front. Microbiol. 2018, 9, 2161. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Ossa, F. The rumen microbiome: Composition, abundance, diversity, and new investigative tools. Prof. Anim. Sci. 2014, 30, e0205670. [Google Scholar] [CrossRef]
- Kim, M.; Morrison, M.; Yu, Z. Status of the phylogenetic diversity census of ruminal microbiomes. FEMS Microbiol. Ecol. 2011, 76, 49–63. [Google Scholar] [CrossRef]
- Morgavi, D.P.; Forano, E.; Martin, C.; Newbold, C.J. Microbial ecosystem and methanogenesis in ruminants. Animal 2010, 4, 1024–1036. [Google Scholar] [CrossRef]
- de La Fuente, G.; Yañez-Ruiz, D.R.; Seradj, A.R.; Balcells, J.; Belanche, A. Methanogenesis in animals with foregut and hindgut fermentation: A review. Anim. Prod. Sci. 2019, 59, 2109–2122. [Google Scholar] [CrossRef]
- Mosoni, P.; Martin, C.; Forano, E.; Morgavi, D.P. Long-term defaunation increases the abundance of cellulolytic ruminococci and methanogens but does not affect the bacterial and methanogen diversity in the rumen of sheep. J. Anim. Sci. 2011, 89, 783–791. [Google Scholar] [CrossRef] [Green Version]
- Newbold, C.J.; De la Fuente, G.; Belanche, A.; Ramos-Morales, E.; McEwan, N.R. The role of ciliate protozoa in the rumen. Front. Microbiol. 2015, 6, 1313. [Google Scholar] [CrossRef]
- Solomon, K.V.; Haitjema, C.H.; Henske, J.K.; Gilmore, S.P.; Borges-Rivera, D.; Lipzen, A.; Brewer, H.M.; Purvine, S.O.; Wright, A.T.; Theodorou, M.K.; et al. Early-branching gut fungi possess large, comprehensive array of biomass-degrading enzymes. Science 2016, 351, 1192–1195. [Google Scholar] [CrossRef]
- Akin, D.E.; Lyon, C.E.; Windham, W.R.; Rigsby, L.L. Physical degradation of lignified stem tissues by ruminal fungi. Appl. Environ. Microbiol. 1989, 55, 611–616. [Google Scholar] [CrossRef]
- Krause, D.O.; Nagaraja, T.G.; Wright, A.D.G.; Callaway, T.R. Board-invited review: Rumen microbiology: Leading the way in microbial ecology. J. Anim. Sci. 2013, 91, 331–341. [Google Scholar] [CrossRef]
- Paul, S.S.; Bu, D.; Xu, J.; Hyde, K.D.; Yu, Z. A phylogenetic census of global diversity of gut anaerobic fungi and a new taxonomic framework. Fungal Divers. 2018, 89, 253–266. [Google Scholar] [CrossRef]
- Adams, J.C.; Gazaway, J.A.; Brailsford, M.D.; Hartman, P.A.; Jacobson, N.L. Isolation of bacteriophages from the bovine rumen. Experientia 1966, 22, 717–718. [Google Scholar] [CrossRef]
- Furman, O.; Shenhav, L.; Sasson, G.; Kokou, F.; Honig, H.; Jacoby, S.; Hertz, T.; Cordero, O.X.; Halperin, E.; Mizrahi, I. Stochasticity constrained by deterministic effects of diet and age drive rumen microbiome assembly dynamics. Nat. Commun. 2020, 11, 1904. [Google Scholar] [CrossRef]
- Jami, E.; Israel, A.; Kotser, A.; Mizrahi, I. Exploring the bovine rumen bacterial community from birth to adulthood. ISME J. 2013, 7, 1069–1079. [Google Scholar] [CrossRef]
- Dill-McFarland, K.A.; Weimer, P.J.; Breaker, J.D.; Suen, G. Diet influences early microbiota development in dairy calves without long-term impacts on milk production. Appl. Environ. Microbiol. 2019, 85, e02141-18. [Google Scholar] [CrossRef]
- Henderson, G.; Cox, F.; Ganesh, S.; Jonker, A.; Young, W.; Collaborators, G.R.C.; Janssen, P.H. Rumen microbial community composition varies with diet and host, but a core microbiome is found across a wide geographical range. Sci. Rep. 2015, 5, srep14567. [Google Scholar] [CrossRef] [Green Version]
- John Wallace, R.; Sasson, G.; Garnsworthy, P.C.; Tapio, I.; Gregson, E.; Bani, P.; Huhtanen, P.; Bayat, A.R.; Strozzi, F.; Biscarini, F.; et al. A heritable subset of the core rumen microbiome dictates dairy cow productivity and emissions. Sci. Adv. 2019, 5, 8391–8394. [Google Scholar] [CrossRef]
- Fraterrigo, J.M.; Rusak, J.A. Disturbance-driven changes in the variability of ecological patterns and processes. Ecol. Lett. 2008, 11, 756–770. [Google Scholar] [CrossRef]
- Svensson, J.R.; Lindegarth, M.; Siccha, M.; Lenz, M.; Molis, M.; Wahl, M.; Pavia, H. Maximum species richness at intermediate frequencies of disturbance: Consistency among levels of productivity. Ecology 2007, 88, 830–838. [Google Scholar] [CrossRef]
- Grime, J.P. Evidence for the existence of three primary strategies in plants and its relevance to ecological and evolutionary theory. Am. Nat. 1977, 111, 1169–1194. [Google Scholar] [CrossRef]
- Sousa, W.P. The role of disturbance in natural communities. Annu. Rev. Ecol. Syst. 1984, 15, 353–391. [Google Scholar] [CrossRef]
- Pickett, S.T.A.; White, P.S. The Ecology of Natural Disturbance as Patch Dynamics; Academic Press: New York, NY, USA, 1985. [Google Scholar]
- Shade, A.; Peter, H.; Allison, S.D.; Baho, D.L.; Berga, M.; Bürgmann, H.; Huber, D.H.; Langenheder, S.; Lennon, J.T.; Martiny, J.B.H.; et al. Fundamentals of microbial community resistance and resilience. Front. Microbiol. 2012, 3, 417. [Google Scholar] [CrossRef]
- Lake, P.S. Disturbance, patchiness, and diversity in streams. J. North Am. Benthol. Soc. 2000, 19, 573–592. [Google Scholar] [CrossRef]
- Connell, J.H. Diversity in tropical rain forests and coral reefs. Science 1978, 199, 1302–1310. [Google Scholar] [CrossRef]
- Bengtsson, J. Disturbance and resilience in soil animal communities. Eur. J. Soil Biol. 2002, 38, 119–125. [Google Scholar] [CrossRef]
- Santillan, E.; Seshan, H.; Constancias, F.; Drautz-Moses, D.I.; Wuertz, S. Frequency of disturbance alters diversity, function, and underlying assembly mechanisms of complex bacterial communities. NPJ Biofilms Microbiomes 2019, 5, 8. [Google Scholar] [CrossRef] [Green Version]
- Sriswasdi, S.; Yang, C.C.; Iwasaki, W. Generalist species drive microbial dispersion and evolution. Nat. Commun. 2017, 8, 1162. [Google Scholar] [CrossRef] [PubMed]
