
Effects of Compound Feed Attractants on Growth Performance, Feed Utilization, Intestinal Histology, Protein Synthesis, and Immune Response of White Shrimp (Litopenaeus Vannamei)

 ,
,  ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Experimental Shrimp and Feeding
2.3. Sampling and Chemical Analysis
2.4. Calculation Formula and Statistical Analysis
3. Results
3.1. Growth Performance, Feed Utilization, and Morphological Parameters
3.2. Whole Body and Muscle Composition of the Shrimp L. vannamei
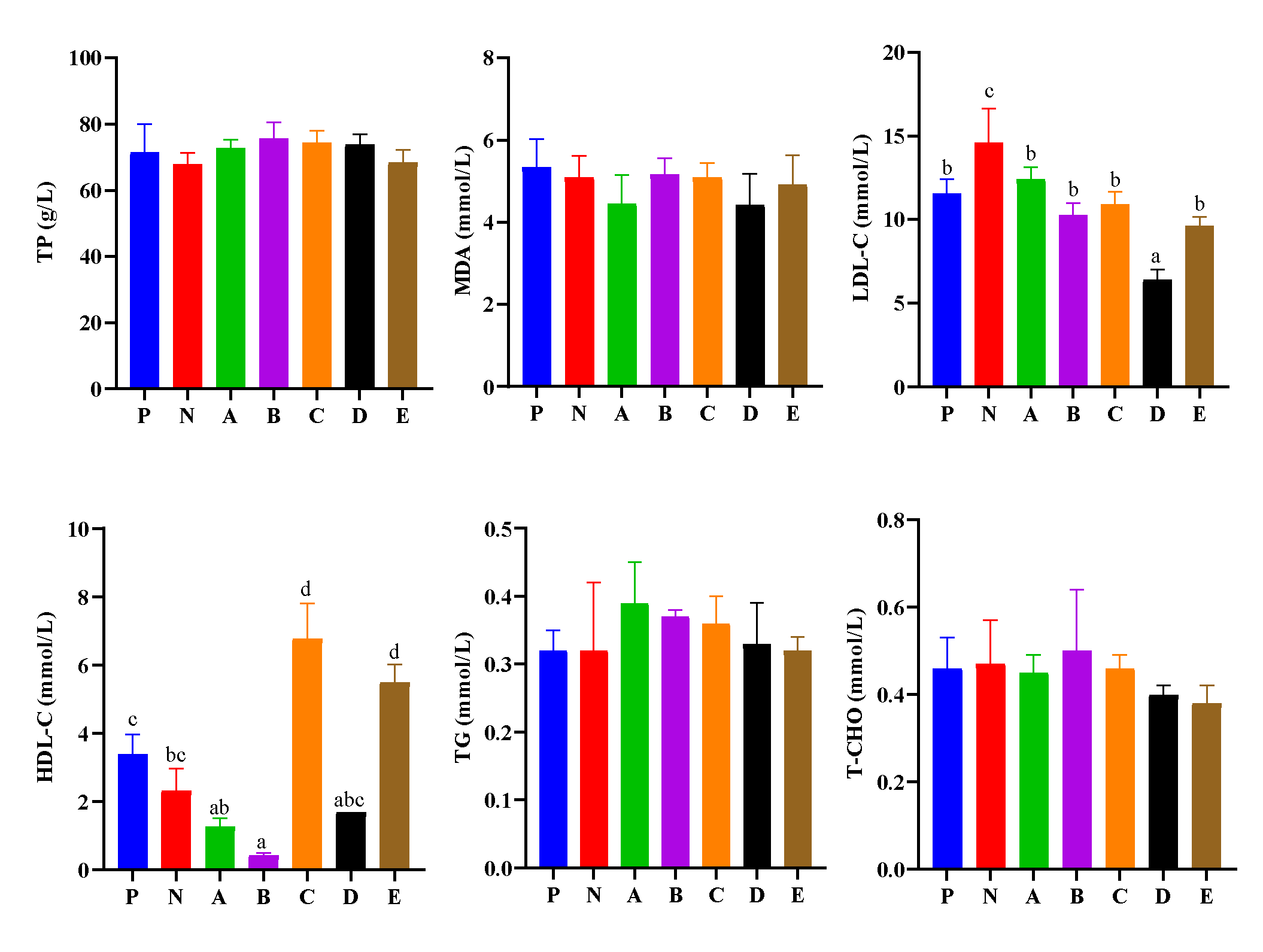
3.3. Hemolymph Biochemical Parameters
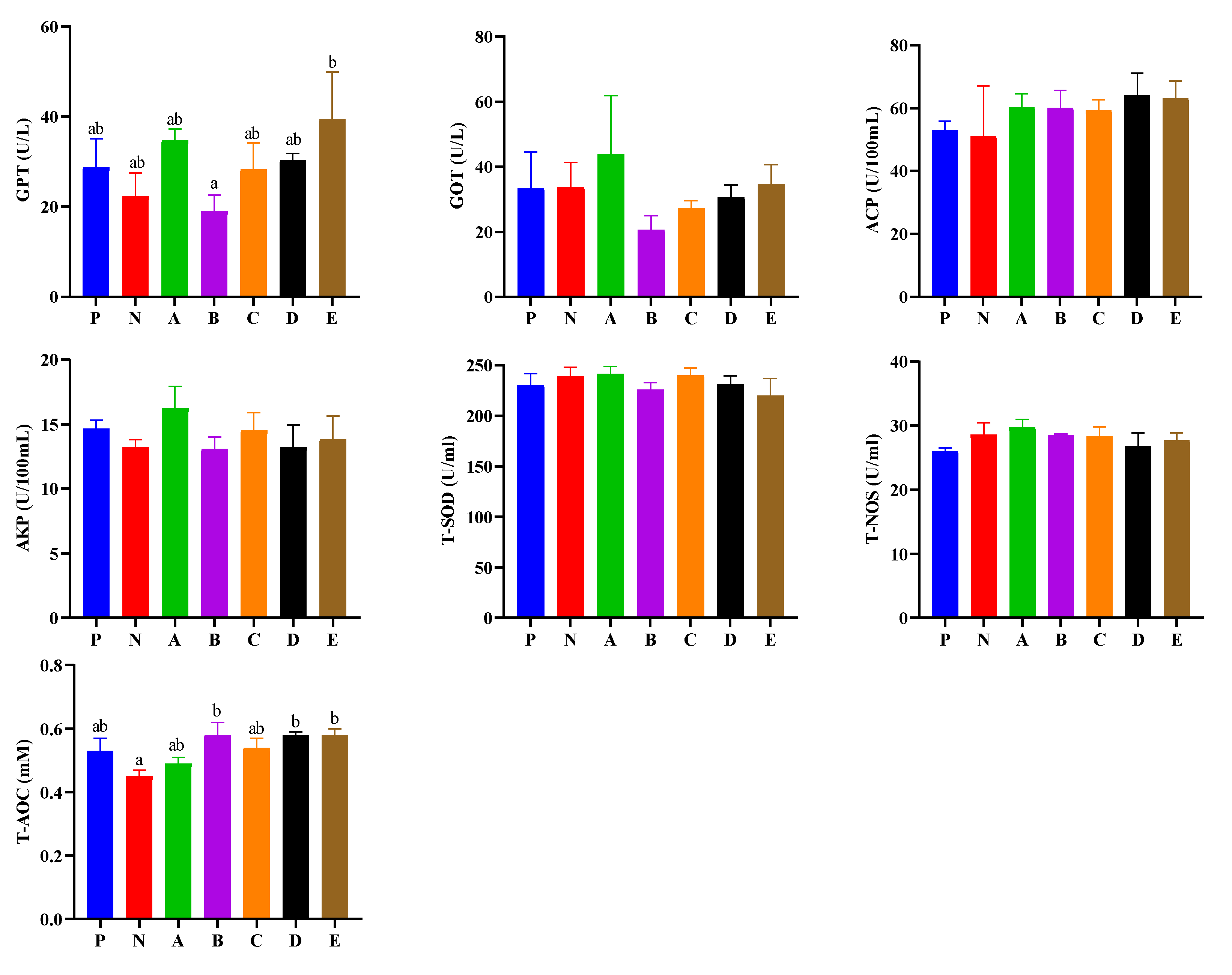
3.4. Evaluation of Hemolymph Enzyme Activities
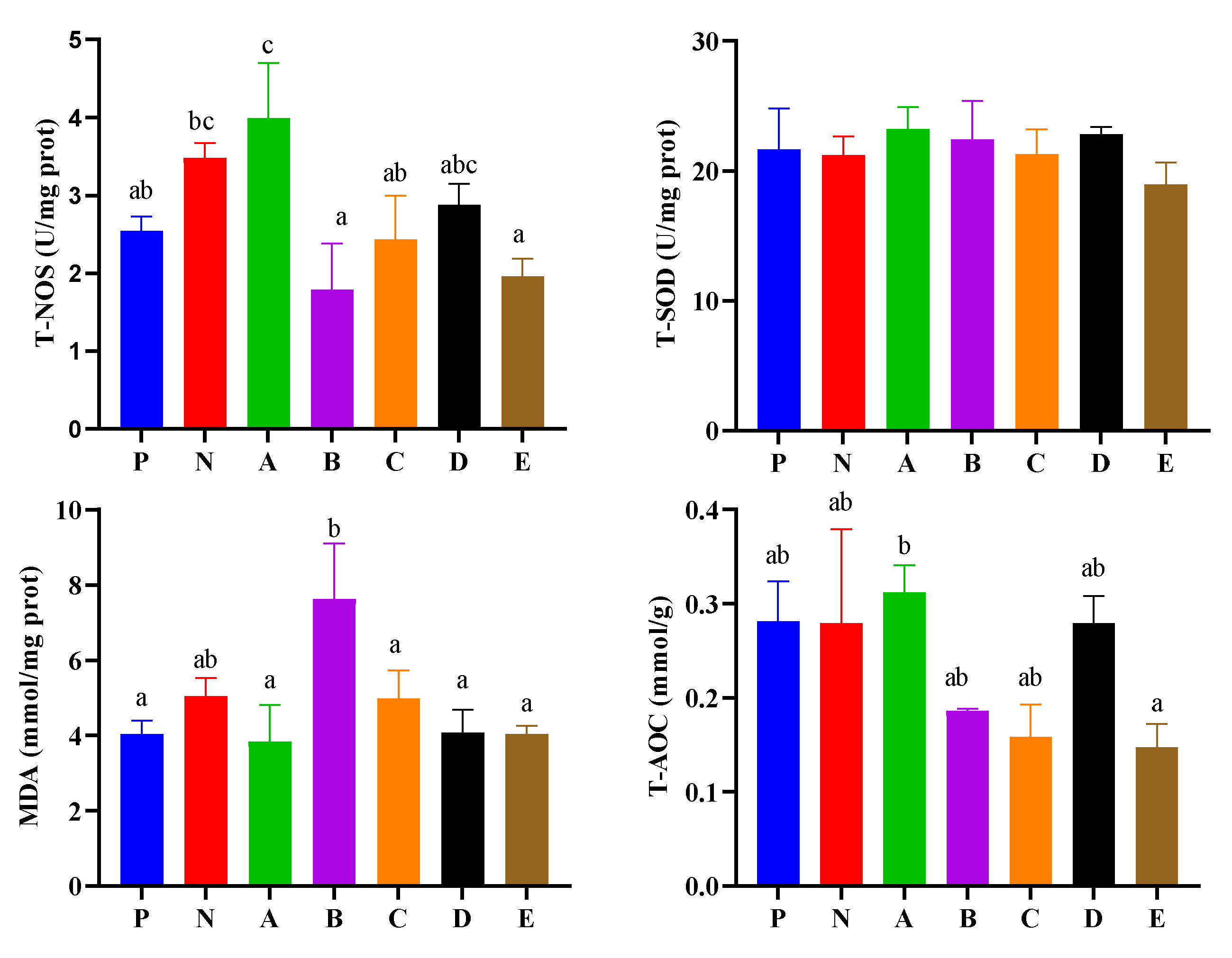
3.5. Enzyme Activities in the Hepatopancreas
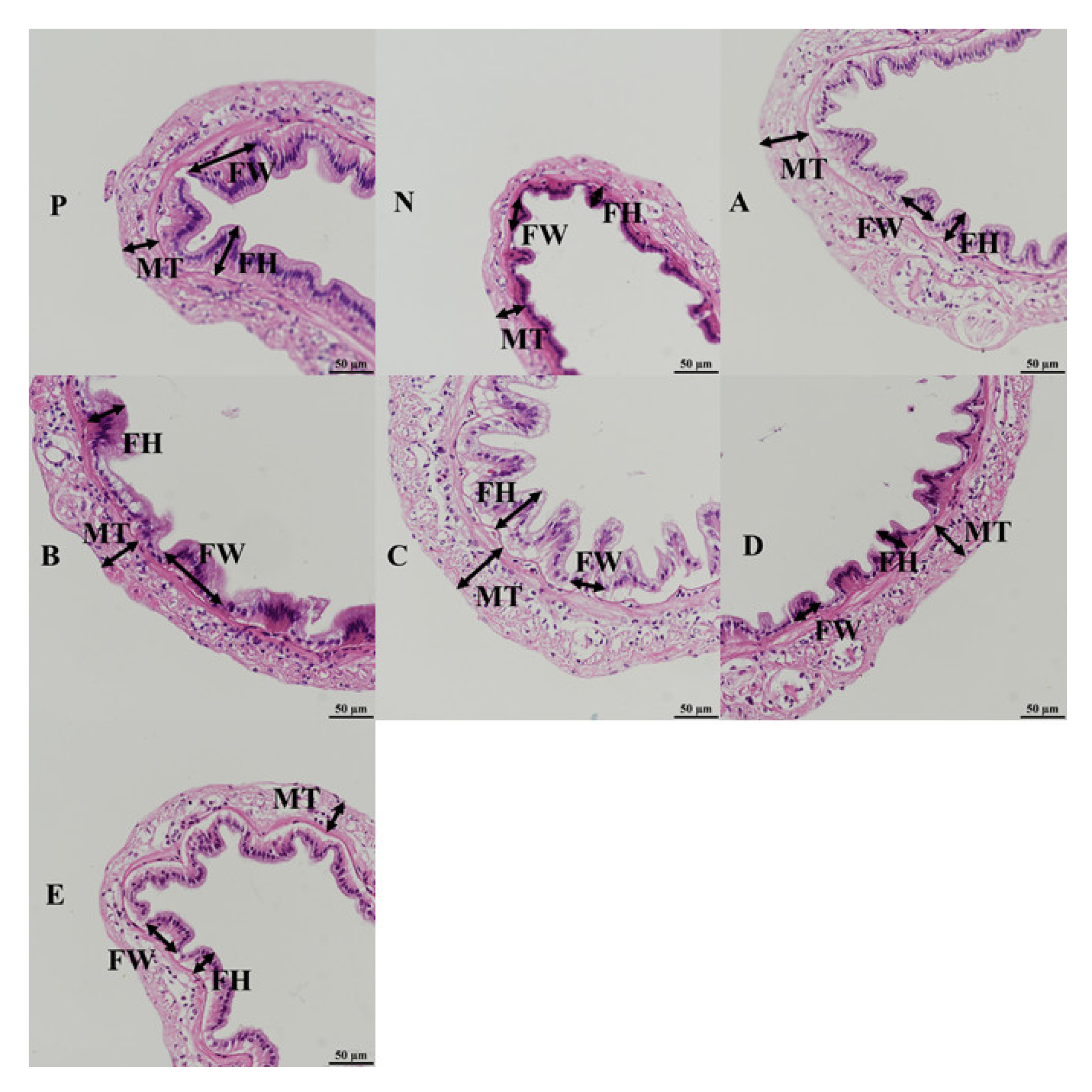
3.6. Intestinal Histology
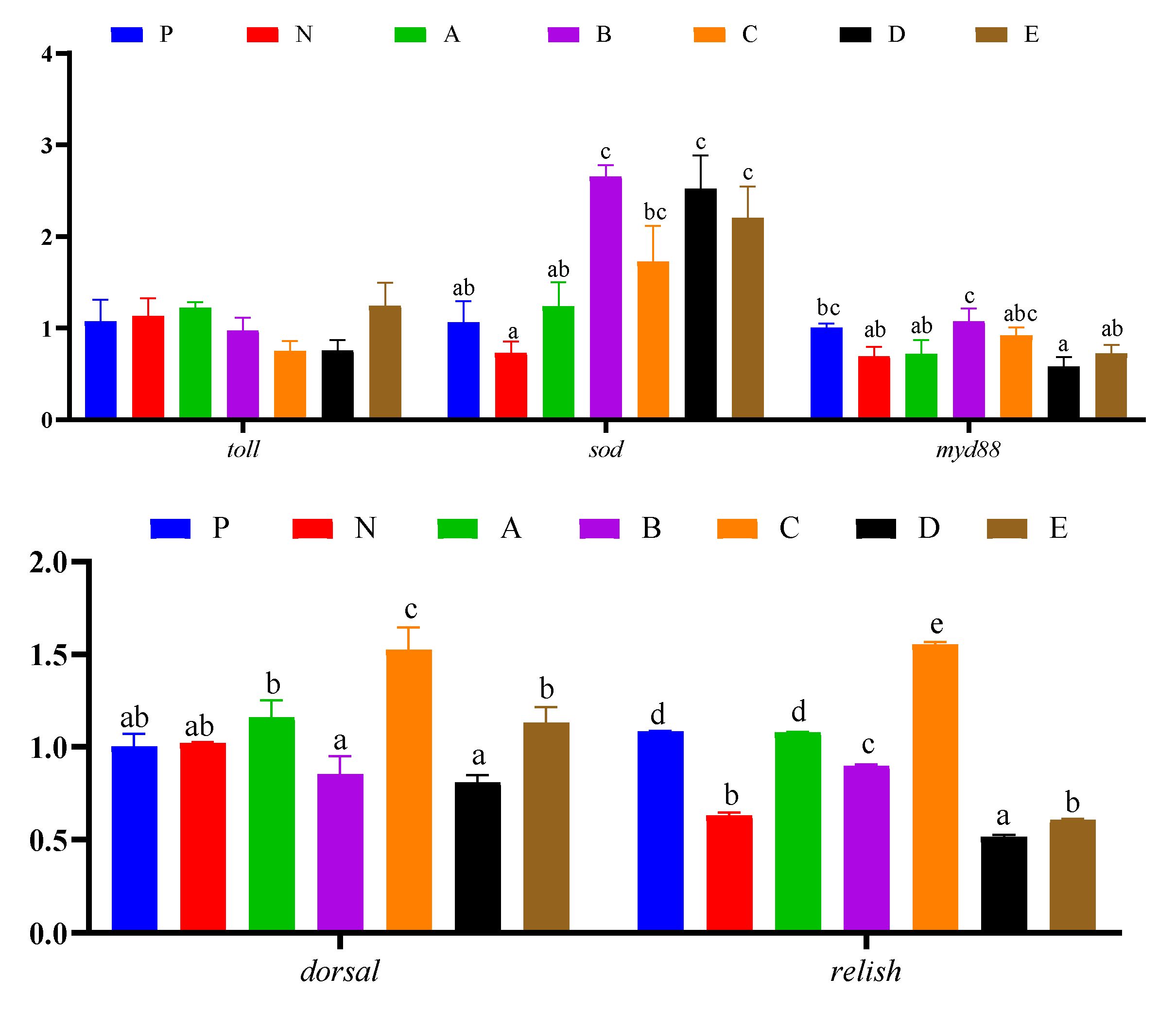
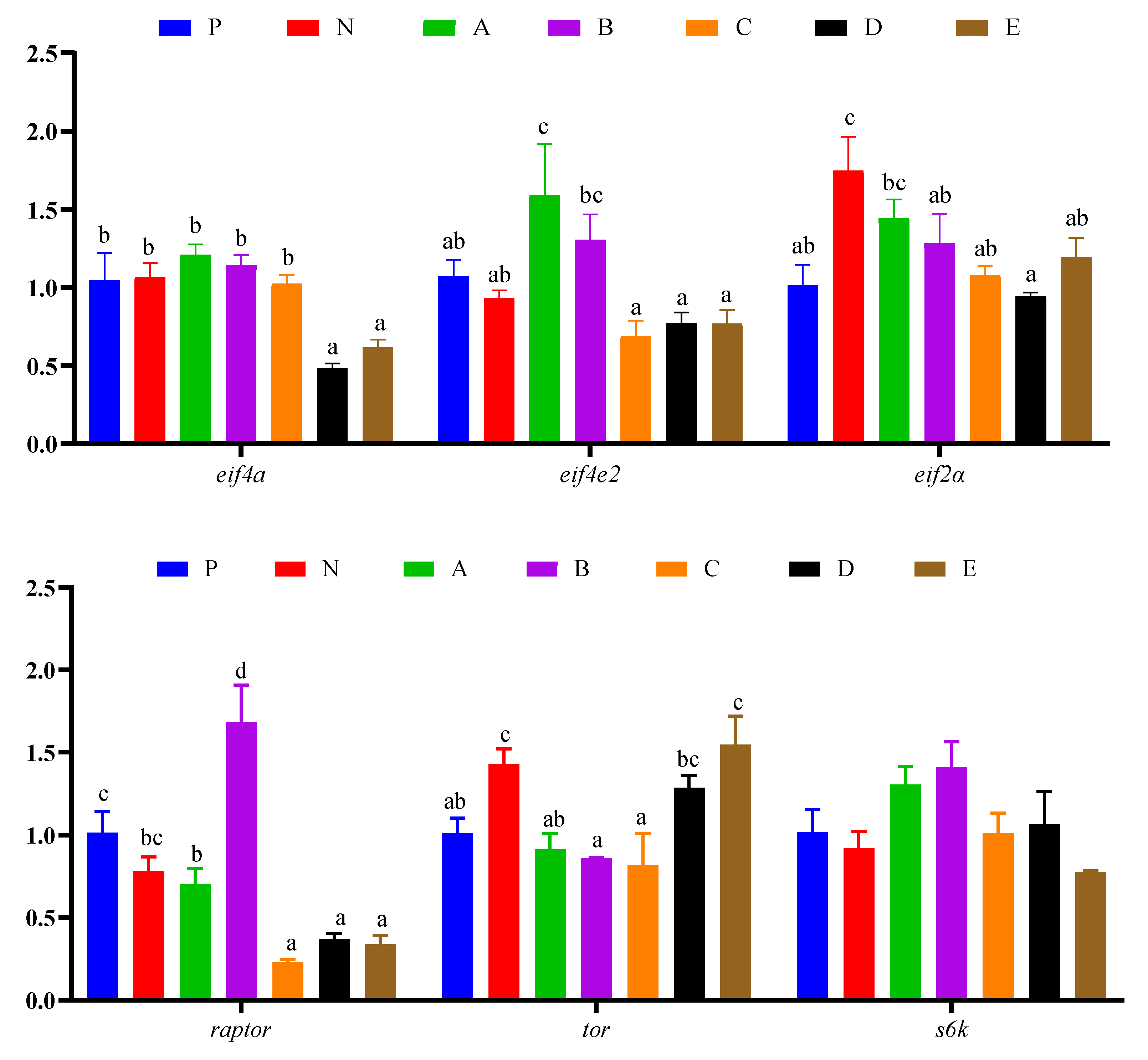
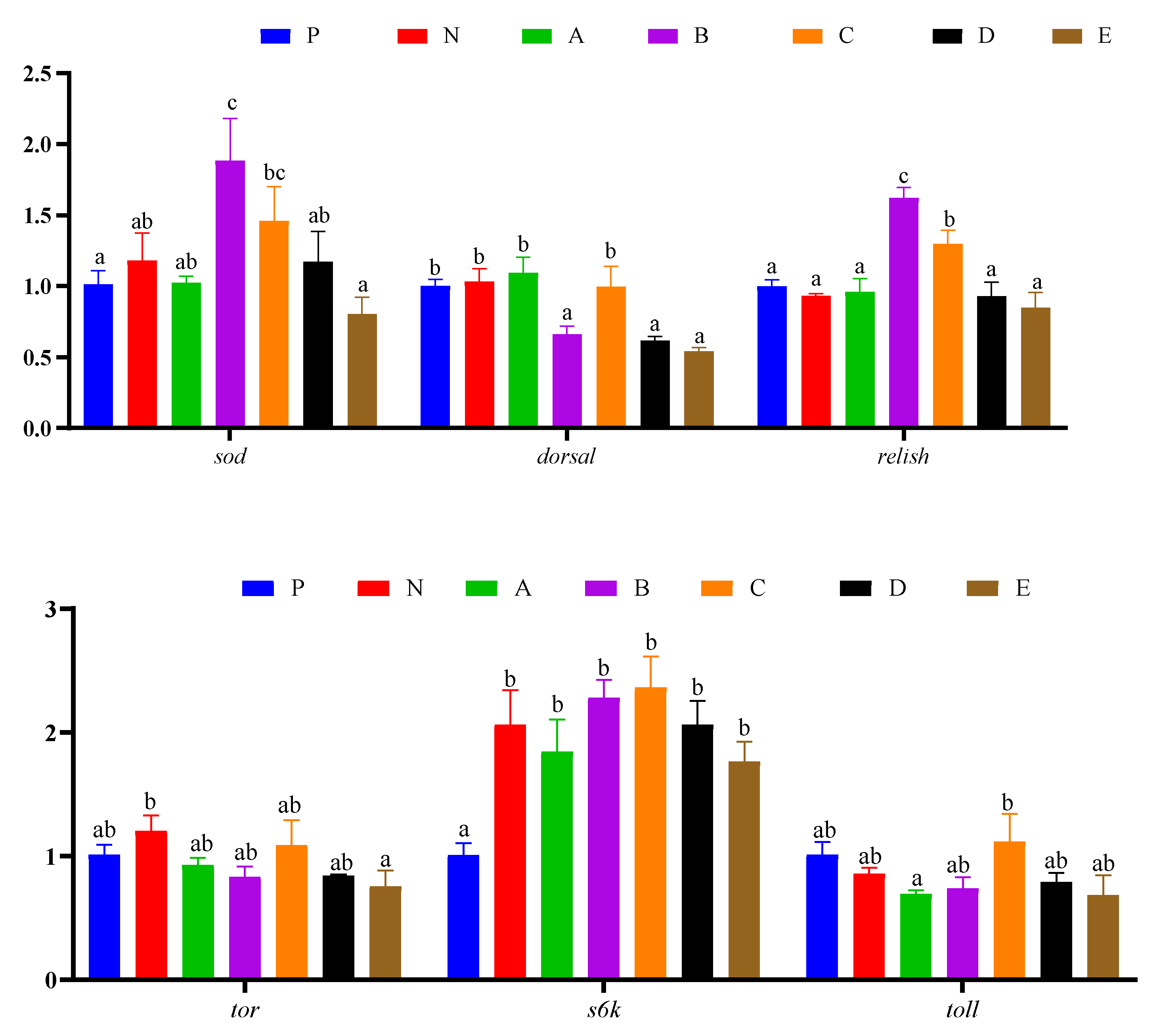
3.7. Immune Response and Protein Synthesis Related Genes Expressions in Hepatopancreas
3.8. Intestinal Genes Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, S.; Guo, Y.; Espe, M.; Yang, F.; Fang, W.; Wan, M.; Niu, J.; Liu, Y.; Tian, L. Growth performance, haematological parameters, antioxidant status and salinity stress tolerance of juvenile Pacific white shrimp (Litopenaeus vannamei) fed different levels of dietary myo-inositol. Aquac. Nutr. 2018, 24, 1527–1539. [Google Scholar] [CrossRef]
- Barang, M. Fishery and Aquaculture Statistics. In FAO Yearbook. Fishery and Aquaculture Statistics = FAO Annuaire. Statistiques des Peches et de l’Aquaculture = FAO Anuario. Estadisticas de Pesca y Acuicultura; FAO: Rome, Italy, 2018; p. I-82. [Google Scholar]
- Chen, Y.; Chi, S.; Zhang, S.; Dong, X.; Yang, Q.; Liu, H.; Tan, B.; Xie, S. Evaluation of Methanotroph (Methylococcus capsulatus, Bath) bacteria meal on body composition, lipid metabolism, protein synthesis and muscle metabolites of Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2022, 547, 737517. [Google Scholar] [CrossRef]
- Yeşilayer, N.; Kaymak, I.E. Effect of partial replacement of dietary fish meal by soybean meal with betaine attractant supplementation on growth performance and fatty acid profiles of juvenile rainbow trout (Oncorhynchus mykiss). Aquac. Res. 2020, 51, 1533–1541. [Google Scholar] [CrossRef]
- Liu, Y.; Meng, L.; Zhang, J. Effect of partial fish meal replacement by soybean meal on the growth performance, Fuscoguttatus. China Feed 2018, 4, 60–64. [Google Scholar]
- Jiang, D.; Zheng, J.; Dan, Z.; Tang, Z.; Ai, Q.; Mai, K. Effects of five compound attractants in high plant-based diets on feed intake and growth performance of juvenile turbot (Scophthalmus maximus L.). Aquac. Res. 2019, 50, 2350–2358. [Google Scholar] [CrossRef]
- Tusche, K.; Berends, K.; Wuertz, S.; Susenbeth, A.; Schulz, C. Evaluation of feed attractants in potato protein concentrate based diets for rainbow trout (Oncorhynchus mykiss). Aquaculture 2011, 321, 54–60. [Google Scholar] [CrossRef]
