gga-miR-449b-5p Regulates Steroid Hormone Synthesis in Laying Hen Ovarian Granulosa Cells by Targeting the IGF2BP3 Gene

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Sample Collection
2.3. Cell Culture
2.4. RNA Extraction, cDNA Synthesis, and Quantitative Real-Time PCR (qRT-PCR)
2.5. Plasmid Construction
2.6. Cell Transfection and Treatment
2.7. Cell Proliferation Assay
2.8. Flow Cytometric Analysis
2.9. ELISA for Steroid Hormones
2.10. Western Blotting Assay
2.11. Dual-Luciferase Reporter Assay
2.12. Statistical Analysis
3. Results
3.1. Differential Expression of gga-miR-449b-5p in TCs and GCs at All Levels
3.2. gga-miR-449b-5p Has No Effect on the Proliferation of GCs
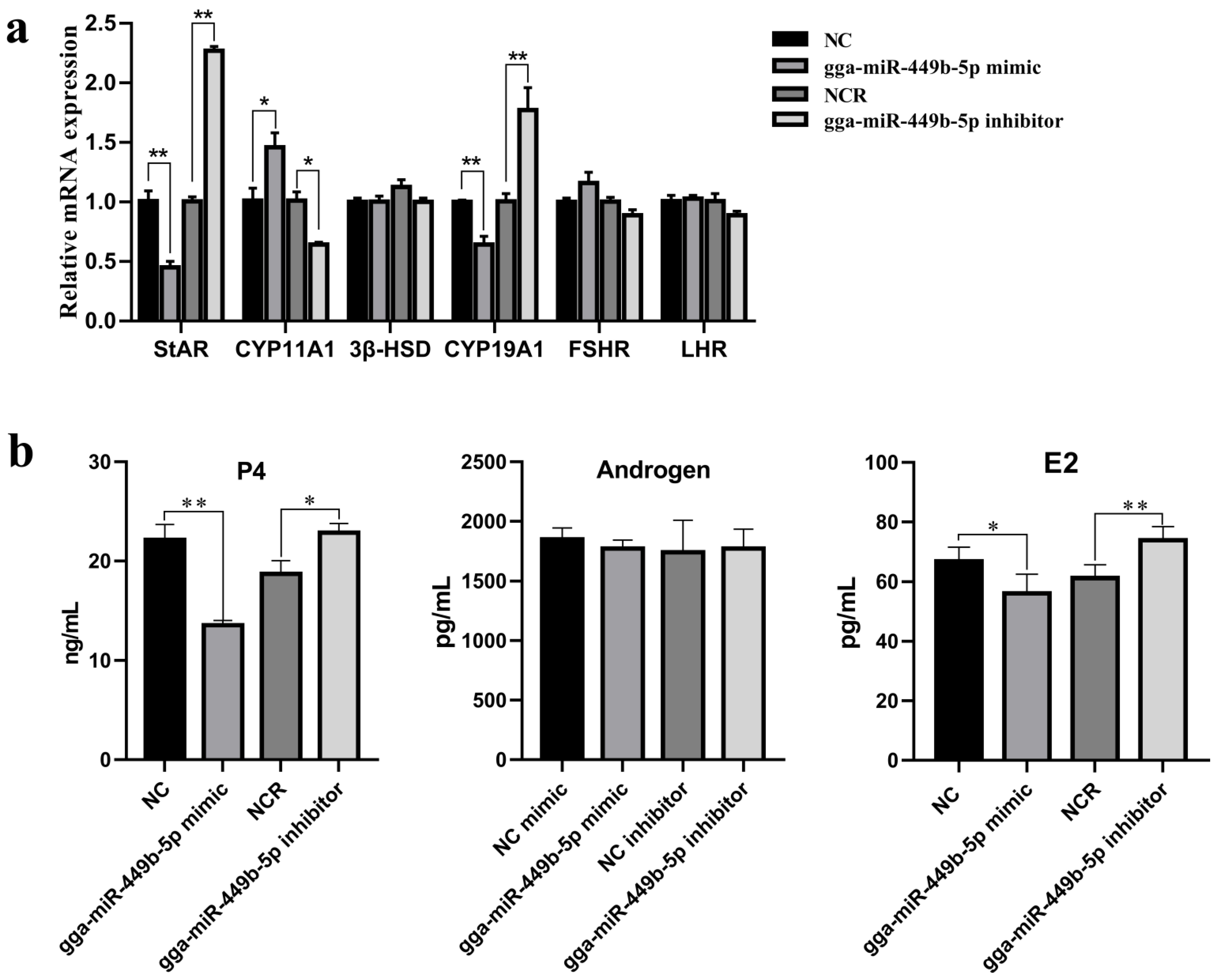
3.3. gga-miR-449b-5p Regulates Steroid Secretion by GCs
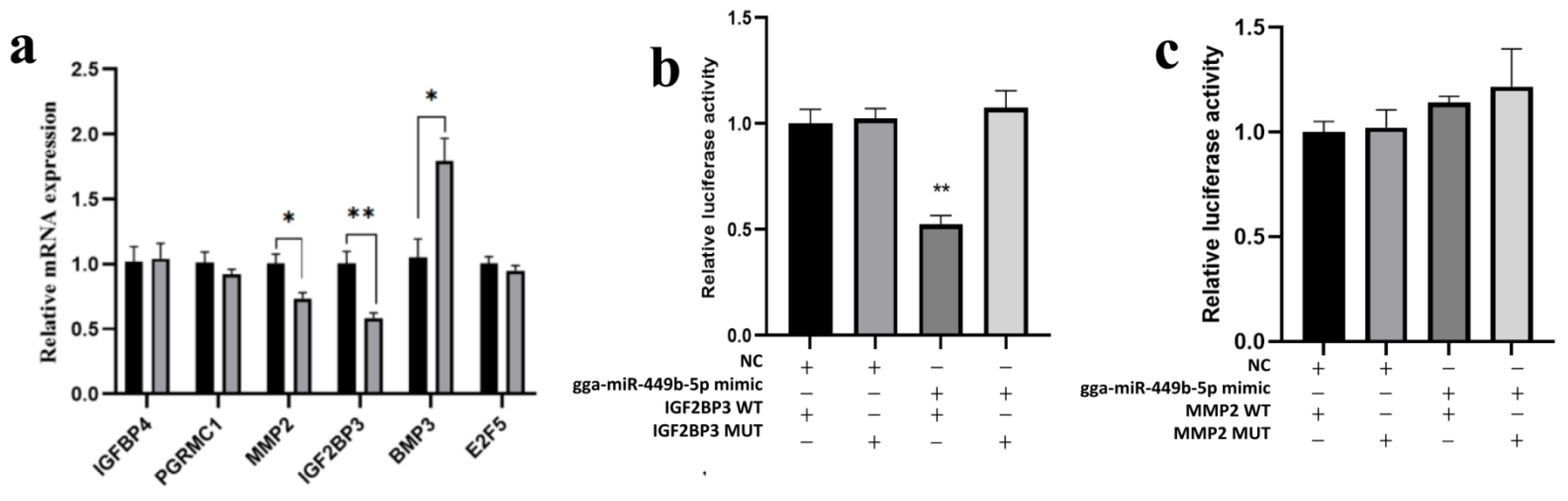
3.4. IGF2BP3 Is a gga-miR-449b-5p Target
3.5. Expression of IGF2BP3 Is Regulated by gga-miR-449b-5p
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
References
- Johnson, A.L. Reproduction in the Female. Sturkie’s Avian Physiology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 635–665. [Google Scholar]
- Bahr, J.M. The chicken ovary as a model of follicular development. In Seminars in Reproductive Endocrinology; Thieme Medical Publishers, Inc.: New York, NY, USA, 1991; pp. 352–359. [Google Scholar]
- Nitta, H.; Osawa, Y.; Bahr, J. Immunolocalization of steroidogenic cells in small follicles of the chicken ovary: Anatomical arrangement and location of steroidogenic cells change during follicular development. Domest. Anim. Endocrinol. 1991, 8, 587–594. [Google Scholar] [CrossRef]
- Sechman, A.; Łakota, P.; Wojtysiak, D.; Hrabia, A.; Mika, M.; Lisowski, M.; Czekalski, P.; Rza˛sa, J.; Kapkowska, E.; Bednarczyk, M. Sex steroids level in blood plasma and ovarian follicles of the chimeric chicken. J. Vet. Med. Ser. A 2006, 53, 501–508. [Google Scholar] [CrossRef] [PubMed]
- Yoshimura, Y.; Okamoto, T.; Tamura, T. Changes in localization of ovarian immunoreactive estrogen receptor during follicular development in hens. Gen. Comp. Endocrinol. 1995, 100, 368–374. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Shimada, K.; Saito, N.; Kansaku, N. Expression of messenger ribonucleic acids of luteinizing hormone and follicle-stimulating hormone receptors in granulosa and theca layers of chicken preovulatory follicles. Gen. Comp. Endocrinol. 1997, 105, 402–409. [Google Scholar] [CrossRef] [PubMed]
- Yamamura, N.; Takeishi, M.; Goto, N.; Tagami, M.; Mizutani, T.; Miyamoto, K.; Doi, O.; Kamiyoshi, M. Expression of messenger RNAs of luteinizing hormone and follicle-stimulating hormone receptors in the granulosa layer during the ovulatory cycle of the hen. Br. Poult. Sci. 2000, 41 (Suppl. 1), 71–72. [Google Scholar] [CrossRef]