- Thomas, M.; Webb, M.; Ghimire, S.; Blair, A.; Olson, K.; Fenske, G.J.; Fonder, A.T.; Christopher-Hennings, J.; Brake, D.; Scaria, J. Metagenomic characterization of the effect of feed additives on the gut microbiome and antibiotic resistome of feedlot cattle. Sci. Rep. 2017, 7, 12257. [Google Scholar] [CrossRef] [PubMed]
- Schären, M.; Drong, C.; Kiri, K.; Riede, S.; Gardener, M.; Meyer, U.; Hummel, J.; Urich, T.; Breves, G.; Dänicke, S. Differential effects of monensin and a blend of essential oils on rumen microbiota composition of transition dairy cows. J. Dairy Sci. 2017, 100, 2765–2783. [Google Scholar] [CrossRef] [PubMed]
- Pitta, D.W.; Pinchak, W.E.; Indugu, N.; Vecchiarelli, B.; Sinha, R.; Fulford, J.D. Metagenomic analysis of the rumen microbiome of steers with wheat-induced frothy bloat. Front. Microbiol. 2016, 7, 689. [Google Scholar] [CrossRef]
- Azad, E.; Derakhshani, H.; Forster, R.J.; Gruninger, R.J.; Acharya, S.; McAllister, T.A.; Khafipour, E. Characterization of the rumen and fecal microbiome in bloated and non-bloated cattle grazing alfalfa pastures and subjected to bloat prevention strategies. Sci. Rep. 2019, 9, 4272. [Google Scholar] [CrossRef]
- Hook, S.E.; Steele, M.A.; Northwood, K.S.; Dijkstra, J.; France, J.; Wright, A.D.G.; Mcbride, B.W. Impact of subacute ruminal acidosis (SARA) adaptation and recovery on the density and diversity of bacteria in the rumen of dairy cows. FEMS Microbiol. Ecol. 2011, 78, 275–284. [Google Scholar] [CrossRef]
- Mao, S.Y.; Zhang, R.Y.; Wang, D.S.; Zhu, W.Y. Impact of subacute ruminal acidosis (SARA) adaptation on rumen microbiota in dairy cattle using pyrosequencing. Anaerobe 2013, 24, 12–19. [Google Scholar] [CrossRef]
- Díaz Carrasco, J.M.; Cabral, C.; Redondo, L.M.; Pin Viso, N.D.; Colombatto, D.; Farber, M.D.; Fernández Miyakawa, M.E. Impact of chestnut and quebracho tannins on rumen microbiota of bovines. BioMed Res. Int. 2017, 2017, 9610810. [Google Scholar] [CrossRef]
- Zhou, K.; Bao, Y.; Zhao, G. Effects of dietary crude protein and tannic acid on rumen fermentation, rumen microbiota and nutrient digestion in beef cattle. Arch. Anim. Nutr. 2019, 73, 30–43. [Google Scholar] [CrossRef]
- Patra, A.K.; Park, T.; Braun, H.S.; Geiger, S.; Pieper, R.; Yu, Z.; Aschenbach, J.R. Dietary bioactive lipid compounds rich in menthol alter interactions among members of ruminal microbiota in sheep. Front. Microbiol. 2019, 10, 2038. [Google Scholar] [CrossRef]
- Wang, B.; Ma, M.P.; Diao, Q.Y.; Tu, Y. Saponin-induced shifts in the rumen microbiome and metabolome of young cattle. Front. Microbiol. 2019, 10, 356. [Google Scholar] [CrossRef]
- Popova, M.; Guyader, J.; Silberberg, M.; Seradj, A.R.; Saro, C.; Bernard, A.; Gérard, C.; Martin, C.; Morgavi, D.P. Changes in the rumen microbiota of cows in response to dietary supplementation with nitrate, linseed, and saponin alone or in combination. Appl. Environ. Microbiol. 2019, 85, e02657-18. [Google Scholar] [CrossRef]
- Azad, E.; Narvaez, N.; Derakhshani, H.; Alazzeh, A.Y.; Wang, Y.; McAllister, T.A.; Khafipour, E. Effect of Propionibacterium acidipropionici P169 on the rumen and faecal microbiota of beef cattle fed a maize-based finishing diet. Benef. Microbes 2017, 8, 785–799. [Google Scholar] [CrossRef]
- Chaucheyras-Durand, F.; Ameilbonne, A.; Auffret, P.; Bernard, M.; Mialon, M.M.; Dunière, L.; Forano, E. Supplementation of live yeast based feed additive in early life promotes rumen microbial colonization and fibrolytic potential in lambs. Sci. Rep. 2019, 9, 19216. [Google Scholar] [CrossRef]
- Ogunade, I.; Schweickart, H.; McCoun, M.; Cannon, K.; McManus, C. Integrating 16s rRNA sequencing and LC–MS-based metabolomics to evaluate the effects of live yeast on rumen function in beef cattle. Animals 2019, 9, 28. [Google Scholar] [CrossRef]
- Zhang, N.; Wang, L.; Wei, Y. Effects of Bacillus amyloliquefaciens and Bacillus pumilus on rumen and intestine morphology and microbiota in weanling Jintang Black goat. Animals 2020, 10, 1604. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, G.; Liu, Z.; Wu, P.; Yu, Z.; Wang, J. Repeated inoculation with fresh rumen fluid before or during weaning modulates the microbiota composition and co-occurrence of the rumen and colon of lambs. BMC Microbiol. 2020, 20, 29. [Google Scholar] [CrossRef]
- Santos, B.P.; Bessegatto, J.A.; Alfieri, A.A.; Lisbôa, J.A.N.; Weese, J.S.; Costa, M.C. Transfaunation of the ruminal fluid from cows alters ruminal microbiota structure but not dominant protozoa in healthy sheep. Small Rumin. Res. 2021, 194, 106283. [Google Scholar] [CrossRef]
- Weimer, P.J.; Cox, M.S.; Vieira de Paula, T.; Lin, M.; Hall, M.B.; Suen, G. Transient changes in milk production efficiency and bacterial community composition resulting from near-total exchange of ruminal contents between high- and low-efficiency holstein cows. J. Dairy Sci. 2017, 100, 7165–7182. [Google Scholar] [CrossRef]
- Mu, Y.Y.; Qi, W.P.; Zhang, T.; Zhang, J.Y.; Mei, S.J.; Mao, S.Y. Changes in rumen fermentation and bacterial community in lactating dairy cows with subacute rumen acidosis following rumen content transplantation. J. Dairy Sci. 2021, 104, 10780–10795. [Google Scholar] [CrossRef]
- Levin, S.A.; Lubchenco, J. Resilience, robustness, and marine ecosystem-based management. BioScience 2008, 58, 27–32. [Google Scholar] [CrossRef]
- Moya, A.; Ferrer, M. Functional redundancy-induced stability of gut microbiota subjected to disturbance. Trends Microbiol. 2016, 24, 402–413. [Google Scholar] [CrossRef]
- Allison, S.D.; Martiny, J.B.H. Resistance, resilience, and redundancy in microbial communities. Light Evol. 2009, 2, 149–166. [Google Scholar] [CrossRef]