- Huang, F.; Wang, L.; Zhang, C.; Song, K. Replacement of fishmeal with soybean meal and mineral supplements in diets of Litopenaeus vannamei reared in low-salinity water. Aquaculture 2017, 473, 172–180. [Google Scholar] [CrossRef]
- Shan, H.; Zhang, L.; Gao, L.; Su, Y.; Bao, W.; Ma, S. A comparative study of intensive Litopenaeus vannamei culture on four bottom substrates without water change. J. Ocean Univ. China 2014, 13, 705–711. [Google Scholar] [CrossRef]
- Pittet, A.; Ellis, J.; Lee, P. Methodology for the identification and quantitative measurement of chemical stimulants for penaeid shrimp. Aquac. Nutr. 1996, 2, 175–182. [Google Scholar] [CrossRef]
- Sanchez, D.R.; Fox, J.M.; Lawrence, A.L.; Castille, F.L.; Dunsford, B. A methodology for evaluation of dietary feeding stimulants for the Pacific white shrimp, Litopenaeus vannamei. J. World Aquac. Soc. 2005, 36, 14–23. [Google Scholar] [CrossRef]
- Hancz, C. Feed efficiency, nutrient sensing and feeding stimulation in aquaculture: A review. Acta Agrar. Kaposváriensis 2020, 24, 35–54. [Google Scholar] [CrossRef]
- Zheng, L.; Xie, S.; Zhuang, Z.; Liu, Y.; Tian, L.; Niu, J. Effects of yeast and yeast extract on growth performance, antioxidant ability and intestinal microbiota of juvenile Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2021, 530, 735941. [Google Scholar] [CrossRef]
- Jin, M.; Xiong, J.; Zhou, Q.-C.; Yuan, Y.; Wang, X.-X.; Sun, P. Dietary yeast hydrolysate and brewer’s yeast supplementation could enhance growth performance, innate immunity capacity and ammonia nitrogen stress resistance ability of Pacific white shrimp (Litopenaeus vannamei). Fish Shellfish Immunol. 2018, 82, 121–129. [Google Scholar] [CrossRef]
- Zhang, R.; Jiang, Y.; Zhou, L.; Chen, Y.; Wen, C.; Liu, W.; Zhou, Y. Effects of dietary yeast extract supplementation on growth, body composition, non-specific immunity, and antioxidant status of Chinese mitten crab (Eriocheir sinensis). Fish Shellfish Immunol. 2019, 86, 1019–1025. [Google Scholar] [CrossRef]
- He, Y.; Chi, S.; Tan, B.; Zhang, H.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S. Effect of yeast culture on intestinal microbiota of Litopenaeus vannamei. J. Guangdong Ocean Univ. 2017, 37, 21–27. [Google Scholar]
- Toften, H.; Arnesen, A.M.; Jobling, M. Feed intake, growth and ionoregulation in Atlantic salmon (Salmo salar L.) smolts in relation to dietary addition of a feeding stimulant and time of seawater transfer. Aquaculture 2003, 217, 647–662. [Google Scholar] [CrossRef]
- Zhu, T.; Morais, S.; Luo, J.; Jin, M.; Lu, Y.; Le, Y.; Zhou, Q. Functional palatability enhancer improved growth, intestinal morphology, and hepatopancreas protease activity, replacing squid paste in white shrimp, Litopenaeus vannamei, diets. J. World Aquac. Soc. 2019, 50, 1064–1077. [Google Scholar] [CrossRef]
- Harrison, G.; Coates, M.E. Interrelationship between the growth-promoting effect of fish solubles and the gut flora of the chick. Br. J. Nutr. 1972, 28, 213–221. [Google Scholar] [CrossRef]