- Du, Y.; Liu, L.; He, Y.; Dou, T.; Jia, J.; Ge, C.J.B.P.S. Endocrine and genetic factors affecting egg laying performance in chickens: A review. Br. Poult. Sci. 2020, 61, 538–549. [Google Scholar] [CrossRef]
- Wang, J.; Gong, Y. Transcription of CYP19A1 is directly regulated by SF-1 in the theca cells of ovary follicles in chicken. Gen. Comp. Endocrinol. 2017, 247, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Jing, R.; Gu, L.; Li, J.; Gong, Y. A transcriptomic comparison of theca and granulosa cells in chicken and cattle follicles reveals ESR2 as a potential regulator of CYP19A1 expression in the theca cells of chicken follicles. Comp. Biochem. Physiol. Part D Genom. Proteom. 2018, 27, 40–53. [Google Scholar] [CrossRef]
- Johnson, A.L. The avian ovary and follicle development: Some comparative and practical insights. Turk. J. Vet. Anim. Sci. 2014, 38, 660–669. [Google Scholar] [CrossRef]
- Nitta, H.; Ian Mason, J.; Bahr, J.M. Localization of 3β-hydroxysteroid dehydrogenase in the chicken ovarian follicle shifts from the theca layer to granulosa layer with follicular maturation. Biol. Reprod. 1993, 48, 110–116. [Google Scholar] [CrossRef]
- Sato, Y.; Suzuki, T.; Hidaka, K.; Sato, H.; Ito, K.; Ito, S.; Sasano, H. Immunolocalization of nuclear transcription factors, DAX-1 and COUP-TF II, in the normal human ovary: Correlation with adrenal 4 binding protein/steroidogenic factor-1 immunolocalization during the menstrual cycle. J. Clin. Endocrinol. Metab. 2003, 88, 3415–3420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bennett, J.; Baumgarten, S.C.; Stocco, C. GATA4 and GATA6 silencing in ovarian granulosa cells affects levels of mRNAs involved in steroidogenesis, extracellular structure organization, IGF-I activity, and apoptosis. Endocrinology 2013, 154, 4845–4858. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amsterdam, A.; Keren-Tal, I.; Aharoni, D.; Dantes, A.; Land-Bracha, A.; Rimon, E.; Sasson, R.; Hirsh, L. Steroidogenesis and apoptosis in the mammalian ovary. Steroids 2003, 68, 861–867. [Google Scholar] [CrossRef] [PubMed]
- Wang, W.; Wu, K.; Jia, M.; Sun, S.; Kang, L.; Zhang, Q.; Tang, H. Dynamic Changes in the Global MicroRNAome and Transcriptome Identify Key Nodes Associated with Ovarian Development in Chickens. Front. Genet. 2018, 9, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ocłoń, E.; Hrabia, A. miRNA expression profile in chicken ovarian follicles throughout development and miRNA-mediated MMP expression. Theriogenology 2021, 160, 116–127. [Google Scholar] [CrossRef]