- de la Guardia-Hidrogo, V.M.; Paz, H.A. Influence of industry standard feeding frequencies on behavioral patterns and rumen and fecal bacterial communities in holstein and jersey cows. PLoS ONE 2021, 16, e0248147. [Google Scholar] [CrossRef]
- Li, R.W.; Wu, S.; Baldwin VI, R.L.; Li, W.; Li, C. Perturbation dynamics of the rumen microbiota in response to exogenous butyrate. PLoS ONE 2012, 7, e29392. [Google Scholar] [CrossRef]
- Petri, R.M.; Schwaiger, T.; Penner, G.B.; Beauchemin, K.A.; Forster, R.J.; McKinnon, J.J.; McAllister, T.A. Changes in the rumen epimural bacterial diversity of beef cattle as affected by diet and induced ruminal acidosis. Appl. Environ. Microbiol. 2013, 79, 3744–3755. [Google Scholar] [CrossRef]
- Taxis, T.M.; Wolff, S.; Gregg, S.J.; Minton, N.O.; Zhang, C.; Dai, J.; Schnabel, R.D.; Taylor, J.F.; Kerley, M.S.; Pires, J.C.; et al. The players may change but the game remains: Network analyses of ruminal microbiomes suggest taxonomic differences mask functional similarity. Nucleic Acids Res. 2015, 43, 9600–9612. [Google Scholar] [CrossRef]
- Söllinger, A.; Tveit, A.T.; Poulsen, M.; Noel, S.J.; Bengtsson, M.; Bernhardt, J.; Frydendahl Hellwing, A.L.; Lund, P.; Riedel, K.; Schleper, C.; et al. Holistic assessment of rumen microbiome dynamics through quantitative metatranscriptomics reveals multifunctional redundancy during key steps of anaerobic feed degradation. mSystems 2018, 3, e00038-18. [Google Scholar] [CrossRef]
- Holling, C.S. Resilience and stability of ecological systems. Annu. Rev. Ecol. Syst. 1973, 4, 1–23. [Google Scholar] [CrossRef]
- Beisner, B.E.; Haydon, D.T.; Cuddington, K. Alternative stable states in ecology. Front. Ecol. Environ. 2003, 1, 376–382. [Google Scholar] [CrossRef]
- Arumugam, M.; Raes, J.; Pelletier, E.; Le Paslier, D.; Yamada, T.; Mende, D.R.; Fernandes, G.R.; Tap, J.; Bruls, T.; Batto, J.M.; et al. Enterotypes of the human gut microbiome. Nature 2011, 473, 174–180. [Google Scholar] [CrossRef] [PubMed]
- Kittelmann, S.; Pinares-Patiño, C.S.; Seedorf, H.; Kirk, M.R.; Ganesh, S.; McEwan, J.C.; Janssen, P.H. Two different bacterial community types are linked with the low-methane emission trait in sheep. PLoS ONE 2014, 9, e103171. [Google Scholar] [CrossRef] [PubMed]
- Shaani, Y.; Zehavi, T.; Eyal, S.; Miron, J.; Mizrahi, I. Microbiome niche modification drives diurnal rumen community assembly, overpowering individual variability and diet effects. ISME J. 2018, 12, 2446–2457. [Google Scholar] [CrossRef] [PubMed]
- Abecia, L.; Ramos-Morales, E.; Martínez-Fernandez, G.; Arco, A.; Martín-García, A.I.; Newbold, C.J.; Yáñez-Ruiz, D.R. Feeding management in early life influences microbial colonisation and fermentation in the rumen of newborn goat kids. Anim. Prod. Sci. 2014, 54, 1449–1454. [Google Scholar] [CrossRef]
- Tapio, I.; Fischer, D.; Blasco, L.; Tapio, M.; Wallace, R.J.; Bayat, A.R.; Ventto, L.; Kahala, M.; Negussie, E.; Shingfield, K.J.; et al. Taxon abundance, diversity, co-occurrence and network analysis of the ruminal microbiota in response to dietary changes in dairy cows. PLoS ONE 2017, 12, e0180260. [Google Scholar] [CrossRef]
- Li, F.; Li, C.; Chen, Y.; Liu, J.; Zhang, C.; Irving, B.; Fitzsimmons, C.; Plastow, G.; Guan, L.L. Host genetics influence the rumen microbiota and heritable rumen microbial features associate with feed efficiency in cattle. Microbiome 2019, 7, 92. [Google Scholar] [CrossRef]
- Moraïs, S.; Mizrahi, I. The Road Not Taken: The rumen microbiome, functional groups, and community states. Trends Microbiol. 2019, 27, 538–549. [Google Scholar] [CrossRef]
- Ramayo-Caldas, Y.; Zingaretti, L.; Popova, M.; Estellé, J.; Bernard, A.; Pons, N.; Bellot, P.; Mach, N.; Rau, A.; Roume, H.; et al. Identification of rumen microbial biomarkers linked to methane emission in holstein dairy cows. J. Anim. Breed. Genet. 2020, 137, 49–59. [Google Scholar] [CrossRef]
- Bier, R.L.; Bernhardt, E.S.; Boot, C.M.; Graham, E.B.; Hall, E.K.; Lennon, J.T.; Nemergut, D.R.; Osborne, B.B.; Ruiz-González, C.; Schimel, J.P.; et al. Linking microbial community structure and microbial processes: An empirical and conceptual overview. FEMS Microbiol. Ecol. 2015, 91, fiv113. [Google Scholar] [CrossRef]
- Macgillivray, C.W.; Grime, J.P. Testing predictions of the resistance and resilience of vegetation subjected to extreme events. Funct. Ecol. 1995, 9, 640–649. [Google Scholar] [CrossRef]
- Griffiths, B.S.; Ritz, K.; Bardgett, R.D.; Cook, R.; Christensen, S.; Ekelund, F.; Sørensen, S.J.; Bååth, E.; Bloem, J.; de Ruiter, P.C.; et al. Ecosystem response of pasture soil communities to fumigation-induced microbial diversity reductions: An examination of the biodiversity-ecosystem function relationship. OIKOS 2000, 90, 279–294. [Google Scholar] [CrossRef]
- Orwin, K.H.; Wardle, D.A. New indices for quantifying the resistance and resilience of soil biota to exogenous disturbances. Soil Biol. Biochem. 2004, 36, 1907–1912. [Google Scholar] [CrossRef]
- Banning, N.C.; Murphy, D.V. Effect of heat-induced disturbance on microbial biomass and activity in forest soil and the relationship between disturbance effects and microbial community structure. Appl. Soil Ecol. 2008, 40, 109–119. [Google Scholar] [CrossRef]
- van Ruijven, J.; Berendse, F. Diversity enhances community recovery, but not resistance, after drought. J. Ecol. 2010, 98, 81–86. [Google Scholar] [CrossRef]
- Lloret, F.; Keeling, E.G.; Sala, A. Components of tree resilience: Effects of successive low-growth episodes in old ponderosa pine forests. Oikos 2011, 120, 1909–1920. [Google Scholar] [CrossRef]
