


Host–Parasite Relationship—Nematode Communities in Populations of Small Mammals
 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Parasitological Techniques
2.2. Statistical Approach
3. Results
3.1. Parasites Diversity and Parasite Loads
3.2. Relationships between Parasite Load and Morphological Traits
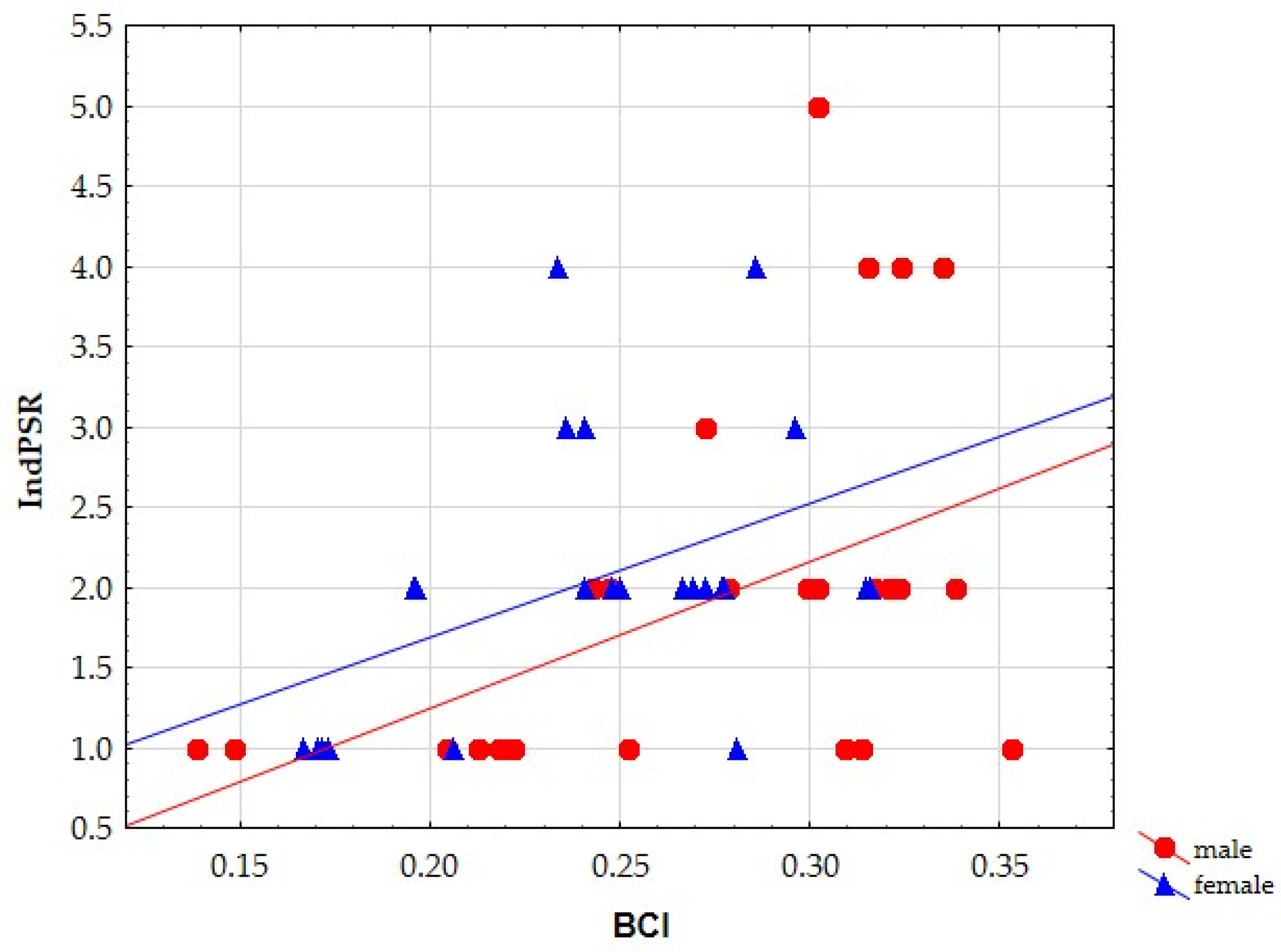
3.2.1. Body Condition Index (BCI) vs. Parasitological Parameters
3.2.2. Spleen Mass vs. Parasitological Parameters
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Byers, J.E.; Schmidt, J.P.; Pappalardo, P.; Haas, S.E.; Stephens, P.R. What factors explain the geographical range of mammalian parasites? Proc. R. Soc. B 2019, 286, 20190673. [Google Scholar] [CrossRef] [PubMed]
- Bordes, F.; Morand, S. The impact of multiple infections on wild animal hosts: A review. Infect. Ecol. Epidemiol. 2011, 1, 7346. [Google Scholar] [CrossRef] [PubMed]
- Morand, S.; Poulin, R. Density, body mass and parasite species richness of terrestrial mammals. Evol. Ecol. 1998, 12, 717–727. [Google Scholar] [CrossRef]
- Poulin, R. Phylogeny, ecology, and the richness of parasite communities in vertebrates: Ecological archives M065-001. Ecol. Monogr. 1995, 65, 283–302. [Google Scholar] [CrossRef]
- Feliu, C.; Renaud, F.; Catzeflis, F.; Hugot, J.-P.; Durand, P.; Morand, S. A comparative analysis of parasite species richness of Iberian rodents. Parasitology 1997, 115, 453–466. [Google Scholar] [CrossRef]
- Nunn, C.L.; Altizer, S.; Jones, K.E.; Sechrest, W. Comparative tests of parasite species richness in primates. Am. Nat. 2003, 162, 597–614. [Google Scholar] [CrossRef]
- Krasnov, B.R.; Shenbrot, G.I.; Khokhlova, I.S.; Degen, A.A. Flea species richness and parameters of host body, host geography and host ‘milieu’. J. Anim. Ecol. 2004, 73, 1121–1128. [Google Scholar] [CrossRef]
- Korallo, N.P.; Vinarski, M.V.; Krasnov, B.R.; Shenbrot, G.I.; Mouillot, D.; Poulin, R. Are there general rules governing parasite diversity? Small mammalian hosts and gamasid mite assemblages. Divers. Distrib. 2007, 13, 353–360. [Google Scholar] [CrossRef]
- Gregory, R.D.; Keymer, A.E.; Harvey, P.H. Helminth parasite richness among vertebrates. Biodivers. Conserv. 1996, 5, 985–997. [Google Scholar] [CrossRef]
- Ezenwa, V.O.; Price, S.A.; Altizer, S.; Vitone, N.D.; Cook, K.C. Host traits and parasite species richness in even and odd-toed hoofed mammals, artiodactyla and perissodactyla. Oikos 2006, 115, 526–536. [Google Scholar] [CrossRef]
- Bordes, F.; Morand, S.; Ricardo, G. Bat fly species richness in neotropical bats: Correlations with host ecology and host brain. Oecologia 2008, 158, 109–116. [Google Scholar] [CrossRef] [PubMed]