- Kang, L.; Cui, X.; Zhang, Y.; Yang, C.; Jiang, Y. Identification of miRNAs associated with sexual maturity in chicken ovary by Illumina small RNA deep sequencing. BMC Genom. 2013, 14, 352. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Chang, L.; Wang, Y.; Wang, R.; Hu, L.; Zhao, Z.; Geng, L.; Liu, Z.; Gong, Y.; Li, J.; et al. Genome-wide differential expression of long noncoding RNAs and mRNAs in ovarian follicles of two different chicken breeds. Genomics 2019, 111, 1395–1403. [Google Scholar] [CrossRef]
- Li, J.; Si, S.-J.; Wu, X.; Zhang, Z.; Li, C.; Tao, Y.; Yang, P.; Li, D.; Li, G.; Liu, X.; et al. CircEML1 facilitates the steroid synthesis in follicular granulosa cells of chicken through sponging gga-miR-449a to release IGF2BP3 expression. bioRxiv 2022. [Google Scholar]
- Stark, A.; Brennecke, J.; Bushati, N.; Russell, R.B.; Cohen, S.M. Animal MicroRNAs confer robustness to gene expression and have a significant impact on 3′ UTR evolution. Cell 2005, 123, 1133–1146. [Google Scholar] [CrossRef] [Green Version]
- Kehl, T.; Backes, C.; Kern, F.; Fehlmann, T.; Ludwig, N.; Meese, E.; Lenhof, H.; Keller, A. About miRNAs, miRNA seeds, target genes and target pathways. Oncotarget 2017, 8, 107167. [Google Scholar] [CrossRef] [Green Version]
- Bushati, N.; Cohen, S.M. microRNA functions. Annu. Rev. Cell. Dev. Biol. 2007, 23, 175–205. [Google Scholar] [CrossRef]
- Reddy, K.B. MicroRNA (miRNA) in cancer. Cancer Cell Int. 2015, 15, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cai, Y.; Yu, X.; Hu, S.; Yu, J. A brief review on the mechanisms of miRNA regulation. Genom. Proteom. Bioinform. 2009, 7, 147–154. [Google Scholar] [CrossRef] [Green Version]
- Gebremedhn, S.; Salilew-Wondim, D.; Ahmad, I.; Sahadevan, S.; Hossain, M.M.; Hoelker, M.; Rings, F.; Neuhoff, C.; Tholen, E.; Looft, C.; et al. MicroRNA expression profile in bovine granulosa cells of preovulatory dominant and subordinate follicles during the late follicular phase of the estrous cycle. PLoS ONE 2015, 10, e0125912. [Google Scholar] [CrossRef] [PubMed]
- Maalouf, S.; Liu, W.; Pate, J.L. MicroRNA in ovarian function. Cell Tissue Res. 2016, 363, 7–18. [Google Scholar] [CrossRef] [PubMed]
- Imbar, T.; Eisenberg, I. Regulatory role of microRNAs in ovarian function. Fertil. Steril. 2014, 101, 1524–1530. [Google Scholar] [CrossRef]
- Deb, B.; Uddin, A.; Chakraborty, S. miRNAs and ovarian cancer: An overview. J. Cell. Physiol. 2018, 233, 3846–3854. [Google Scholar] [CrossRef]
- Yin, M.; Lü, M.; Yao, G.; Tian, H.; Lian, J.; Liu, L.; Liang, M.; Wang, Y.; Sun, F. Transactivation of microRNA-383 by steroidogenic factor-1 promotes estradiol release from mouse ovarian granulosa cells by targeting RBMS1. Mol. Endocrinol. 2012, 26, 1129–1143. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Liu, Y.; Lv, M.; Xing, Q.; Zhang, Z.; He, X.; Xu, Y.; Wei, Z.; Cao, Y. miR-323-3p regulates the steroidogenesis and cell apoptosis in polycystic ovary syndrome (PCOS) by targeting IGF-1. Gene 2019, 683, 87–100. [Google Scholar] [CrossRef]