- Isbell, F.; Craven, D.; Connolly, J.; Loreau, M.; Schmid, B.; Beierkuhnlein, C.; Bezemer, T.M.; Bonin, C.; Bruelheide, H.; de Luca, E.; et al. biodiversity increases the resistance of ecosystem productivity to climate extremes. Nature 2015, 526, 574–577. [Google Scholar] [CrossRef]
- Nimmo, D.G.; Mac Nally, R.; Cunningham, S.C.; Haslem, A.; Bennett, A.F. Vive la résistance: Reviving resistance for 21st century conservation. Trends Ecol. Evol. 2015, 30, 516–523. [Google Scholar] [CrossRef]
- MacArthur, R. Fluctuations of animal populations and a measure of community stability. Ecology 1955, 36, 533–536. [Google Scholar] [CrossRef]
- Yachi, S.; Loreau, M. Biodiversity and ecosystem productivity in a fluctuating environment: The insurance hypothesis. Proc. Natl. Acad. Sci. USA 1999, 96, 1463–1468. [Google Scholar] [CrossRef]
- Tilman, D.; Downing, J.A. Biodiversity and stability in grasslands. Nature 1994, 367, 363–365. [Google Scholar] [CrossRef]
- Van Elsas, J.D.; Chiurazzi, M.; Mallon, C.A.; Elhottova, D.; Krištůfek, V.; Salles, J.F. Microbial diversity determines the invasion of soil by a bacterial pathogen. Proc. Natl. Acad. Sci. USA 2012, 109, 1159–1164. [Google Scholar] [CrossRef]
- Wittebolle, L.; Marzorati, M.; Clement, L.; Balloi, A.; Daffonchio, D.; Heylen, K.; De Vos, P.; Verstraete, W.; Boon, N. Initial community evenness favours functionality under selective stress. Nature 2009, 458, 623–626. [Google Scholar] [CrossRef]
- McCann, K.S. The diversity-stability debate. Nature 2000, 405, 228–233. [Google Scholar] [CrossRef]
- Preston, F.W. Time and space and the variation of species. Ecology 1960, 41, 611–627. [Google Scholar] [CrossRef]
- Adler, P.B.; Lauenroth, W.K. The power of time: Spatiotemporal scaling of species diversity. Ecol. Lett. 2003, 6, 749–756. [Google Scholar] [CrossRef]
- Korhonen, J.J.; Soininen, J.; Hillebrand, H. A quantitative analysis of temporal turnover in aquatic species assemblages across ecosystems. Ecology 2010, 91, 508–517. [Google Scholar] [CrossRef]
- Swenson, N.G.; Stegen, J.C.; Davies, S.J.; Erickson, D.L.; Forero-Montaña, J.; Hurlbert, A.H.; Kress, W.J.; Thompson, J.; Uriarte, M.; Wright, S.J.; et al. Temporal turnover in the composition of tropical tree communities: Functional determinism and phylogenetic stochasticity. Ecology 2012, 93, 490–499. [Google Scholar] [CrossRef]
- Shade, A.; Gregory Caporaso, J.; Handelsman, J.; Knight, R.; Fierer, N. A meta-analysis of changes in bacterial and archaeal communities with time. ISME J. 2013, 7, 1493–1506. [Google Scholar] [CrossRef]
- Koenig, J.E.; Spor, A.; Scalfone, N.; Fricker, A.D.; Stombaugh, J.; Knight, R.; Angenent, L.T.; Ley, R.E. Succession of microbial consortia in the developing infant gut microbiome. Proc. Natl. Acad. Sci. USA 2011, 108, 4578–4585. [Google Scholar] [CrossRef]
- Xiong, J.; Xuan, L.; Yu, W.; Zhu, J.; Qiu, Q.; Chen, J. Spatiotemporal successions of shrimp gut microbial colonization: High consistency despite distinct species pool. Environ. Microbiol. 2019, 21, 1383–1394. [Google Scholar] [CrossRef]
- Anderson, K.E.; Ricigliano, V.A.; Mott, B.M.; Copeland, D.C.; Floyd, A.S.; Maes, P. The queen’s gut refines with age: Longevity phenotypes in a social insect model. Microbiome 2018, 6, 108. [Google Scholar] [CrossRef] [PubMed]
- Karimi, B.; Maron, P.A.; Chemidlin-Prevost Boure, N.; Bernard, N.; Gilbert, D.; Ranjard, L. Microbial diversity and ecological networks as indicators of environmental quality. Environ. Chem. Lett. 2017, 15, 265–281. [Google Scholar] [CrossRef]
- Valiente-Banuet, A.; Aizen, M.A.; Alcántara, J.M.; Arroyo, J.; Cocucci, A.; Galetti, M.; García, M.B.; García, D.; Gómez, J.M.; Jordano, P.; et al. Beyond species loss: The extinction of ecological interactions in a changing world. Funct. Ecol. 2015, 29, 299–307. [Google Scholar] [CrossRef]
- Landi, P.; Minoarivelo, H.O.; Brännström, Å.; Hui, C.; Dieckmann, U. Complexity and Stability of Ecological Networks: A Review of the Theory. Popul. Ecol. 2018, 60, 319–345. [Google Scholar] [CrossRef]
- Heleno, R.; Devoto, M.; Pocock, M. Connectance of species interaction networks and conservation value: Is it any good to be well connected? Ecol. Indic. 2012, 14, 7–10. [Google Scholar] [CrossRef]
- Little, A.E.F.; Robinson, C.J.; Peterson, S.B.; Raffa, K.F.; Handelsman, J. Rules of engagement: Interspecies interactions that regulate microbial communities. Annu. Rev. Microbiol. 2008, 62, 375–401. [Google Scholar] [CrossRef]
- Zappelini, C.; Karimi, B.; Foulon, J.; Lacercat-Didier, L.; Maillard, F.; Valot, B.; Blaudez, D.; Cazaux, D.; Gilbert, D.; Yergeau, E.; et al. Diversity and complexity of microbial communities from a chlor-alkali tailings dump. Soil Biol. Biochem. 2015, 90, 101–110. [Google Scholar] [CrossRef]
- Karimi, B.; Meyer, C.; Gilbert, D.; Bernard, N. Air pollution below WHO levels decreases by 40% the links of terrestrial microbial networks. Environ. Chem. Lett. 2016, 14, 467–475. [Google Scholar] [CrossRef]
- Tylianakis, J.M.; Laliberté, E.; Nielsen, A.; Bascompte, J. Conservation of species interaction networks. Biol. Conserv. 2010, 143, 2270–2279. [Google Scholar] [CrossRef]
- Coyte, K.Z.; Schluter, J.; Foster, K.R. The ecology of the microbiome: Networks, competition, and stability. Science 2015, 350, 663–666. [Google Scholar] [CrossRef]
- Wilmers, C.C. Understanding ecosystem robustness. Trends Ecol. Evol. 2007, 22, 504–506. [Google Scholar] [CrossRef]
- Albert, R.; Barabási, A.L. Topology of evolving networks: Local events and universality. Phys. Rev. Lett. 2000, 85, 5234–5237. [Google Scholar] [CrossRef]
- Wang, L.; Li, Y.; Zhang, Y.; Wang, L. The effects of different concentrate-to-forage ratio diets on rumen bacterial microbiota and the structures of Holstein cows during the feeding cycle. Animals 2020, 10, 957. [Google Scholar] [CrossRef]