- Morand, S. (Macro-) Evolutionary ecology of parasite diversity: From determinants of parasite species richness to host diversification. Int. J. Parasitol. Parasites Wildl. 2015, 4, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Sánchez, C.A.; Becker, D.J.; Teitelbaum, C.S.; Barriga, P.; Brown, L.M.; Majewska, A.A.; Hall, R.J.; Altizer, S. On the relationship between body condition and parasite infection in wildlife: A review and meta-analysis. Ecol. Lett. 2018, 21, 1869–1884. [Google Scholar] [CrossRef]
- Dobson, F.S. Body mass, structural size, and life-history patterns of the Columbian ground squirrel. Am. Nat. 1992, 140, 109–125. [Google Scholar] [CrossRef] [PubMed]
- Schulte-Hostedde, A.I.; Millar, J.S.; Hickling, G.J. Evaluating body condition in small mammals. Can. J. Zool. 2001, 79, 1021–1029. [Google Scholar] [CrossRef]
- Wauters, L.A.; Dhondt, A.A. Lifetime reproductive success and its correlates in female Eurasian red squirrels. Oikos 1995, 72, 402. [Google Scholar] [CrossRef]
- Maceda-Veiga, A.; Green, A.J.; Poulin, R.; Lagrue, C. Body condition peaks at intermediate parasite loads in the common bully Gobiomorphus cotidianus. PLoS ONE 2016, 11, e0168992. [Google Scholar] [CrossRef]
- Bordes, F.; Ponlet, N.; de Bellocq, J.G.; Ribas, A.; Krasnov, B.R.; Morand, S. Is there sex-biased resistance and tolerance in Mediterranean wood mouse (Apodemus sylvaticus) populations facing multiple helminth infections? Oecologia 2012, 170, 123–135. [Google Scholar] [CrossRef]
- Stradiotto, A.; Cagnacci, F.; Delahay, R.; Tioli, S.; Nieder, L.; Rizzoli, A. Spatial organization of the yellow-necked mouse: Effects of density and resource availability. J. Mammal. 2009, 90, 704–714. [Google Scholar] [CrossRef]
- Ostfeld, R.; Glass, G.; Keesing, F. Spatial Epidemiology: An emerging (or re-emerging) discipline. Trends Ecol. Evol. 2005, 20, 328–336. [Google Scholar] [CrossRef]
- Khokhlova, I.S.; Serobyan, V.; Degen, A.A.; Krasnov, B.R. Discrimination of host sex by a haematophagous ectoparasite. Anim. Behav. 2011, 81, 275–281. [Google Scholar] [CrossRef]
- Kowalski, K.; Bogdziewicz, M.; Eichert, U.; Rychlik, L. Sex differences in flea infections among rodent hosts: Is there a male bias? Parasitol. Res. 2015, 114, 337–341. [Google Scholar] [CrossRef] [PubMed]
- Rolff, J. Bateman’s principle and immunity. Proc. R. Soc. Lond. B 2002, 269, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Ezenwa, V.O.; Stefan Ekernas, L.; Creel, S. Unravelling complex associations between testosterone and parasite infection in the wild. Func. Ecol. 2012, 26, 123–133. [Google Scholar] [CrossRef]
- Behnke, J.M.; Lewis, J.W.; Zain, S.N.M.; Gilbert, F.S. Helminth infections in Apodemus sylvaticus in southern England: Interactive effects of host age, sex and year on the prevalence and abundance of infections. J. Helminthol. 1999, 73, 31–44. [Google Scholar] [CrossRef] [PubMed]