- Zhang, C.-L.; Wang, H.; Yan, C.-Y.; Gao, X.-F.; Ling, X.-J. Deregulation of RUNX2 by miR-320a deficiency impairs steroidogenesis in cumulus granulosa cells from polycystic ovary syndrome (PCOS) patients. Biochem. Biophys. Res. Commun. 2017, 482, 1469–1476. [Google Scholar] [CrossRef]
- Zhu, L.; Jing, J.; Qin, S.; Zheng, Q.; Lu, J.; Zhu, C.; Liu, Y.; Fang, F.; Li, Y.; Ling, Y. miR-130a-3p regulates steroid hormone synthesis in goat ovarian granulosa cells by targeting the PMEPA1 gene. Theriogenology 2021, 165, 92–98. [Google Scholar] [CrossRef] [PubMed]
- Guo, L.; Xu, H.; Li, Y.; Liu, H.; Zhao, J.; Lu, W.; Wang, J. Kisspeptin-10 Promotes Progesterone Synthesis in Bovine Ovarian Granulosa Cells via Downregulation of microRNA-1246. Genes 2022, 13, 298. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Zhao, J.; Xu, Q.; Guo, Y.; Liu, M.; Zhang, C.; Schinckel, A.P.; Zhou, B. MiR-31 targets HSD17B14 and FSHR, and miR-20b targets HSD17B14 to affect apoptosis and steroid hormone metabolism of porcine ovarian granulosa cells. Theriogenology 2022, 180, 94–102. [Google Scholar] [CrossRef]
- Shi, S.; Zhou, X.; Li, J.; Zhang, L.; Hu, Y.; Li, Y.; Yang, G.; Chu, G. MiR-214-3p promotes proliferation and inhibits estradiol synthesis in porcine granulosa cells. J. Anim. Sci. Biotechnol. 2020, 11, 94. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Chen, Y.; Zhou, Z.; He, X.; Tao, L.; Jiang, Y.; Lan, R.; Hong, Q.; Chu, M. chi-miR-324-3p Regulates Goat Granulosa Cell Proliferation by Targeting DENND1A. Front. Vet. Sci. 2021, 8, 732440. [Google Scholar] [CrossRef] [PubMed]
- Yao, L.; Li, M.; Hu, J.; Wang, W.; Gao, M. MiRNA-335-5p negatively regulates granulosa cell proliferation via SGK3 in PCOS. Reproduction 2018, 156, 439–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, L.; Yang, C.; Wu, H.; Chen, Q.; Huang, L.; Li, X.; Tang, H.; Jiang, Y. miR-26a-5p regulates TNRC6A expression and facilitates theca cell proliferation in chicken ovarian follicles. DNA Cell Biol. 2017, 36, 922–929. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Hou, L.; Sun, Y.; Xing, J.; Jiang, Y.; Kang, L. Single nucleotide polymorphism rs737028527 (G > A) affect miR-1b-3p biogenesis and effects on chicken egg-laying traits. Anim. Reprod. Sci. 2020, 218, 106476. [Google Scholar] [CrossRef] [PubMed]
- Wei, Q.; Li, J.; He, H.; Cao, Y.; Li, D.; Amevor, F.K.; Zhang, Y.; Wang, J.; Yu, C.; Yang, C.; et al. miR-23b-3p inhibits chicken granulosa cell proliferation and steroid hormone synthesis via targeting GDF9. Theriogenology 2022, 177, 84–93. [Google Scholar] [CrossRef]