- Ramos, S.C.; Jeong, C.D.; Mamuad, L.L.; Kim, S.H.; Kang, S.H.; Kim, E.T.; Cho, Y.I.; Lee, S.S.; Lee, S.S. Diet transition from high-forage to high-concentrate alters rumen bacterial community composition, epithelial transcriptomes and ruminal fermentation parameters in dairy cows. Animals 2021, 11, 838. [Google Scholar] [CrossRef]
- Costa-Roura, S.; Balcells, J.; de la Fuente, G.; Mora-Gil, J.; Llanes, N.; Villalba, D. Nutrient utilization efficiency, ruminal fermentation and microbial community in Holstein bulls fed concentrate-based diets with different forage source. Anim. Feed. Sci. Technol. 2020, 269, 114662. [Google Scholar] [CrossRef]
- Zhang, J.; Shi, H.; Wang, Y.; Li, S.; Cao, Z.; Ji, S.; He, Y.; Zhang, H. Effect of dietary forage to concentrate ratios on dynamic profile changes and interactions of ruminal microbiota and metabolites in holstein heifers. Front. Microbiol. 2017, 8, 2206. [Google Scholar] [CrossRef]
- Belanche, A.; Kingston-Smith, A.H.; Griffith, G.W.; Newbold, C.J. A multi-kingdom study reveals the plasticity of the rumen microbiota in response to a shift from non-grazing to grazing diets in sheep. Front. Microbiol. 2019, 10, 122. [Google Scholar] [CrossRef]
- Liu, H.; Xu, T.; Xu, S.; Ma, L.; Han, X.; Wang, X.; Zhang, X.; Hu, L.; Zhao, N.; Chen, Y.; et al. Effect of dietary concentrate to forage ratio on growth performance, rumen fermentation and bacterial diversity of Tibetan sheep under barn feeding on the Qinghai-Tibetan plateau. PeerJ 2019, 2019, e7462. [Google Scholar] [CrossRef]
- Zhang, R.Y.; Liu, Y.J.; Yin, Y.Y.; Jin, W.; Mao, S.Y.; Liu, J.H. Response of rumen microbiota, and metabolic profiles of rumen fluid, liver and serum of goats to high-grain diets. Animal 2019, 13, 1855–1864. [Google Scholar] [CrossRef]
- Qian, W.; Ao, W.; Hui, X.; Wu, J. Lower dietary concentrate level increases bacterial diversity in the rumen of Cervus elaphus yarkandensis. Can. J. Microbiol. 2018, 64, 501–509. [Google Scholar] [CrossRef]
- Ali, A.I.M.; Wassie, S.E.; Korir, D.; Goopy, J.P.; Merbold, L.; Butterbach-Bahl, K.; Dickhoefer, U.; Schlecht, E. Digesta passage and nutrient digestibility in Boran steers at low feed intake levels. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1325–1337. [Google Scholar] [CrossRef]
- Wang, Y.; Cao, P.; Wang, L.; Zhao, Z.; Chen, Y.; Yang, Y. Bacterial community diversity associated with different levels of dietary nutrition in the rumen of sheep. Appl. Microbiol. Biotechnol. 2017, 101, 3717–3728. [Google Scholar] [CrossRef]
- Dufreneix, F.; Faverdin, P.; Peyraud, J.L. Influence of particle size and density on mean retention time in the rumen of dairy cows. J. Dairy Sci. 2019, 102, 3010–3022. [Google Scholar] [CrossRef]
- Clauss, M.; Lechner, I.; Barboza, P.; Collins, W.; Tervoort, T.A.; Südekum, K.H.; Codron, D.; Hummel, J. The effect of size and density on the mean retention time of particles in the reticulorumen of cattle (Bos primigenius f. taurus), muskoxen (Ovibos moschatus) and moose (Alces alces). Br. J. Nutr. 2011, 105, 634–644. [Google Scholar] [CrossRef]
- Ishaq, S.L.; Lachman, M.M.; Wenner, B.A.; Baeza, A.; Butler, M.; Gates, E.; Olivo, S.; Geddes, J.B.; Hatfield, P.; Yeoman, C.J. Pelleted-hay alfalfa feed increases sheep wether weight gain and rumen bacterial richness over loose-hay alfalfa feed. PLoS ONE 2019, 14, e0215797. [Google Scholar] [CrossRef]
- Costa-Roura, S.; Balcells, J.; de la Fuente, G.; Mora-Gil, J.; Llanes, N.; Villalba, D. Effects of protein restriction on performance, ruminal fermentation and microbial community in Holstein bulls fed high-concentrate diets. Anim. Feed. Sci. Technol. 2020, 264, 114479. [Google Scholar] [CrossRef]
- Park, T.; Ma, L.; Ma, Y.; Zhou, X.; Bu, D.; Bu, D.; Yu, Z. Dietary energy sources and levels shift the multi-kingdom microbiota and functions in the rumen of lactating dairy cows. J. Anim. Sci. Biotechnol. 2020, 11, 66. [Google Scholar] [CrossRef]
- Lv, X.; Chai, J.; Diao, Q.; Huang, W.; Zhuang, Y.; Zhang, N. The signature microbiota drive rumen function shifts in goat kids introduced to solid diet regimes. Microorganisms 2019, 7, 516. [Google Scholar] [CrossRef]
- Zeevi, D.; Korem, T.; Godneva, A.; Bar, N.; Kurilshikov, A.; Lotan-Pompan, M.; Weinberger, A.; Fu, J.; Wijmenga, C.; Zhernakova, A.; et al. Structural variation in the gut microbiome associates with host health. Nature 2019, 568, 43–48. [Google Scholar] [CrossRef]
- Durack, J.; Lynch, S.V. The gut microbiome: Relationships with disease and opportunities for therapy. J. Exp. Med. 2019, 216, 20–40. [Google Scholar] [CrossRef] [Green Version]
- Cammack, K.M.; Austin, K.J.; Lamberson, W.R.; Conant, G.C.; Cunningham, H.C. Ruminnat nutrition symposium: Tiny but mighty: The role of the rumen microbes in livestock production. J. Anim. Sci. 2018, 96, 752–770. [Google Scholar] [CrossRef] [PubMed]
- Zeineldin, M.; Barakat, R.; Elolimy, A.; Salem, A.Z.M.; Elghandour, M.M.Y.; Monroy, J.C. Synergetic action between the rumen microbiota and bovine health. Microb. Pathog. 2018, 124, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Malmuthuge, N.; Guan, L.L. Understanding host-microbial interactions in rumen: Searching the best opportunity for microbiota manipulation. J. Anim. Sci. Biotechnol. 2017, 8, 8. [Google Scholar] [CrossRef] [PubMed]
- Dogra, S.K.; Doré, J.; Damak, S. Gut microbiota resilience: Definition, link to health and strategies for intervention. Front. Microbiol. 2020, 11, 2014–2021. [Google Scholar] [CrossRef]
- Fassarella, M.; Blaak, E.E.; Penders, J.; Nauta, A.; Smidt, H.; Zoetendal, E.G. Gut microbiome stability and resilience: Elucidating the response to perturbations in order to modulate gut health. Gut 2020, 595–605. [Google Scholar] [CrossRef]