- Cai, L.; Liu, S.; Sun, L.; Wang, Y.; Ji, H.; Li, J. Application of tea polyphenols in combination with 6-gingerol on shrimp paste of during storage: Biogenic amines formation and quality determination. Front. Microbiol. 2015, 6, 981. [Google Scholar] [CrossRef]
- Lim, L.S.; Lai, S.J.; Yong, S.K.; Shapawi, R.; Kawamura, G. Feeding response of marble goby (Oxyeleotris marmorata) to organic acids, amino acids, sugars and some classical taste substances. Appl. Anim. Behav. Sci. 2017, 190, 113–118. [Google Scholar] [CrossRef]
- Xue, M.; Xie, S.; Cui, Y. Effect of a feeding stimulant on feeding adaptation of gibel carp Carassius auratus gibelio (Bloch), fed diets with replacement of fish meal by meat and bone meal. Aquac. Res. 2004, 35, 473–482. [Google Scholar] [CrossRef]
- Qiu, X.; Davis, D. Evaluation of flash dried yeast as a nutritional supplement in plant-based practical diets for Pacific white shrimp Litopenaeus vannamei. Aquac. Nutr. 2017, 23, 1244–1253. [Google Scholar] [CrossRef]
- Gui, L.; Mai, H.; Chi, S.; Zhou, W.; Li, Y.; Tan, B.; Dong, X.; Yang, Q.; LIU, h.; Zhang, S. Effects of Yeast Culture on Growth Performance, Hematological Parameters, Immunity and Disease Resistance of Litopenaeus vannamei. J. Guangdong Ocean Univ. 2019, 39, 30–37. [Google Scholar]
- AOAC. Official Method of Analysis of The Association of Official Analytical Chemistry, 16th ed.; Association of Official Analytical Chemists: Arlingron, VA, USA, 1995. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Kader, M.A.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Bulbul, M. Supplemental effects of some crude ingredients in improving nutritive values of low fishmeal diets for red sea bream, Pagrus major. Aquaculture 2010, 308, 136–144. [Google Scholar] [CrossRef]
- Fang, H.; Xie, J.; Liao, S.; Guo, T.; Xie, S.; Liu, Y.; Tian, L.; Niu, J. Effects of Dietary Inclusion of Shrimp Paste on Growth Performance, Digestive Enzymes Activities, Antioxidant and Immunological Status and Intestinal Morphology of Hybrid Snakehead (Channa maculata ♀ × Channa argus ♂). Front. Physiol. 2019, 10, 1–8. [Google Scholar] [CrossRef]
- Peng, D.; Peng, B.; Li, J.; Zhang, Y.; Luo, H.; Xiao, Q.; Tang, S.; Liang, X.-F. Effects of three feed attractants on the growth, biochemical indicators, lipid metabolism and appetite of Chinese perch (Siniperca chuatsi). Aquac. Rep. 2022, 23, 101075. [Google Scholar] [CrossRef]
- Deng, D.; Mei, C.; Mai, K.; Tan, B.P.; Ai, Q.; Ma, H. Effects of a yeast-based additive on growth and immune responses of white shrimp, Litopenaeus vannamei (B oone, 1931), and aquaculture environment. Aquac. Res. 2013, 44, 1348–1357. [Google Scholar] [CrossRef]
- Utne-Palm, A.C.; Bogevik, A.S.; Humborstad, O.-B.; Aspevik, T.; Pennington, M.; Løkkeborg, S. Feeding response of Atlantic cod (Gadus morhua) to attractants made from by-products from the fishing industry. Fish. Res. 2020, 227, 105535. [Google Scholar] [CrossRef]