- Li, J.; Li, C.; Li, Q.; Li, W.-T.; Li, H.; Li, G.-X.; Kang, X.; Liu, X.; Tian, Y. Identification of the Key microRNAs and miRNA-mRNA Interaction Networks during the Ovarian Development of Hens. Animals 2020, 10, 1680. [Google Scholar] [CrossRef] [PubMed]
- Bell, J.L.; Wächter, K.; Mühleck, B.; Pazaitis, N.; Köhn, M.; Lederer, M.; Hüttelmaier, S. Insulin-like growth factor 2 mRNA-binding proteins (IGF2BPs): Post-transcriptional drivers of cancer progression? Cell. Mol. Life Sci. Cmls 2013, 70, 2657–2675. [Google Scholar] [CrossRef] [PubMed]
- Nielsen, F.C.; Nielsen, J.; Christiansen, J. A family of IGF-II mRNA binding proteins (IMP) involved in RNA trafficking. Scand. J. Clin. Lab. Investig. 2001, 61, 93–99. [Google Scholar] [CrossRef]
- Taniuchi, K.; Furihata, M.; Hanazaki, K.; Saito, M.; Saibara, T. IGF2BP3-mediated translation in cell protrusions promotes cell invasiveness and metastasis of pancreatic cancer. Oncotarget 2014, 5, 6832–6845. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mancarella, C.; Scotlandi, K. IGF2BP3 From Physiology to Cancer: Novel Discoveries, Unsolved Issues, and Future Perspectives. Front. Cell Dev. Biol. 2019, 7, 363. [Google Scholar] [CrossRef] [PubMed]
- Xu, W.; Sheng, Y.; Guo, Y.; Huang, Z.; Huang, Y.; Wen, D.; Liu, C.; Cui, L.; Yang, Y.; Du, P. Increased IGF2BP3 expression promotes the aggressive phenotypes of colorectal cancer cells in vitro and vivo. J. Cell. Physiol. 2019, 234, 18466–18479. [Google Scholar] [CrossRef] [PubMed]
- Lovell, T.; Gladwell, R.; Groome, N.; Knight, P. Ovarian follicle development in the laying hen is accompanied by divergent changes in inhibin, A.; inhibin, B.; activin A and follistatin production in granulosa and theca layers. J. Endocrinol. 2003, 177, 45–56. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Ma, X.-J.; Wu, X.; Si, S.-J.; Li, C.; Yang, P.-K.; Li, G.; Liu, X.; Tian, Y.; Kang, X. Adiponectin modulates steroid hormone secretion, granulosa cell proliferation and apoptosis via binding its receptors during hens’ high laying period. Poult. Sci. 2021, 100, 101197. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2− ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Himly, M.; Foster, D.N.; Bottoli, I.; Iacovoni, J.S.; Vogt, P.K. The DF-1 chicken fibroblast cell line: Transformation induced by diverse oncogenes and cell death resulting from infection by avian leukosis viruses. Virology 1998, 248, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Zhang, B.; Chen, L.; Feng, G.; Xiang, W.; Zhang, K.; Chu, M.; Wang, P. MicroRNA mediating networks in granulosa cells associated with ovarian follicular development. BioMed Res. Int. 2017, 2017, 4585213. [Google Scholar] [CrossRef] [Green Version]