- Raymond, F.; Ouameur, A.A.; Déraspe, M.; Iqbal, N.; Gingras, H.; Dridi, B.; Leprohon, P.; Plante, P.L.; Giroux, R.; Bérubé, È.; et al. The initial state of the human gut microbiome determines its reshaping by antibiotics. ISME J. 2016, 10, 707–720. [Google Scholar] [CrossRef]
- Mondot, S.; Lepage, P.; Seksik, P.; Allez, M.; Tréton, X.; Bouhnik, Y.; Colombel, J.F.; Leclerc, M.; Pochart, P.; Doré, J.; et al. Structural robustness of the gut mucosal microbiota is associated with Crohn’s disease remission after surgery. Gut 2016, 65, 954–962. [Google Scholar] [CrossRef]
- González, L.A.; Manteca, X.; Calsamiglia, S.; Schwartzkopf-Genswein, K.S.; Ferret, A. Ruminal acidosis in feedlot cattle: Interplay between feed ingredients, rumen function and feeding behavior (a review). Anim. Feed. Sci. Technol. 2012, 172, 66–79. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Disturbance | Animals | Treatment | Effects on Ruminal Microbiota 1 | Reference |
|---|---|---|---|---|
| In-feed antibiotic administration | Crossbred steers (finishing) | NA: non-hormone, non-antibiotic treatment AB: hormone-implanted cattle fed a beta-agonist (ractopamine) and antibiotics (monensin, 478.3 g/ton; tylosin, 96.1 g/ton) during the finishing period | Decreased alpha diversity in AB steers (reduced Shannon H and inverse Simpson indices and richness). Beta diversity analysis showed no differences in microbial community composition between NA and AB steers. At the phylum level, no differences in taxon abundance between NA and AB steers. At the genus level, decreased abundance of Bacteroidetes (Spirosoma, Dyadobacter, Leadbetterella and Zunongwangia) in AB steers. Gram-positive Firmicutes were partially replaced by Gram-negative Negativicutes in AB steers. | [34] |
| In-feed antibiotic administration | Holstein cows (pluriparous) | CTR: non-antibiotic treatment MO: administration of 335 mg monensin/d during the transition period | Decreased alpha diversity in MO cows (reduced values of Shannon index and richness). Beta diversity analysis showed significant differences in microbial community composition between MO and CTR cows. Decreased abundance of 23 bacterial OTUs in MO (mainly belonging to Bacteroidetes and Firmicutes phyla). Increased abundance of 10 bacterial OTUs in MO (belonging to Actinobacteria, Bacteroidetes, Cyanobacteria and Firmicutes phyla). No difference between CTR and MO cows in archaeal abundance (i.e., Methanobrevibacter). | [35] |
| Frothy bloat | Steers | Steers grazed on winter wheat and were visually scored for bloat: BS0: normal; no visible signs of bloat BS2: marked distention of left side of animal; rumen distended toward top of back | Increased abundance of archaeal community and decreased abundance of bacterial community in BS2 steers. At the bacterial phylum level, increased abundance of Firmicutes and Proteobacteria but decreased abundance of Bacteroidetes and Actinobacteria in BS2 steers. Among archaea, increased abundance of Methanobrevibacter but decreased abundance of Methanosphaera, Methanosarcina, Methanocorpusculum, Methanococcus and Methanococcoides in BS2 steers. Decreased number of interactions among both bacteria and archaea in BS2 steers. | [36] |
| Frothy bloat | Angus steers (3–4 years) | PA: pure alfalfa pasture AA: pure alfalfa pasture, but steers treated with detergent AS: mixed alfalfa–sainfoin pasture Steers were visually scored for bloat: NB: non-bloated steers B: slightly to severely bloated steers | Increased alpha diversity in rumen solid fraction of B steers (increased values of Shannon index and richness). No effect of bloat on alpha diversity in rumen liquid fraction. Beta diversity analysis showed that rumen solid fraction microbiota composition differed between B and NB steers. Beta diversity analysis showed that rumen liquid fraction microbiota composition differed between PA-B and AA-NB and tended to differ between PA-B and AS-NB steers. At the genus level, increased abundance of Succinivibrio and Streptococcus but decreased abundance of Fibrobacteres and Ruminococcus in B steers. | [37] |
| Disturbance | Animals | Treatments 1 | Effects on Ruminal Microbiota 1,2 | Reference |
|---|---|---|---|---|
| Subacute ruminal acidosis | Dairy cows (760 kg) | Sampling protocol lasted for 7 weeks: Week 0: baseline. Week 1 and 3: 65% grain diet. Week 4 and 6: chopped hay diet. | Bacterial density in rumen solids increased during weeks 4 and 6 compared to weeks 1 and 3. Alpha diversity decreased in week 3 compared to week 0 (lower values of Shannon index). Beta diversity analysis showed that ruminal microbiota composition in week 3 was different from week 0 and week 6, but week 0 and week 6 did not differ. | [38] |
| Subacute ruminal acidosis | Holstein cows (pluriparous, 460 kg) | Crossover design 2 treatments × 2 periods (21 d): COD: 40% concentrate diet. SAID: 70% concentrate diet. | Decreased alpha diversity in SAID cows (lower values of Shannon index and richness). At phylum level, increased abundance of Firmicutes and Actinobacteria whereas decreased abundance of Bacteroidetes, Lentisphaerae and Proteobacteria in SAID cows. At genus level, increased abundance of Ruminococcus, Atopobium and Bifidobacterium whereas decreased abundance of Prevotella, Treponema, Papillibacter, Anaeroplasma and Acinetobacter in SAID cows. More abundant gram-positive bacteria than gram-negative bacteria in SAID cows. | [39] |