- Fuentes, M.V.; Fuentes, M.V.; Sáez, S.; Trelis, M.; Galán-Puchades, M.T.; Esteban, J.G. The helminth community of the wood mouse, Apodemus sylvaticus, in the Sierra Espuña, Murcia, Spain. J. Helminthol. 2004, 78, 219–223. [Google Scholar] [CrossRef] [PubMed]
- Debenedetti, Á.L.; Sainz-Elipe, S.; Sáez-Durán, S.; Galicia, D.; Imaz, A.; Galán-Puchades, M.T.; Fuentes, M.V. First report on the helminthfauna of the yellow-necked mouse, Apodemus flavicollis, in the Iberian Peninsula. Helminthologia 2016, 53, 294–297. [Google Scholar] [CrossRef]
- Abt, K.F.; Bock, W.F. Seasonal Variations of diet composition in farmland field mice Apodemus spp. and bank voles Clethrionomys glareolus. Acta Theriol. 1998, 43, 379–389. [Google Scholar] [CrossRef]
- Goüy de Bellocq, J.; Sarà, M.; Casanova, J.C.; Feliu, C.; Morand, S. A Comparison of the structure of helminth communities in the woodmouse, Apodemus sylvaticus, on islands of the western Mediterranean and continental Europe. Parasitol. Res. 2003, 90, 64–70. [Google Scholar] [CrossRef]
- Eira, C.; Torres, J.; Vingada, J.; Miquel, J. Ecological Aspects influencing the helminth community of the wood mouse Apodemus sylvaticus in Dunas de Mira, Portugal. Acta Parasitol. 2006, 51, 300–308. [Google Scholar] [CrossRef]
- Milazzo, C.; Casanova, J.C.; Aloise, G.; Ribas, A.; Cagnin, M. Helminths of the bank vole Clethrionomys glareolus (Rodentia, Arvicolinae) in Southern Italy. Ital. J. Zool. 2003, 70, 333–337. [Google Scholar] [CrossRef]
- Klimpel, S.; Förster, M.; Schmahl, G. Parasite Fauna of the bank vole (Clethrionomys glareolus) in an urban region of germany: Reservoir host of zoonotic metazoan parasites? Parasitol. Res. 2007, 102, 69–75. [Google Scholar] [CrossRef] [PubMed]
- Ondríková, J.; Miklisová, D.; Ribas, A.; Stanko, M. The helminth parasites of two sympatric species of the genus Apodemus (Rodentia, Muridae) from South-Eastern Slovakia. Acta Parasitol. 2010, 55, 369–378. [Google Scholar] [CrossRef]
- Bužan, E.; Čabrilo, B.; Ivović, V.; Bjelić Čabrilo, O. Environmental effects of anthropogenic waste on intestinal nematode parasites of murid rodents in Istria, Croatia. Biol. Serb. 2019, 41, 78–88. [Google Scholar] [CrossRef]
- Bjelić Čabrilo, O.; Kostić, D.; Popović, E.; Ćirković, M.; Aleksić, N.; Lujić, J. Helminth fauna of the bank vole Myodes glareolus (Rodentia, Arvicolinae) on the territory of Fruska Gora mountain (Serbia)—A potential source of zoonoses. Bulg. J. Agric. Sci. 2011, 17, 829–836. [Google Scholar]
- Bjelić-Čabrilo, O.; Čabrilo, B.; Popović, E. Helminth Fauna of Rodents (Mammalia, Rodentia) from Zasavica (Serbia). Biol. Serb. 2014, 35, 43–47. [Google Scholar]