- Jiang, J.; Yang, X.; He, X.; Ma, W.; Wang, J.; Zhou, Q.; Li, M.; Yu, S. MicroRNA-449b-5p suppresses the growth and invasion of breast cancer cells via inhibiting CREPT-mediated Wnt/β-catenin signaling. Chem.-Biol. Interact. 2019, 302, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Dong, X.; Tao, S.J.; Liu, X.L.; Li, Z.; Liu, J.M.; Chen, Y. MDM4 is targeted by miR-449b-5p to promote the proliferation of endometrial carcinoma. Eur. Rev. Med. Pharmacol. Sci. 2020, 24, 11528–11535. [Google Scholar] [PubMed]
- Cheng, L.; Shi, X.; Huo, D.; Zhao, Y.; Zhang, H. MiR-449b-5p regulates cell proliferation, migration and radioresistance in cervical cancer by interacting with the transcription suppressor FOXP1. Eur. J. Pharmacol. 2019, 856, 172399. [Google Scholar] [CrossRef] [PubMed]
- Haneke, K.; Schott, J.; Lindner, D.; Hollensen, A.K.; Damgaard, C.K.; Mongis, C.; Knop, M.; Palm, W.; Ruggieri, A.; Stoecklin, G. CDK1 couples proliferation with protein synthesis. J. Cell Biol. 2020, 219, e201906147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, I.; Dynlacht, B.D. New insights into cyclins, CDKs, and cell cycle control. In Seminars in Cell & Developmental Biology; Elsevier: Amsterdam, The Netherlands, 2005; pp. 311–321. [Google Scholar]
- Wen, X.; Li, D.; Tozer, A.J.; Docherty, S.M.; Iles, R.K. Estradiol, progesterone, testosterone profiles in human follicular fluid and cultured granulosa cells from luteinized pre-ovulatory follicles. Reprod. Biol. Endocrinol. 2010, 8, 117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hillier, S.G.; Whitelaw, P.F.; Smyth, C.D. Follicular oestrogen synthesis: The ‘two-cell, two-gonadotrophin’model revisited. Mol. Cell. Endocrinol. 1994, 100, 51–54. [Google Scholar] [CrossRef]
- Hu, M.C.; Hsu, H.J.; Guo, I.C.; Chung, B.C. Function of Cyp11a1 in animal models. Mol. Cell. Endocrinol. 2004, 215, 95–100. [Google Scholar] [CrossRef] [PubMed]
- Sechman, A.; Pawlowska, K.; Hrabia, A. Effect of 3, 3′, 5-triiodothyronine and 3, 5-diiodothyronine on progesterone production, cAMP synthesis, and mRNA expression of STAR, CYP11A1, and HSD3B genes in granulosa layer of chicken preovulatory follicles. Domest. Anim. Endocrinol. 2011, 41, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Huang, E.S.-R.; Nalbandov, A. Steroidogenesis of chicken granulosa and theca cells: In vitro incubation system. Biol. Reprod. 1979, 20, 442–453. [Google Scholar] [CrossRef] [Green Version]
- Johnson, A. Steroidogenesis and actions of steroids in the hen ovary. Crit. Rev. Poult. Biol. 1990, 2, 319–346. [Google Scholar]
- Marrone, B.L.; Hertelendy, F. Steroid metabolism by avian ovarian cells during follicular maturation. Biol. Reprod. 1983, 29, 953–962. [Google Scholar] [CrossRef] [Green Version]