| Tannins | Holstein cows (584 kg) | CTR: no supplementation. TA: 2 g chestnut and quebracho tannins blend/kg DM for 12 d. | Slight effects on alpha diversity (richness tended to decrease in TA cows). Beta diversity analysis showed no differences between TA and CTR microbial composition in rumen. At phylum level, increased abundance of Firmicutes in TA cows. At genus level, increased abundance of Ruminococcus, L7A-E11, Blautia, Anaerofustis, Anaerovibrio whereas decreased abundance of RFN20, Fibrobacter, Treponema and Methanosphaera in TA cows. | [40] |
| Tannins | Simmental steers (350 kg) | CTR: no supplementation. TA: 16.9 g tannic acid/kg DM for 5 d. | Increased alpha diversity in TA steers (higher values of Shannon index). Beta diversity analysis showed no differences between TA and CTR microbial composition in rumen. At phylum level, increased abundance of Tenericutes in TA steers. At genus level, increased abundance of Saccharofermentans in TA steers. | [41] |
| Essential oils | Holstein cows (pluriparous) | CTR: no supplementation. EO: 1 g essential oils blend/d containing thymol, guaiacol, eugenol, vanillin, salicylaldehyde and limonene during the transition period. | No effects on alpha diversity. Beta diversity analysis showed significant differences between EO and CTR microbial composition in rumen. No difference between CTR and EO cows in archaeal or bacterial abundance at any taxonomic level. | [35] |
| Essential oils | Suffolk lambs (121 d, 33 kg) | CTR: no supplementation PBLC-L: 80 mg menthol-rich PBLC/d for 4 weeks PBLC-H: 160 mg menthol-rich PBLC/d for 4 weeks | No effects on alpha diversity. In the rumen solid fraction, increased abundance of Dehalobacteriaceae, Mycoplasmataceae, UG Lachnospiraceae, US Dehalobacterium, US Desulfovibrio but decreased abundance of Christensenellaceae, UG Paraprevotellaceae, Euryarchaeota, US Methanosphaera, US Prevotella, LD1-PB3 and UG LD1-PB3 in PBLC lambs. In the rumen liquid fraction, increased abundance of WCHB1-25, US WCHB1-25, Bacteroidaceae, US BF311 and US YRC22 but decreased abundance of Christensenellaceae, UG Christensenellaceae, Thermoplasmata, Methanomassiliiococcaceae, vadinCA11, US Blautia 2 and UC Proteobacteria in PBLC lambs. Reduced microbial network complexity (fewer edges, nodes and unique interactions) in PBLC lambs. | [42] |
| Saponins | Holstein bulls (150 d, 150 kg) | AH: concentrate plus alfalfa hay AHS: AH plus 9 g camellia seed saponins/d for 4 weeks SH: concentrate plus soybean hulls SHS: SH plus 9 g camellia seed saponins/d for 4 weeks | No effects on alpha diversity in AH and AHS. Increased alpha diversity (higher values of richness) in SHS bulls compared to SH. Beta diversity analysis showed differences in microbial composition between the four treatments. No difference between AH and AHS bulls in microbial abundance at any taxonomic level. Increased abundance of Prevotella 1, Christensenellaceae R-7, Prevotellaceae Ga6A1, Clostridium sensu stricto 1, Ruminococcaceae UCG-002 and Prevotellaceae YAB2003 group bur decreased abundance of Ruminococcaceae NK4A214 and Syntrophococcus in SHS bulls compared to SH. | [43] |
| Saponins | Holstein cows (658 kg) | CTR: no supplementation TEA: 0.77% tea saponin for 5 weeks | No effects on alpha diversity. Beta diversity analysis showed no differences in microbial composition between CTR and TEA cows. Decreased abundance of UC Deltaproteobacteria in TEA cows. Slight reduction in microbial network complexity (fewest edges) in TEA cows. | [44] |
| Disturbance | Animals | Treatments | Effects on Ruminal Microbiota 1,2 | Reference |
|---|---|---|---|---|
| Probiotic administration | Crossbred steers (434 kg) | CTR: no probiotic administration. P169: administration of Propionibacterium acidipropionici strain P169 (1011 cfu/d) during the finishing period. | No effects on alpha diversity. Beta diversity analysis showed no differences in microbial community composition between CTR and P169 steers. Increased gene copy numbers of Propionibacterium acidipropionici strain P169 in P169 steers by qPCR. At phylum level, no differences in taxon abundance between CTR and P169 steers. At genus level, increased abundance of Phascolarctobacterium, UC Clostridiaceae and Lachnospiraceae, whereas decreased abundance of Prevotella, Succinivibrio, YRC22 and UC Veillonellaceae in P169 steers. | [45] |
| Probiotic administration | Romane lambs (fattening) | CTR: no probiotic administration. SUP: administration of a combination of live yeast Saccharomyces cerevisiae CNCM I-1077 and selected yeast metabolites in milk replacer (3 × 109 cfu/d plus 0.45 g yeast metabolites/d) and in feed (6 × 106 cfu/g plus 1.5 kg yeast metabolites/ton) during the whole fattening period. | No effects on alpha diversity. Beta diversity analysis showed no differences in microbial community composition between CTR and SUP lambs. At OTU level, increased abundance of Snodrgrassella, Megasphaera, Bifidobacterium, Butyricimonas, Succinivibrio and Fibrobacter, whereas decreased abundance of Desulfovibrio and Bacteroides. | [46] |
| Probiotic administration | Holstein steers (504 kg) | CON: no probiotic administration. YEA: administration of a feed additive containing Saccharomyces cerevisiae and other active ingredients from yeast cell wall (15 g commercial product/d for 25 d). | No effects on alpha diversity. Beta diversity analysis showed significant differences in microbial community composition between CON and YEA steers. At phylum level, increased abundance of Saccharibacteria in YEA steers. At genus level, increased abundance of Ruminococcaceae NK4A214, Christensenellaceae R-7, Ruminococcaceae UCG-010, Candidatus Saccharimonas, Bacteroidales BS11 gut group, Ruminococcus 2, Anaerovorax, Lachnospiraceae UCG-008 and Ruminococcaceae UCG-005, whereas decreased abundance of Lachnoclostridium, Lachnoclostridium 5 and Bacillus in YEA steers. | [47] |