- Hoseinifar, S.H.; Mirvaghefi, A.; Merrifield, D.L.; Amiri, B.M.; Yelghi, S.; Bastami, K.D. The study of some haematological and serum biochemical parameters of juvenile beluga (Huso huso) fed oligofructose. Fish Physiol. Biochem. 2011, 37, 91–96. [Google Scholar] [CrossRef] [PubMed]
- Brum, A.; Pereira, S.A.; Cardoso, L.; Chagas, E.C.; Chaves, F.C.M.; Mouriño, J.L.P.; Martins, M.L. Blood biochemical parameters and melanomacrophage centers in Nile tilapia fed essential oils of clove basil and ginger. Fish Shellfish Immunol. 2018, 74, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Ollila, O.S.; Lamberg, A.; Lehtivaara, M.; Koivuniemi, A.; Vattulainen, I. Interfacial tension and surface pressure of high density lipoprotein, low density lipoprotein, and related lipid droplets. Biophys. J. 2012, 103, 1236–1244. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Yang, Q.; Tan, B.; Wang, F.; Dong, X.; Chi, S.; Liu, H.; Zhang, S.; Wang, H. Study of the requirements of dietary cholesterol at two different growth stages of Pacific white shrimps, Litopenaeus vannamei. Aquac. Int. 2019, 27, 1583–1597. [Google Scholar] [CrossRef]
- Cai, R.; Zhang, J.; Huang, J.; Chen, G.; Zhang, J.; Pan, C.; Wang, Z.; Xie, R.; Tang, B. Effects of Low Temperature Stress on Physiology and Biochemistry of Lipid Metabolism of Juvenile Cobia, Rachycentron canadum. J. Guangdong Ocean Univ. 2021, 41, 123–130. [Google Scholar]
- Grao-Cruces, E.; Varela, L.M.; Martin, M.E.; Bermudez, B.; Montserrat-de la Paz, S. High-Density Lipoproteins and Mediterranean Diet: A Systematic Review. Nutrients 2021, 13, 955. [Google Scholar] [CrossRef]
- Kalhoro, H.; Zhou, J.; Hua, Y.; Ng, W.K.; Ye, L.; Zhang, J.; Shao, Q. Soy protein concentrate as a substitute for fish meal in diets for juvenile Acanthopagrus schlegelii: Effects on growth, phosphorus discharge and digestive enzyme activity. Aquac. Res. 2018, 49, 1896–1906. [Google Scholar] [CrossRef]
- Nazeri, S.; Farhangi, M.; Modarres, S. The effect of different dietary inclusion levels of rutin (a flavonoid) on some liver enzyme activities and oxidative stress indices in rainbow trout, Oncorhynchus mykiss (Walbaum) exposed to Oxytetracycline. Aquac. Res. 2017, 48, 4356–4362. [Google Scholar] [CrossRef]
- Xie, S.; Wei, D.; Fang, W.; Yin, P.; Liu, Y.; Niu, J.; Tian, L. Survival and protein synthesis of post-larval White Shrimp, Litopenaeus vannamei were affected by dietary protein level. Anim. Feed Sci. Technol. 2020, 263, 114462. [Google Scholar] [CrossRef]
- Xie, S.; Zhou, W.; Tian, L.; Niu, J.; Liu, Y. Effect of N-acetyl cysteine and glycine supplementation on growth performance, glutathione synthesis, anti-oxidative and immune ability of Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 2016, 55, 233–241. [Google Scholar] [CrossRef]
- Rašić, I.; Rašić, A.; Akšamija, G.; Radović, S. The relationship between serum level of malondialdehyde and progression of colorectal cancer. Acta Clin. Croat. 2018, 57, 411–416. [Google Scholar] [CrossRef]