- Treinen, K.A.; Dodson, W.C.; Heindel, J.J. Inhibition of FSH-stimulated cAMP accumulation and progesterone production by mono (2-ethylhexyl) phthalate in rat granulosa cell cultures. Toxicol. Appl. Pharmacol. 1990, 106, 334–340. [Google Scholar] [CrossRef]
- Lee, H.T.; Bahr, J.M. Inhibition of the activities of P450 cholesterol side-chain cleavage and 3β-hydroxysteroid dehydrogenase and the amount of P450 cholesterol side-chain cleavage by testosterone and estradiol-17β in hen granulosa cells. Endocrinology 1990, 126, 779–786. [Google Scholar] [CrossRef]
- Nielsen, J.; Christiansen, J.; Lykke-Andersen, J.; Johnsen, A.H.; Wewer, U.M.; Nielsen, F.C. A family of insulin-like growth factor II mRNA-binding proteins represses translation in late development. Mol. Cell. Biol. 1999, 19, 1262–1270. [Google Scholar] [CrossRef] [Green Version]
- Giudice, L.C. Insulin-like growth factor family in Graafian follicle development and function. J. Soc. Gynecol. Investig. 2001, 8, S26–S29. [Google Scholar]
- Spicer, L.; Aad, P. Insulin-like growth factor (IGF) 2 stimulates steroidogenesis and mitosis of bovine granulosa cells through the IGF1 receptor: Role of follicle-stimulating hormone and IGF2 receptor. Biol. Reprod. 2007, 77, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Hsu, K.; Shen, M.; Huang, Y.; Cheng, Y.; Lin, S.; Chow, N.; Cheng, S.; Chou, C.; Ho, C. Overexpression of the RNA-binding proteins Lin28B and IGF2BP3 (IMP3) is associated with chemoresistance and poor disease outcome in ovarian cancer. Br. J. Cancer 2015, 113, 414–424. [Google Scholar] [CrossRef]
- Ren, F.; Lin, Q.; Gong, G.; Du, X.; Dan, H.; Qin, W.; Cheng, S.-W.; Chou, C.-Y.; Ho, C.-L. Igf2bp3 maintains maternal RNA stability and ensures early embryo development in zebrafish. Commun. Biol. 2020, 3, 414–424. [Google Scholar] [CrossRef] [Green Version]
- Wu, X.; Zhang, Y.; Xu, S.; Chang, Y.; Ye, Y.; Guo, A.; Kang, Y.; Guo, H.; Xu, H.; Chen, L.; et al. Loss of Gsdf leads to a dysregulation of Igf2bp3-mediated oocyte development in medaka. Gen. Comp. Endocrinol. 2019, 277, 122–129. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primer Name | Primer Sequence (5′–3′) | Product Length (bp) | Genbank Number |
|---|---|---|---|
| CCND1 | F: ATAGTCGCCACTTGGATGCT | 122 | NM_205381 |
| R: AACCGGCTTTTCTTGAGGGG | |||
| CCND2 | F: TCCGGAAACATGCACAAACG | 257 | XM_015292118.2 |
| R: CCGGACTTGCCTAAGGTTGC | |||
| CDK1 | F: TGGCCTTGAACCACCCATAC | 147 | NM_205314.1 |
| R: AGGCAGGCAGGCAAAGATAA | |||
| CDK2 | F: ACGTGATCCACACGGAGAAC | 132 | NM_001199857 |
| R: GCAGCTGGAACAGGTAGCTC | |||
| CDK6 | F: AGCAGCCCAGAAGAGATGATT | 132 | NM_001007892.2 |
| R: GAGAAATACGCACAAACCCTGT | |||
| StAR | F: GTCCCTCGCAGACCAAGT | 196 | NM_204686 |
| R: TCCCTACTGTTAGCCCTGA | |||
| CYP11A1 | F: GTGGACACGACTTCCATGACT | 174 | NM_001001756 |