- Čabrilo, B.; Jovanović, V.M.; Bjelić-Čabrilo, O.; Budinski, I.; Blagojević, J.; Vujošević, M. Diversity of nematodes in the yellow-necked field mouse Apodemus flavicollis from the Peripannonic region of Serbia. J. Helminthol. 2016, 90, 14–20. [Google Scholar] [CrossRef]
- Čabrilo, B.; Jovanović, V.M.; Čabrilo, O.B.; Budinski, I.; Blagojević, J.; Vujošević, M. Is there a host sex bias in intestinal nematode parasitism of the yellow-necked mouse (Apodemus flavicollis) at obedska bara pond, Serbia? Helminthologia 2018, 55, 247–250. [Google Scholar] [CrossRef]
- Jovanović, V.M.; Čabrilo, B.; Budinski, I.; Bjelić-Čabrilo, O.; Adnađević, T.; Blagojević, J.; Vujošević, M. Host B chromosomes as potential sex ratio distorters of intestinal nematode infrapopulations in the yellow-necked mouse (Apodemus flavicollis). J. Helminthol. 2019, 93, 552–558. [Google Scholar] [CrossRef]
- Adnađević, T.; Jovanović, V.M.; Blagojević, J.; Budinski, I.; Čabrilo, B.; Bijelić-Čabrilo, O.; Vujošević, M. Data from: Possible influence of B chromosomes on genes included in immune response and parasite burden in Apodemus flavicollis. PLoS ONE 2015, 11, e112260. [Google Scholar]
- Kataranovski, M.; Mirkov, I.; Zolotarevski, L.; Popov, A.; Belij, S.; Stosic, J.; Kataranovski, D. Basic indices of spleen immune activity in natural populations of norway rats (Rattus norvegicus Berkenhout, 1769) in Serbia. Arch. Biol. Sci. 2009, 61, 723–732. [Google Scholar] [CrossRef]
- Ponlet, N.; Chaisiri, K.; Claude, J.; Morand, S. Incorporating parasite systematics in comparative analyses of variation in spleen mass and testes sizes of rodents. Parasitology 2011, 138, 1804–1814. [Google Scholar] [CrossRef] [PubMed]
- Hofmeester, T.R.; Bügel, E.J.; Hendrikx, B.; Maas, M.; Franssen, F.F.J.; Sprong, H.; Matson, K.D. Parasite load and site-specific parasite pressure as determinants of immune indices in two sympatric rodent species. Animals 2019, 9, 1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shutler, D.; Alisauskas, R.T.; McLaughlin, J.D. Mass dynamics of the spleen and other organs in geese: Measures of immune relationships to helminths? Can. J. Zool. 1999, 77, 351–359. [Google Scholar] [CrossRef]
- Hõrak, P.; Tummeleht, L.; Talvik, H. Predictors and markers of resistance to neurotropic nematode infection in rodent host. Parasitol. Res. 2006, 98, 396–402. [Google Scholar] [CrossRef]
- Cowan, K.M.; Shutler, D.; Herman, T.B.; Stewart, D.T. Splenic mass of masked shrews, sorex cinereus, in relation to body mass, sex, age, day of the year, and bladder nematode, Liniscus (=Capillaria) Maseri, infection. J. Parasitol. 2009, 95, 228–230. [Google Scholar] [CrossRef]
- Nunn, C.L. Spleen size, disease risk and sexual selection: A comparative study in primates. Evol. Ecol. Res. 2002, 4, 91–107. [Google Scholar]