- Moghadam, H.; Sourinejad, I.; Johari, S.A. Growth performance, haemato-immunological responses and antioxidant status of Pacific white shrimp Penaeus vannamei fed with turmeric powder, curcumin and curcumin nanomicelles. Aquac. Nutr. 2021, 27, 2294–2306. [Google Scholar] [CrossRef]
- Garcia, V.; Sessa, W.C. Endothelial NOS: Perspective and recent developments. Br. J. Pharmacol. 2019, 176, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Chen, T.; Wong, N.-K.; Jiang, X.; Luo, X.; Zhang, L.; Yang, D.; Ren, C.; Hu, C. Nitric oxide as an antimicrobial molecule against Vibrio harveyi infection in the hepatopancreas of Pacific white shrimp, Litopenaeus vannamei. Fish Shellfish Immunol. 2015, 42, 114–120. [Google Scholar] [CrossRef] [PubMed]
- Zhu, W.; Qiu, D.; Gan, Z.; Lu, Y.; Jian, J. Antivirus Effects of Vibrio alginolyticus Peptidoglycan on Litopenaeus vannamei against White Spot Syndrome Virus. J. Guangdong Ocean Univ. 2015, 35, 40–46. [Google Scholar]
- Ostaszewska, T.; Dabrowski, K.; Kamaszewski, M.; Grochowski, P.; Verri, T.; Rzepkowska, M.; Wolnicki, J. The effect of plant protein-based diet supplemented with dipeptide or free amino acids on digestive tract morphology and PepT1 and PepT2 expressions in common carp (Cyprinus carpio L.). Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2010, 157, 158–169. [Google Scholar] [CrossRef]
- Xie, S.; Wei, D.; Tian, L.; Liu, Y. Dietary supplementation of chenodeoxycholic acid improved the growth performance, immune response and intestinal health of juvenile Penaeus monodon fed a low fish-meal diet. Aquac. Rep. 2021, 20, 100773. [Google Scholar] [CrossRef]
- Li, J.; Xu, Q.; Wang, C.; Luo, L.; Zhao, Z. Effects of glutamine and its precursors on intestinal digestive enzyme activity and intestinal morphology of Songpu mirror carp (Cyprinus carpio specularis). Chin. J. Anim. Nutr. 2014, 26, 1347–1352. [Google Scholar]
- Chen, F.; Wang, H. Study on histological structure of intestine in Gymnocypris przewalskii with different age. Prog. Vet. Med. 2013, 34, 34–37. [Google Scholar]
- Xiong, J.; Jin, M.; Yuan, Y.; Luo, J.X.; Lu, Y.; Zhou, Q.C.; Liang, C.; Tan, Z.L. Dietary nucleotide-rich yeast supplementation improves growth, innate immunity and intestinal morphology of Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1425–1435. [Google Scholar] [CrossRef]
- Anthoney, N.; Foldi, I.; Hidalgo, A. Toll and Toll-like receptor signalling in development. Development 2018, 145, dev156018. [Google Scholar] [CrossRef]
- Hayden, M.S.; Ghosh, S. NF-κB in immunobiology. Cell Res. 2011, 21, 223–244. [Google Scholar] [CrossRef]
- Huang, X.-D.; Yin, Z.-X.; Liao, J.-X.; Wang, P.-H.; Yang, L.-S.; Ai, H.-S.; Gu, Z.-H.; Jia, X.-T.; Weng, S.-P.; Yu, X.-Q. Identification and functional study of a shrimp Relish homologue. Fish Shellfish Immunol. 2009, 27, 230–238. [Google Scholar] [CrossRef]