| Probiotic administration | Jintang black male goats (80 d) | CTR: no probiotic administration BA: administration of Bacillus amyloliquefaciens fszne-06 (109 cfu every 2 d for 30 d) BP: administration of Bacillus pumilus fszne-09 (109 cfu every 2 d for 30 d) | Increased alpha diversity (increased values of Shannon and Simpson indices and richness) in BA and BP goats. Beta diversity analysis showed significant differences in microbial community composition between treatments. At the phylum level, increased abundance of Firmicutes in BA and BP goats and decreased abundance of Bacteroidetes in BA goats. At the genus level, increased abundance of Succiniclasticum in BA and of UC Ruminococcaceae in BP goats. Decreased abundance of Klebsiella in BA and BP goats. | [48] |
| Inoculation with rumen content from donor animals | Donors: Hu sheep (36 kg) Recipients: Hu lambs (1–28 d) | C: non-transfaunated lambs. IBW: lambs transfaunated before weaning (20 mL of sheep ruminal fluid mixture via stomach tube, four inoculations) IDW: lambs transfaunated during weaning (20 mL of sheep ruminal fluid mixture, two inoculations) | Increased alpha diversity in donor ruminal fluid mixture did not translate to increased alpha diversity in transfaunated lambs. Beta diversity analysis showed no differences in microbial community composition between treatments. At the genus level, increased abundance of Prevotellaceae UCG-001, Moryella, Succiniclasticum and Tyzzerella 4 in IBW lambs compared to C and increased abundance of Erysipelatoclostridium, Eubacterium coprostanoligenes and Sharpea in IDW lambs compared to C. | [49] |
| Inoculation with rumen content from donor animals | Donors: crossbred cows (adults) Recipients: sheep (1–4 years, 35 kg) | CON: non-transfaunated sheep TRANS: transfaunated sheep (administration of 1.5 L of cow ruminal fluid mixture via stomach tube once) | Increased alpha diversity in donor ruminal fluid mixture did not translate to increased alpha diversity in TRANS sheep. Beta diversity analysis showed changes in community structure but not membership in TRANS sheep, suggesting that the new species introduced by transfaunation were not able to colonize recipient rumen but favored or inhibited growth of some established species. At the phylum level, no differences in taxon abundance between CON and TRANS sheep. At the genus level, decreased abundance of Selenomonas in TRANS sheep. | [50] |
| Disturbance | Animals | Treatments 1 | Effects on Ruminal Microbiota 1,2 | Reference |
|---|---|---|---|---|
| Complete exchange of rumen content from donor animals | Holstein cows (multiparous) | HE: high-efficiency cows exchanged rumen content with low-efficiency cows LE: low-efficiency cows exchanged rumen content with high-efficiency cows Sampling period included: Pre: from 8 to 0 d before exchange. Post 1: from 0 to 10 d after exchange. Post 2: from 10 to 56 d after exchange. | In Pre, LE cows exhibited higher alpha diversity than HE cows. After exchange, HE cows showed increased Shannon index and richness values in Post 1 and returned to pre-exchange levels in Post 2. LE cows had decreased Shannon index values in Post 1 and returned to pre-exchange levels in Post 2, whereas no change in richness values was observed over time. Beta diversity analysis showed significant differences in microbial community composition between treatments. The general trend was that of a donor-like community in Post 1 induced by the exchange and, in contrast to Pre, returned to a community similar to Pre in the host in Post 2. | [51] |
| Complete exchange of rumen content from donor animals | Holstein cows (multiparous, 582 kg) | SARA induction period: CON: 40% concentrate diet HG: 60% concentrate diet After the SARA induction period, the rumen content transplant period began, and cows in the HG group were categorized as: DR: cows receiving 70% rumen content from CON cows SR: cows receiving 70% self-derived rumen content Sampling period lasted until 20 d after exchange. | Increased alpha diversity (increased values of Shannon index) in DR cows. Beta diversity analysis showed significant differences in microbial community composition between DR and SR, despite samples starting to cluster together 4 d following exchange. At the phylum level, increased abundance of Firmicutes and decreased abundance of Bacteroidetes and Spirochaetes in DR cows. At the genus level, increased abundance of UC Ruminococcaceae and Saccharofermentans but decreased abundance of UC Prevotellaceae and Treponema in DR cows. Microbial network analysis showed that rumen content exchange only affects non-keystone OTUs (i.e., taxa that display weak interactions with other taxa). | [52] |
| Changes Immediately Following a Disturbance 1 | Changes at a Certain Time Point After a Disturbance 1 | Reference |
|---|---|---|
| [72] | ||
| [73] | ||
| [74] | ||
| [75] | ||
| [76] | ||
| [77] | ||
| [78] | ||
| [79] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Costa-Roura, S.; Villalba, D.; Balcells, J.; De la Fuente, G. First Steps into Ruminal Microbiota Robustness. Animals 2022, 12, 2366. https://doi.org/10.3390/ani12182366
Costa-Roura S, Villalba D, Balcells J, De la Fuente G. First Steps into Ruminal Microbiota Robustness. Animals. 2022; 12(18):2366. https://doi.org/10.3390/ani12182366
Chicago/Turabian StyleCosta-Roura, Sandra, Daniel Villalba, Joaquim Balcells, and Gabriel De la Fuente. 2022. "First Steps into Ruminal Microbiota Robustness" Animals 12, no. 18: 2366. https://doi.org/10.3390/ani12182366
APA StyleCosta-Roura, S., Villalba, D., Balcells, J., & De la Fuente, G. (2022). First Steps into Ruminal Microbiota Robustness. Animals, 12(18), 2366. https://doi.org/10.3390/ani12182366