- Morand, S.; Poulin, R. Nematode parasite species richness and the evolution of spleen size in birds. Can. J. Zool. 2000, 78, 1356–1360. [Google Scholar] [CrossRef]
- Schulte-Hostedde, A.I.; Elsasser, S.C. Spleen mass, body condition, and parasite load in male american mink (Neovison vison). J. Mammal. 2011, 92, 221–226. [Google Scholar] [CrossRef]
- Ryžikov, K. Key to Rodent Helminth Fauna of the USSR. Tapeworms and Flukes; Izdatelstvo ‘Nauka: Moscow, Soviet Union, 1978. [Google Scholar]
- Ryžikov, K. Key to Rodent Helminth Fauna of the USSR. Roundworms and Acanthocephalans; Izdatelstvo ‘Nauka: Moscow, Soviet Union, 1979. [Google Scholar]
- Genov, T. Helminths of Insectivorous Mammals and Rodents in Bulgaria; Publishing House of the Bulgarian Academy of Sciences: Sofia, Bulgaria, 1984; 348p. [Google Scholar]
- Bush, A.O.; Lafferty, K.D.; Lotz, J.M.; Shostak, A.W. Parasitology meets ecology on its own terms: Margolis et al. Revisited. J. Parasitol. 1997, 83, 575. [Google Scholar] [CrossRef]
- Dobson, A.; Lafferty, K.D.; Kuris, A.M.; Hechinger, R.F.; Jetz, W. Homage to Linnaeus: How many Parasites? How many Hosts? Proc. Nat. Acad. Sci. USA 2008, 105, 11482–11489. [Google Scholar] [CrossRef] [PubMed]
- Čabrilo, B. Diversity and Ecology of Intestinal Nematodes of the Yellow-Necked Mouse (Apodemus flavicollis Melchior, 1834) on the Territory of Serbia. Ph.D. Thesis, Department of Biology and Ecology, Faculty of Sciences, Novi Sad, Serbia, 2017. [Google Scholar]
- Milazzo, C.; Di Bella, C.; Casanova, J.C.; Ribas, A.; Cagnin, M. Helminth communities of wood mouse (Apodemus sylvaticus) on the River Avena (Calabria, Southern Italy). Hystrix Ital. J. Mamm. 2010, 21, 171–176. [Google Scholar] [CrossRef]
- Sáez-Durán, S.; Debenedetti, Á.L.; Sainz-Elipe, S.; Sabater-Tena, M.; Galán-Puchades, M.T.; Fuentes, M.V. Ecological analysis of the helminth community of the wood mouse, Apodemus sylvaticus, along an 18-year post-fire regeneration period in a Mediterranean ecosystem. Animals 2021, 11, 2926. [Google Scholar] [CrossRef]
- Gryczyńska-Siemiątkowska, A.; Gortat, T.; Kozakiewicz, A.; Rutkowski, R.; Pomorski, J.; Kozakiewicz, M. Multiple paternity in a wild population of the yellow-necked mouse Apodemus flavicollis. Acta Theriol. 2008, 53, 251–258. [Google Scholar] [CrossRef]
- Wolff, J.O.; Macdonald, D.W. Promiscuous females protect their offspring. Trends Ecol. Evol. 2004, 19, 127–134. [Google Scholar] [CrossRef]
- Baker, R.J.; Makova, K.D.; Chesser, R.K. Microsatellites indicate a high frequency of multiple paternity in Apodemus (Rodentia). Mol. Ecol. 1999, 8, 107–111. [Google Scholar] [CrossRef]