- Hetru, C.; Hoffmann, J.A. NF-κB in the immune response of Drosophila. Cold Spring Harb. Perspect. Biol. 2009, 1, a000232. [Google Scholar] [CrossRef]
- Li, F.; Yan, H.; Wang, D.; Priya, T.J.; Li, S.; Wang, B.; Zhang, J.; Xiang, J. Identification of a novel relish homolog in Chinese shrimp Fenneropenaeus chinensis and its function in regulating the transcription of antimicrobial peptides. Dev. Comp. Immunol. 2009, 33, 1093–1101. [Google Scholar] [CrossRef]
- Li, F.; Wang, D.; Li, S.; Hui, Y.; Zhang, J.; Wang, B.; Zhang, J.; Xiang, J. A Dorsal homolog (FcDorsal) in the Chinese shrimp Fenneropenaeus chinensis is responsive to both bacteria and WSSV challenge. Dev. Comp. Immunol. 2010, 34, 874–883. [Google Scholar] [CrossRef]
- Guo, J.; Guo, B.; Zhang, H.; Xu, W.; Zhang, W.; Mai, K. Effects of nucleotides on growth performance, immune response, disease resistance and intestinal morphology in shrimp Litopenaeus vannamei fed with a low fish meal diet. Aquac. Int. 2016, 24, 1007–1023. [Google Scholar] [CrossRef]
- Hossain, M.; Koshio, S.; Ishikawa, M.; Yokoyama, S.; Sony, N.; Fujieda, T. Nucleoside by-product dietary supplementation influences blood chemistry, immune response, oxidative stress resistance and intestinal morphology of juvenile amberjack, Seriola dumerili. Aquac. Nutr. 2017, 23, 1390–1400. [Google Scholar] [CrossRef]
- Dyachok, J.; Earnest, S.; Iturraran, E.N.; Cobb, M.H.; Ross, E.M. Amino acids regulate mTORC1 by an obligate two-step mechanism. J. Biol. Chem. 2016, 291, 22414–22426. [Google Scholar] [CrossRef]
- Xu, C.; Li, E.; Liu, Y.; Wang, S.; Wang, X.; Chen, K.; Qin, J.; Chen, L. Effect of dietary lipid level on growth, lipid metabolism and health status of the Pacific white shrimp Litopenaeus vannamei at two salinities. Aquac. Nutr. 2018, 24, 204–214. [Google Scholar] [CrossRef]
- Preiss, T.; Hentze, W.M. Starting the protein synthesis machine: Eukaryotic translation initiation. Bioessays 2003, 25, 1201–1211. [Google Scholar] [CrossRef]
- Wang, X.; Proud, C.G. The mTOR pathway in the control of protein synthesis. Physiology 2006, 21, 362–369. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formula | P | N | A | B | C | D | E |
|---|---|---|---|---|---|---|---|
| Domestic fish meal | 15.38 | 15.38 | 15.38 | 15.38 | 15.38 | 15.38 | 15.38 |
| Brown fish meal | 6.15 | 6.15 | 6.15 | 6.15 | 6.15 | 6.15 | 6.15 |
| Chicken by-product meal | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Peanut meal | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 |
| Fish by-product meal | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 |
| Shrimp meal | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 | 6.00 |
| Corn protein powder | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Cottonseed protein | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 | 4.00 |
| Soybean meal | 7.00 | 15.00 | 7.00 | 7.00 | 7.00 | 7.00 | 7.00 |
| Wheat flour | 17.20 | 17.20 | 17.20 | 17.20 | 17.20 | 17.20 | 17.20 |
| Choline chloride | 0.31 | 0.31 | 0.31 | 0.31 | 0.31 | 0.31 | 0.31 |
