
The Influence of Forestry Management on the Selection of a Non-Vegetative Diet by the Eurasian Beaver (Castor fiber L.)
 , ,
, , 
Abstract
:Simple Summary
Abstract
1. Introduction
- (a)
- To analyze on the individual woody plant level if there are preferences for some taxa of woody plants, especially for commercial species, i.e., Quercus and Fraxinus;
- (b)
- To analyze on the individual level, what is the role of distance from water and diameter of the woody plant on the probability of the felling by beaver and if these effects are taxa-specific;
- (c)
- To analyze on the patch level if some patch characteristics, primarily the type of forest stand (commercial monoculture, commercial mixed, natural), affect the proportion of felled basal area, the proportion of used woody plant taxa from their diversity in the stand, and the maximal distance of felled woody plants.
2. Materials and Methods
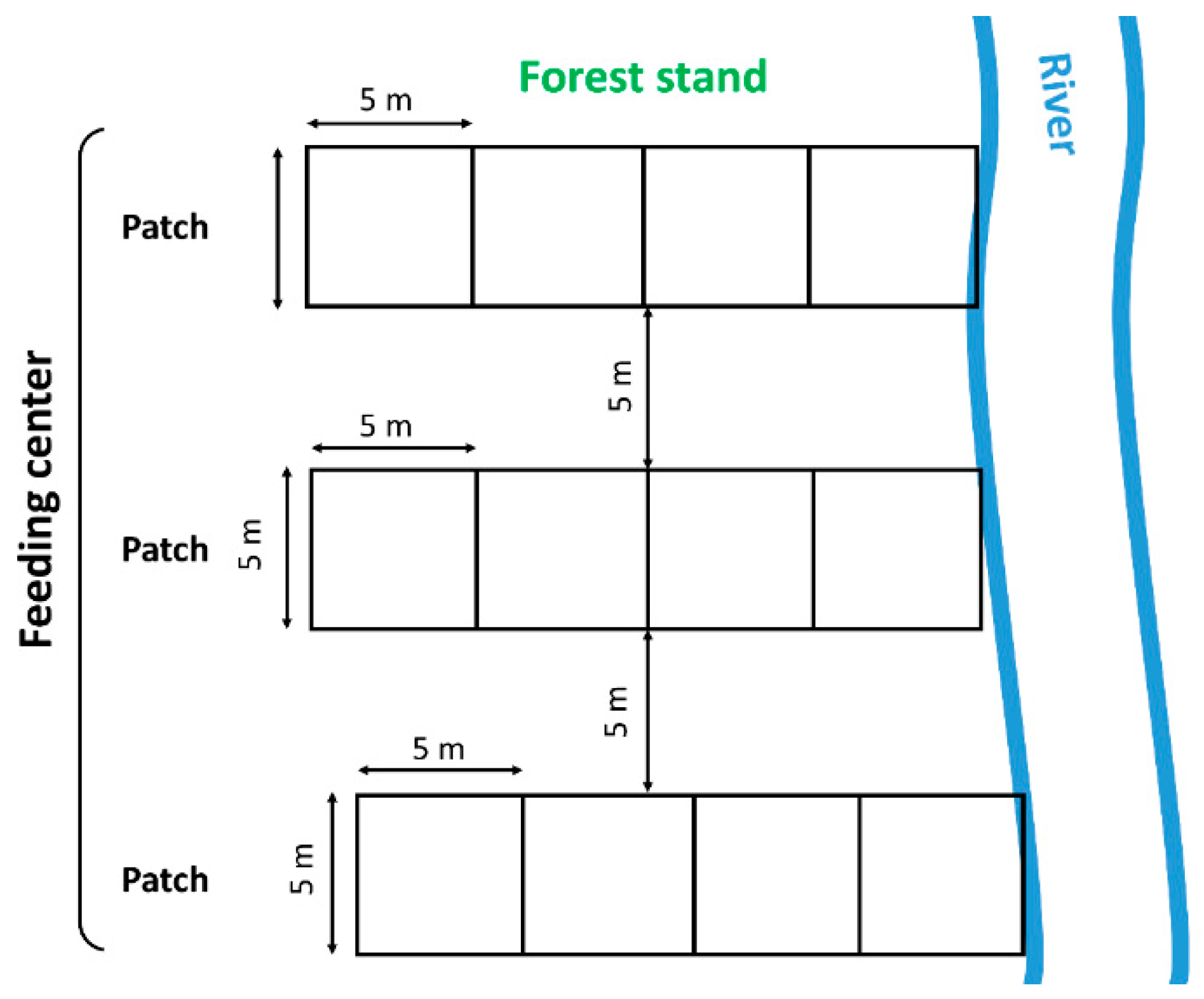
2.1. Study Sites
2.2. Data Collection
2.3. Data Processing and Evaluation
3. Results
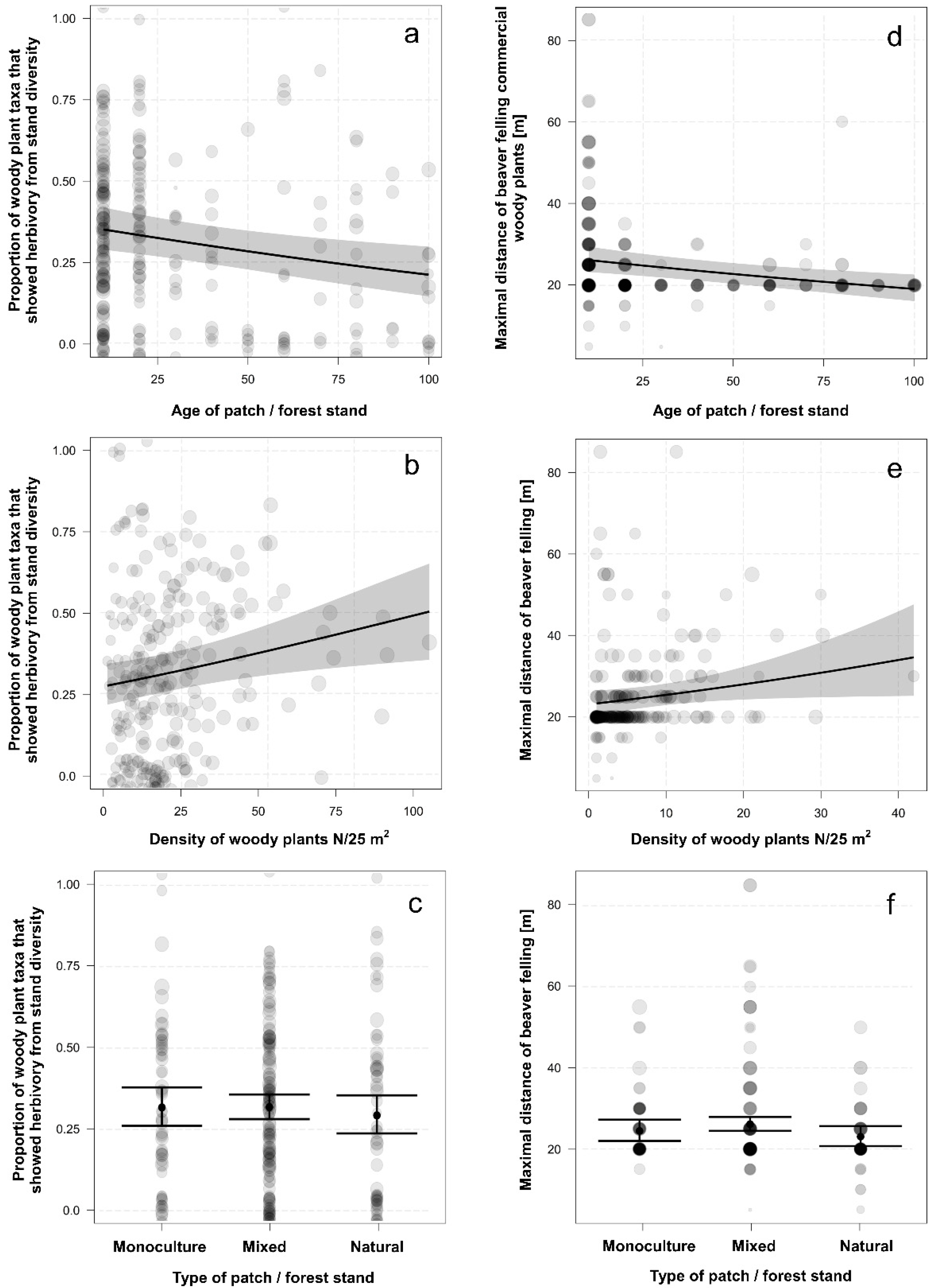