- Booth, W.; Montgomery, W.I.; Prodöhl, P.A. Polyandry by wood mice in natural populations. J. Zool. 2007, 273, 176–182. [Google Scholar] [CrossRef]
- Klemme, I.; Eccard, J.A.; Ylönen, H. Do female bank voles (Clethrionomys glareolus) mate multiply to improve on previous mates? Behav. Ecol. Sociobiol. 2006, 60, 415–421. [Google Scholar] [CrossRef]
- Bryja, J.; Stopka, P. Facultative Promiscuity in a presumably monogamous mouse Apodemus microps. Acta Theriol. 2005, 50, 189–196. [Google Scholar] [CrossRef]
- Clutton-Brock, T.H. The Evolution of Parental Care; Princeton University Press: Princeton, NJ, USA, 1991; 352p. [Google Scholar]
- Lonstein, J.S.; De Vries, G.J. Sex differences in the parental behavior of rodents. Neurosci. Biobehav. Rev. 2000, 24, 669–686. [Google Scholar] [CrossRef]
- Girard, I.; Swallow, J.G.; Carter, P.A.; Koteja, P.; Rhodes, J.S.; Garland, T. Maternal-care behavior and life-history traits in house mice (Mus domesticus) artificially selected for high voluntary wheel-running activity. Behav. Processes 2002, 57, 37–50. [Google Scholar] [CrossRef]
- Kiffner, C.; Stanko, M.; Morand, S.; Khokhlova, I.S.; Shenbrot, G.I.; Laudisoit, A.; Leirs, H.; Hawlena, H.; Krasnov, B.R. Sex-biased parasitism is not universal: Evidence from rodent–flea associations from three biomes. Oecologia 2013, 173, 1009–1022. [Google Scholar] [CrossRef] [PubMed]
- Krasnov, B.R.; Morand, S.; Hawlena, H.; Khokhlova, I.S.; Shenbrot, G.I. Sex-biased parasitism, seasonality and sexual size dimorphism in desert rodents. Oecologia 2005, 146, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Patterson, B.D.; Dick, C.W.; Dittmar, K. Sex biases in parasitism of neotropical bats by bat flies (Diptera: Streblidae). J. Trop. Ecol. 2008, 24, 387–396. [Google Scholar] [CrossRef]
- Vicente, J.; Pérez-Rodríguez, L.; Gortazar, C. Sex, age, spleen size, and kidney fat of red deer relative to infection intensities of the lungworm Elaphostrongylus cervi. Naturwissenschaften 2007, 94, 581–587. [Google Scholar] [CrossRef]
- Fumagalli, M.; Pozzoli, U.; Cagliani, R.; Comi, G.P.; Riva, S.; Clerici, M.; Bresolin, N.; Sironi, M. Parasites represent a major selective force for interleukin genes and shape the genetic predisposition to autoimmune conditions. J. Exp. Med. 2009, 206, 1395–1408. [Google Scholar] [CrossRef]
- Fumagalli, M.; Pozzoli, U.; Cagliani, R.; Comi, G.P.; Bresolin, N.; Clerici, M.; Sironi, M. The landscape of human genes involved in the immune response to parasitic worms. BMC Evol. Biol. 2010, 10, 264. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Localities | A. sylvaticus | A. flavicolis | M. glareolus | Total |
|---|---|---|---|---|
| Zvezdarska šuma (Belgrade) | 18 | - | - | 18 |
| Ravnište (Jastrebac Mt.) | - | 6 | 8 | 14 |
| Vlasina | - | 15 | 5 | 20 |