| Monocalcium phosphate | 1.23 | 1.23 | 1.23 | 1.23 | 1.23 | 1.23 | 1.23 |
| Calcium chloride | 0.98 | 0.98 | 0.98 | 0.98 | 0.98 | 0.98 | 0.98 |
| Shell powder | 0.62 | 0.62 | 0.62 | 0.62 | 0.62 | 0.62 | 0.62 |
| Yeast extract | 1.30 | 0 | 2.00 | 0 | 2.00 | 2.00 | 2.00 |
| Squid visceral powder | 4.80 | 0 | 2.00 | 2.00 | 0.00 | 2.00 | 2.00 |
| Fish soluble | 1.90 | 0 | 2.00 | 2.00 | 2.00 | 0 | 2.00 |
| Squid paste | 0 | 0 | 2.00 | 2.00 | 2.00 | 2.00 | 0 |
| Shrimp paste | 0 | 0 | 0 | 2.00 | 2.00 | 2.00 | 2.00 |
| Soybean phospholipid | 1.23 | 1.23 | 1.23 | 1.23 | 1.23 | 1.23 | 1.23 |
| Lysine | 0.37 | 0.37 | 0.37 | 0.37 | 0.37 | 0.37 | 0.37 |
| Additive premix1 | 9.53 | 9.53 | 9.53 | 9.53 | 9.53 | 9.53 | 9.53 |
| Proximate composition (% wet weight) | |||||||
| Crude protein | 43.00 | 42.07 | 44.64 | 45.57 | 44.37 | 43.32 | 45.11 |
| Crude lipid | 6.60 | 5.84 | 5.77 | 5.95 | 5.17 | 6.16 | 6.14 |
| Moisture | 6.78 | 6.44 | 6.44 | 6.88 | 7.35 | 6.48 | 8.19 |
| Ash | 16.13 | 15.97 | 16.41 | 16.30 | 16.36 | 15.56 | 16.13 |
| Primers | Sequences (5′-3′) | GenBank No. |
|---|---|---|
| ef-1a-R | GTATTGGAACAGTGCCCGTG | JF288785.1 |
| ef-1a-F | ACCAGGGACAGCCTCAGTAAG | |
| toll-R | CCTCGCACATCCAGGACTTTTA | / |
| toll-F | GACCATCCCTTTTACACCAGACT | |
| sod-R | CAGAGCCTTTCACTCCAACG | / |
| sod-F | GCAATGAATGCCCTTCTACC | |
| tor-R | GGGTGTTTGTGGACGGA | MN398907.1 |
| tor-F | TGCCAACGGGTGGTAGA | |
| s6k-R | CCGCCCTTGCCCAAAACCT | XM_027368997.1 |
| s6k-F | GCAAGAGGAAGACGCCATA | |
| myd88-R | GGGAGTGGCAGAAACTTATC | / |
| myd88-F | GTGCACCAGAGTCATTGTAG | |
| dorsal-R | CGTAACTTGAGGGCATCTTC | FJ998202.1 |
| dorsal-F | TGGGGAAGGAAGGATGG | |
| relish-R | GGCTGGCAAGTCGTTCTCG | EF432734 |
| relish-F | CTACATTCTGCCCTTGACTCTGG | |
| eif4a-R | GTAGTCGCCAAGAGCAAGCAC | / |
| eif4a-F | CTCCAAATCCCAAGAAACCCA | |
| eif4e2-R | GTCCTCCTGGAAGCGTA | / |
| eif4e2-F | TGGAATCAAACCTATGTGGG | |
| eif2a-R | CTAATGCCCTAAGACCATCCTG | / |
| eif2a-F | GAATAAACCTAATCGCACCACC | |
| raptor-R | TCACAATCCAAGGTCCAG | XM_027360909.1 |
| raptor-F | CTGCTTTCCAGGCTACTC |
| Groups | FBW (g) | SR (%) | FI (g) | FCR | WGR (%) | SGR (%) | CF | HSI (%) |
|---|---|---|---|---|---|---|---|---|
| P | 8.23 ± 0.14 a | 97.49 ± 0.83 | 10.05 ± 0.11 a | 1.33 ± 0.03 | 1066.04 ± 20.56 a | 4.90 ± 0.03 a | 0.64 ± 0.01 a | 4.46 ± 0.23 |
| N | 8.44 ± 0.34 ab | 93.33 ± 1.35 | 10.52 ± 0.16 ab | 1.36 ± 0.03 | 1094.04 ± 47.85 ab | 4.95 ± 0.08 ab | 0.71 ± 0.02 b | 3.97 ± 0.34 |
| A | 8.64 ± 0.11 ab | 94.16 ± 2.49 | 10.35 ± 0.24 ab | 1.30 ± 0.02 | 1125.3 ± 17.17 ab | 5.00 ± 0.02 ab | 0.61 ± 0.01 a | 4.26 ± 0.05 |
| B | 9.04 ± 0.23 b | 95.82 ± 0.83 | 10.86 ± 0.23 b | 1.30 ± 0.18 | 1181.16 ± 32.14 b | 5.09 ± 0.05 b | 0.62 ± 0.02 a | 4.57 ± 0.12 |
| C | 8.87 ± 0.09 ab | 93.33 ± 1.35 | 10.74 ± 0.14 b | 1.32 ± 0.00 | 1155.74 ± 13.77 ab | 5.05 ± 0.02 ab | 0.63 ± 0.02 a | 4.05 ± 0.08 |
| D | 9.07 ± 0.19 b | 93.33 ± 1.92 | 10.85 ± 0.10 b | 1.30 ± 0.09 | 1181.95 ± 27.59 b | 5.09 ± 0.04 b | 0.59 ± 0.01 a | 4.24 ± 0.19 |