| R: GAGAGTCTCCTTGATGGCGG | |||
| 3β-HSD | F: TGGAAGAAGATGAGGCGCTG | 185 | NM_205118 |
| R: GGAAGCTGTGTGGATGACGA | |||
| CYP19A1 | F: GGCCTCCAGCAGGTTGAAAG | 214 | NM_001001761.3 |
| R: ATAGGCACTGTGGCAACTGG | |||
| FSHR | F: GAGCGAGGTCTACATACA | 281 | NM_205079 |
| R: GCACAAGCCATAGTCA | |||
| LHR | F: GGGCTTTCCCAAGCCTACAT | 133 | NM_204936.2 |
| R: TGGTGTCTTTATTGGCGGCT | |||
| IGFBP4 | F: AACTTCCACCCCAAGCAGT | 123 | NM_204353.1 |
| R: GCAATCCAAGTCCCCCTTCA | |||
| PGRMC1 | F: AGATCGTGGGCTCACCTCTA | 157 | NM_001271939.1 |
| R: AGCTGCTCCAGTGTGAAGTC | |||
| MMP2 | F: CGATGCTGTCTACGAGTCCC | 96 | NM_204420.2 |
| R: TAGCCCCTATCCAGGTTGCT | |||
| IGF2BP3 | F: TCCTGGTGAAGACGGGCTAC | 133 | XM_015281444.4 |
| R: CTTTTAGGGACCGAATGCTC | |||
| BMP3 | F: ACAGGGCAAAGAGTAAGAAAAAG | 136 | NM_001034819.2 |
| R: AGATAGCGTCGGGCACAATA | |||
| E2F5 | F: GCCTTCCAGACTCAGTGTTG | 148 | NM_001030942.1 |
| R: GGCTCCTCCATCTTTGCTAT | |||
| β-actin | F: CAGCCAGCCATGGATGATGA | 147 | NM_205518.2 |
| R: ACCAACCATCACACCCTGAT | |||
| IGF2BP3 WT | F: ccgctcgagTTACATAACACTGCCATGAATA | 244 | - |
| R: ataagaatgcggccgcAGTCCGTAGTACTCCTGGCTGG | |||
| IGF2BP3 MUT | F: ccgctcgagTTACATAATGACATAGTGAATAACCTAAGGGA | 244 | - |
| R: ataagaatgcggccgcAGTCCGTAGTACTCCTGGCTGG | |||
| MMP2 WT | F: ccgctcgagCGAGTTTGATCATTACTGCCA | 337 | - |
| R: ataagaatgcggccgcGAAAGCCTAACCAAACAAAAC | |||
| MMP2 MUT | F: ccgctcgagCGAGTTTGATCATTGACATTGTTTATTTACATAAT | 337 | - |
| R: ataagaatgcggccgcGAAAGCCTAACCAAACAAAAC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, X.; Zhang, N.; Li, J.; Zhang, Z.; Guo, Y.; Li, D.; Zhang, Y.; Gong, Y.; Jiang, R.; Li, H.; et al. gga-miR-449b-5p Regulates Steroid Hormone Synthesis in Laying Hen Ovarian Granulosa Cells by Targeting the IGF2BP3 Gene. Animals 2022, 12, 2710. https://doi.org/10.3390/ani12192710
Wu X, Zhang N, Li J, Zhang Z, Guo Y, Li D, Zhang Y, Gong Y, Jiang R, Li H, et al. gga-miR-449b-5p Regulates Steroid Hormone Synthesis in Laying Hen Ovarian Granulosa Cells by Targeting the IGF2BP3 Gene. Animals. 2022; 12(19):2710. https://doi.org/10.3390/ani12192710
Chicago/Turabian StyleWu, Xing, Na Zhang, Jing Li, Zihao Zhang, Yulong Guo, Donghua Li, Yanhua Zhang, Yujie Gong, Ruirui Jiang, Hong Li, and et al. 2022. "gga-miR-449b-5p Regulates Steroid Hormone Synthesis in Laying Hen Ovarian Granulosa Cells by Targeting the IGF2BP3 Gene" Animals 12, no. 19: 2710. https://doi.org/10.3390/ani12192710
APA StyleWu, X., Zhang, N., Li, J., Zhang, Z., Guo, Y., Li, D., Zhang, Y., Gong, Y., Jiang, R., Li, H., Li, G., Liu, X., Kang, X., & Tian, Y. (2022). gga-miR-449b-5p Regulates Steroid Hormone Synthesis in Laying Hen Ovarian Granulosa Cells by Targeting the IGF2BP3 Gene. Animals, 12(19), 2710. https://doi.org/10.3390/ani12192710