| Babin zub (Stara planina Mt) | - | 2 | 4 | 6 |
| Petnica | - | 12 | 4 | 12 |
| Goč Mt. | - | 5 | - | 9 |
| Maljen Mt. | - | 4 | - | 4 |
| Ruski Krstur | - | 5 | - | 5 |
| Total | 18 | 49 | 21 | 88 |
| Parasites/Hosts | Apodemus flavicollis | Apodemus sylvaticus | Myodes glareolus | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| n | P% | MI | MA | n | P% | MI | MA | n | P% | MI | MA | |
| Aonchotheca annulosa | 120 | 10.2 | 24 | 2.45 | 4 | 11.1 | 2 | 0.22 | 25 | 14.3 | 8.33 | 1.19 |
| Aonchotheca murissylvatici | - | - | - | - | - | - | - | - | 70 | 9.5 | 35 | 3.33 |
| Aspiculuris tetraptera | 2 | 4.1 | 1 | 0.04 | - | - | - | - | - | - | - | - |
| Eucoleus sp. | 23 | 10.2 | 4.60 | 0.47 | - | - | - | - | - | - | - | - |
| Heligmosomoides sp. | - | - | - | - | - | - | - | - | 65 | 23.8 | 13 | 3.1 |
| Heligmosomum costellatum | - | - | - | - | - | - | - | - | 13 | 19.05 | 3.25 | 0.62 |
| Heligmosomoides glareoli | 1 | 2.0 | 1 | 0.02 | - | - | - | - | 56 | 66.7 | 4 | 2.67 |
| Heligmosomoides polygyrus | 357 | 69.4 | 10.50 | 7.29 | 10 | 33.3 | 1.67 | 0.56 | 2 | 4.7 | 2 | 0.10 |
| Heterakis spumosa | 1 | 2.0 | 1 | 0.02 | - | - | - | - | - | - | - | - |
| Mastophorus muris | 5 | 4.1 | 2.5 | 0.1 | - | - | - | - | - | - | - | - |
| Rictularia proni | 14 | 10.2 | 2.8 | 0.29 | - | - | - | - | - | - | - | - |
| Syphacia sp. | 1 | 2.0 | 1 | 0.02 | 10 | 16.7 | 3.33 | 0.56 | - | - | - | - |
| Syphacia frederici | 229 | 26.5 | 17.62 | 4.67 | 369 | 22.2 | 92.25 | 20.5 | - | - | - | - |
| Syphacia stroma | 702 | 28.6 | 50.14 | 14.33 | - | - | - | - | - | - | - | - |
| Trichuris muris | 532 | 28.6 | 38 | 10.86 | - | - | - | - | 1 | 4.8 | 1 | 0.05 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miljević, M.; Čabrilo, B.; Budinski, I.; Rajičić, M.; Bajić, B.; Bjelić-Čabrilo, O.; Blagojević, J. Host–Parasite Relationship—Nematode Communities in Populations of Small Mammals. Animals 2022, 12, 2617. https://doi.org/10.3390/ani12192617
Miljević M, Čabrilo B, Budinski I, Rajičić M, Bajić B, Bjelić-Čabrilo O, Blagojević J. Host–Parasite Relationship—Nematode Communities in Populations of Small Mammals. Animals. 2022; 12(19):2617. https://doi.org/10.3390/ani12192617
Chicago/Turabian StyleMiljević, Milan, Borislav Čabrilo, Ivana Budinski, Marija Rajičić, Branka Bajić, Olivera Bjelić-Čabrilo, and Jelena Blagojević. 2022. "Host–Parasite Relationship—Nematode Communities in Populations of Small Mammals" Animals 12, no. 19: 2617. https://doi.org/10.3390/ani12192617
APA StyleMiljević, M., Čabrilo, B., Budinski, I., Rajičić, M., Bajić, B., Bjelić-Čabrilo, O., & Blagojević, J. (2022). Host–Parasite Relationship—Nematode Communities in Populations of Small Mammals. Animals, 12(19), 2617. https://doi.org/10.3390/ani12192617