| E | 8.48 ± 0.19 ab | 93.33 ± 1.35 | 10.52 ± 0.10 ab | 1.36 ± 0.07 | 1102.58 ± 27.01 ab | 4.97 ± 0.04 ab | 0.59 ± 0.01 a | 4.41 ± 0.39 |
| Groups | Moisture (%) | Ash (%) | Crude Lipid (%) | Crude Protein (%) |
|---|---|---|---|---|
| Body | ||||
| P | 77.35 ± 0.70 ab | 3.34 ± 0.09 | 1.70 ± 0.26 a | 16.37 ± 0.35 ab |
| N | 76.53 ± 0.27 a | 3.28 ± 0.07 | 1.64 ± 0.08 a | 17.00 ± 0.19 b |
| A | 77.08 ± 0.45 ab | 3.18 ± 0.10 | 1.87 ± 0.13 a | 16.71 ± 0.24 ab |
| B | 75.70 ± 0.24 a | 3.21 ± 0.15 | 2.37 ± 0.13 b | 17.07 ± 0.22 b |
| C | 76.64 ± 1.18 a | 3.21 ± 0.15 | 1.84 ± 0.17 a | 17.01 ± 0.97 b |
| D | 77.62 ± 0.66 ab | 3.12 ± 0.10 | 1.72 ± 0.06 a | 16.18 ± 0.29 ab |
| E | 79.04 ± 0.44 b | 3.03 ± 0.10 | 1.41 ± 0.19 a | 15.44 ± 0.24 a |
| Muscle | ||||
| P | 73.86 ± 0.81 | 0.57 ± 0.06 | 1.08 ± 0.10 | 22.02 ± 0.41 |
| N | 74.51 ± 0.82 | 0.55 ± 0.02 | 0.93 ± 0.17 | 21.83 ± 0.33 |
| A | 74.93 ± 0.38 | 0.61 ± 0.03 | 0.81 ± 0.09 | 21.88 ± 0.30 |
| B | 74.41 ± 0.29 | 0.52 ± 0.04 | 0.93 ± 0.06 | 22.13 ± 0.13 |
| C | 74.52 ± 0.65 | 0.60 ± 0.03 | 0.85 ± 0.09 | 22.26 ± 0.37 |
| D | 74.59 ± 0.18 | 0.49 ± 0.07 | 0.87 ± 0.04 | 22.23 ± 0.05 |
| E | 74.60 ± 0.18 | 0.74 ± 0.20 | 0.83 ± 0.03 | 22.45 ± 0.13 |
| Groups | FH | FW | MT |
|---|---|---|---|
| P | 43.55 + 5.51 | 50.56 + 9.02 | 66.86 + 4.32 ab |
| N | 33.89 + 5.34 | 39.70 + 2.35 | 62.54 + 7.83 ab |
| A | 37.60 + 6.88 | 37.33 + 6.52 | 65.33 + 3.59 ab |
| B | 43.17 + 2.28 | 37.97 + 6.71 | 60.72 + 3.24 ab |
| C | 50.52 + 11.94 | 47.21 + 5.43 | 74.21 + 6.31 b |
| D | 36.44 + 4.17 | 38.01 + 2.62 | 58.56 + 2.20 ab |
| E | 36.03 + 5.60 | 32.33 + 5.74 | 56.63 + 3.75 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
He, G.; Chen, X.; Zeng, Q.; Zhu, W.; Chen, Z.; Tan, B.; Xie, S. Effects of Compound Feed Attractants on Growth Performance, Feed Utilization, Intestinal Histology, Protein Synthesis, and Immune Response of White Shrimp (Litopenaeus Vannamei). Animals 2022, 12, 2550. https://doi.org/10.3390/ani12192550
He G, Chen X, Zeng Q, Zhu W, Chen Z, Tan B, Xie S. Effects of Compound Feed Attractants on Growth Performance, Feed Utilization, Intestinal Histology, Protein Synthesis, and Immune Response of White Shrimp (Litopenaeus Vannamei). Animals. 2022; 12(19):2550. https://doi.org/10.3390/ani12192550
Chicago/Turabian StyleHe, Guilun, Xin Chen, Qingtian Zeng, Wenbo Zhu, Zhengbang Chen, Beiping Tan, and Shiwei Xie. 2022. "Effects of Compound Feed Attractants on Growth Performance, Feed Utilization, Intestinal Histology, Protein Synthesis, and Immune Response of White Shrimp (Litopenaeus Vannamei)" Animals 12, no. 19: 2550. https://doi.org/10.3390/ani12192550
APA StyleHe, G., Chen, X., Zeng, Q., Zhu, W., Chen, Z., Tan, B., & Xie, S. (2022). Effects of Compound Feed Attractants on Growth Performance, Feed Utilization, Intestinal Histology, Protein Synthesis, and Immune Response of White Shrimp (Litopenaeus Vannamei). Animals, 12(19), 2550. https://doi.org/10.3390/ani12192550