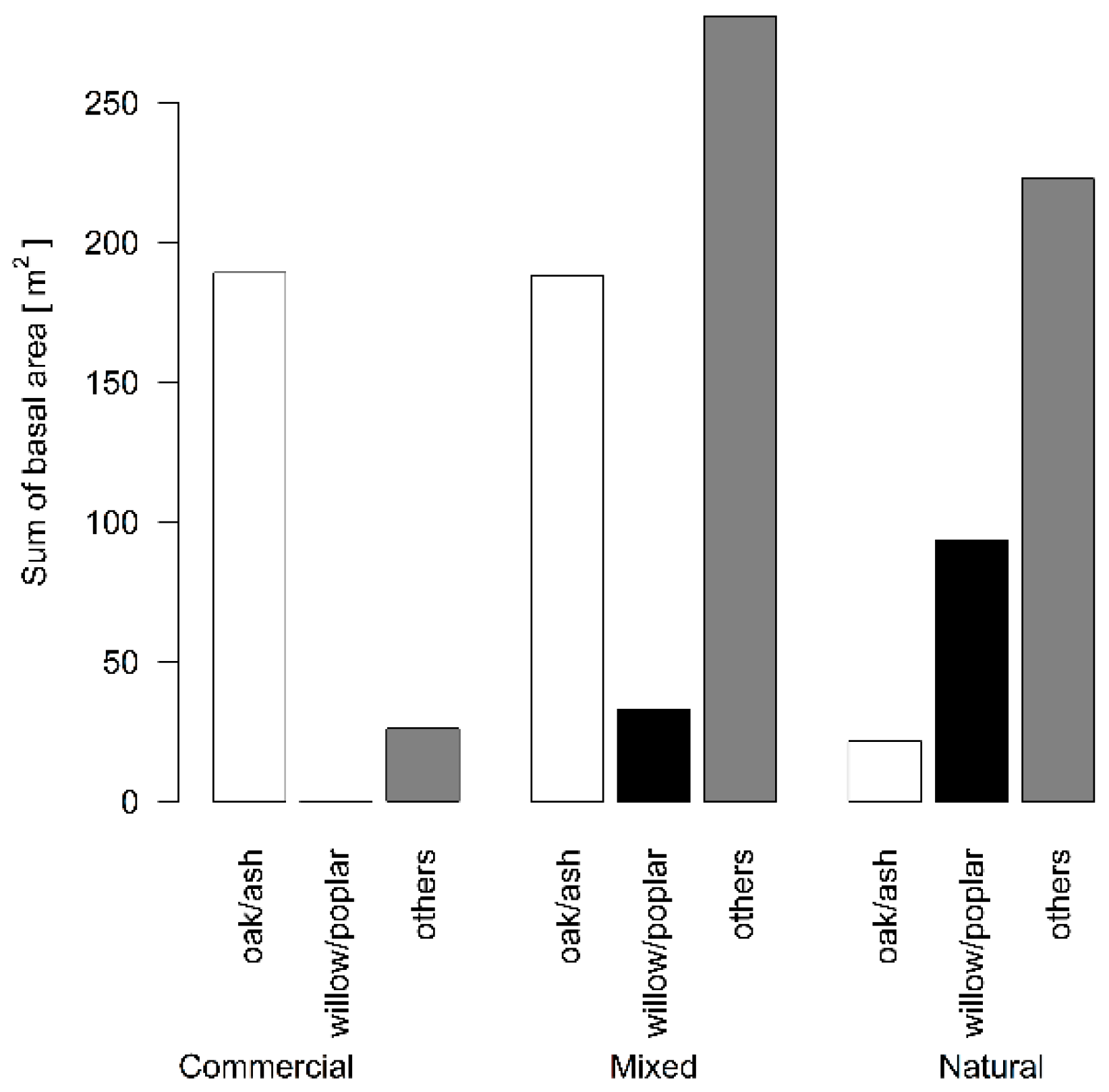
3.1. Differences in Proportion of Felled Woody Plants
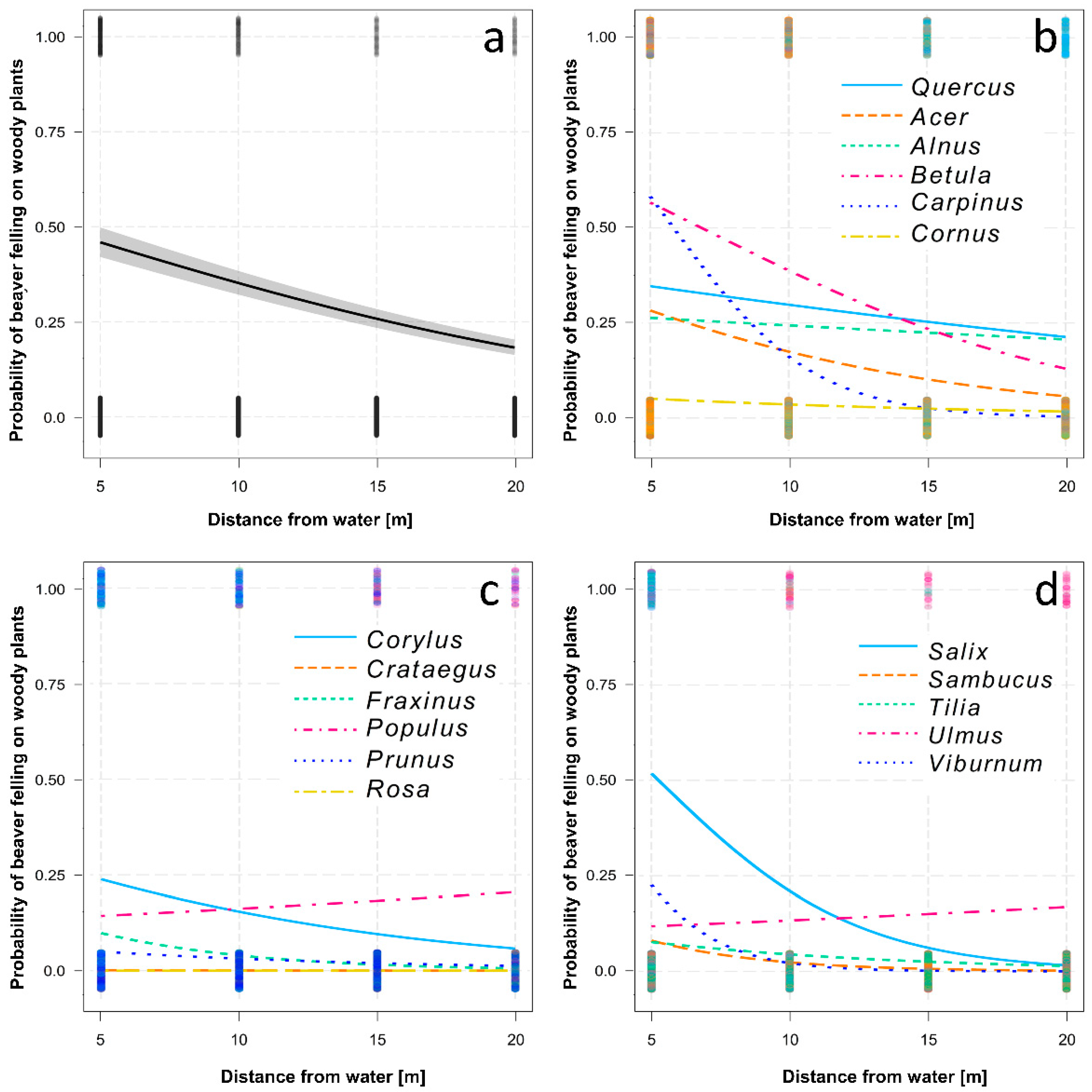
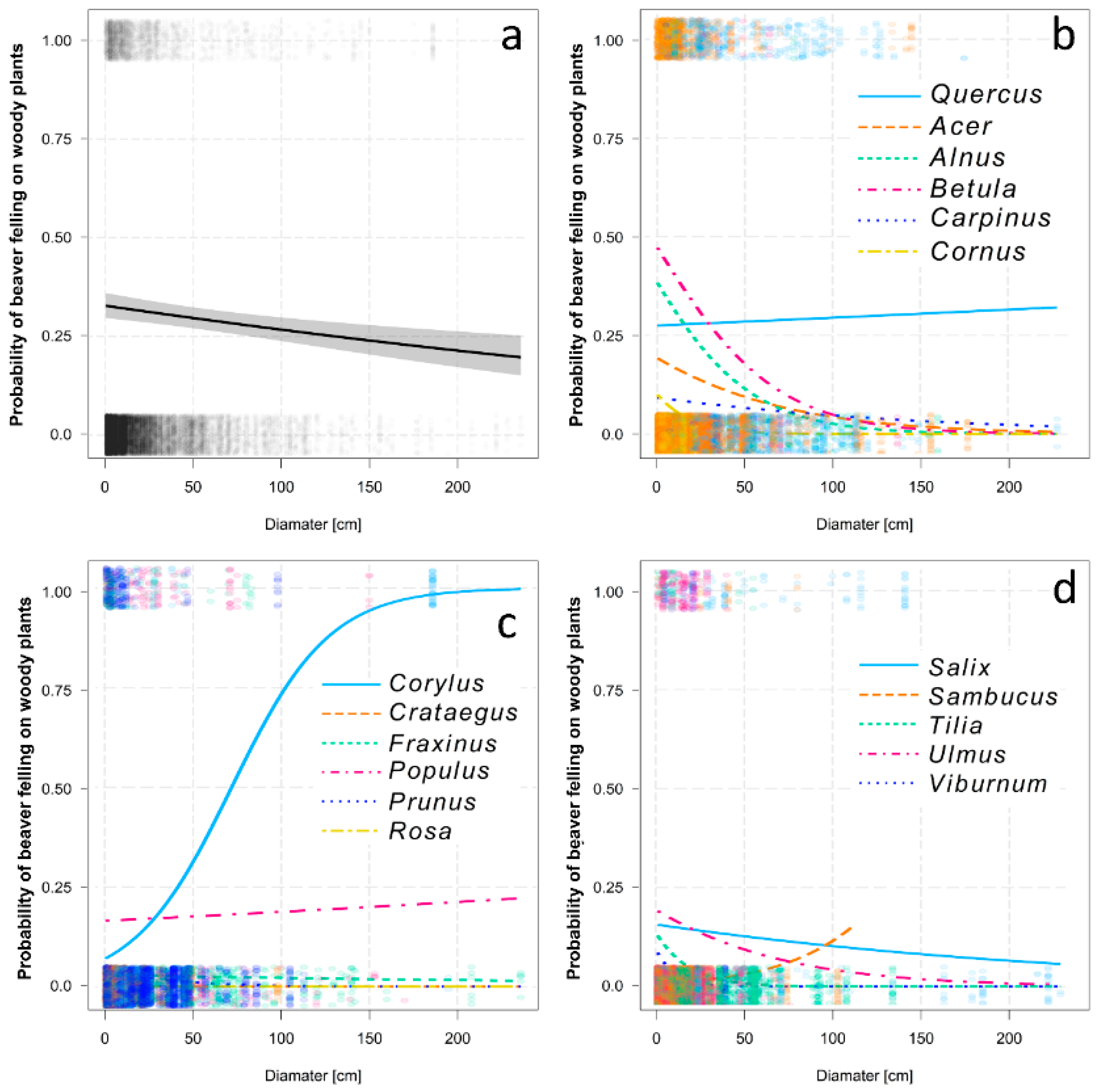
3.2. Woody Plant Diameter and Distance of Felled Woody Plants from the Water
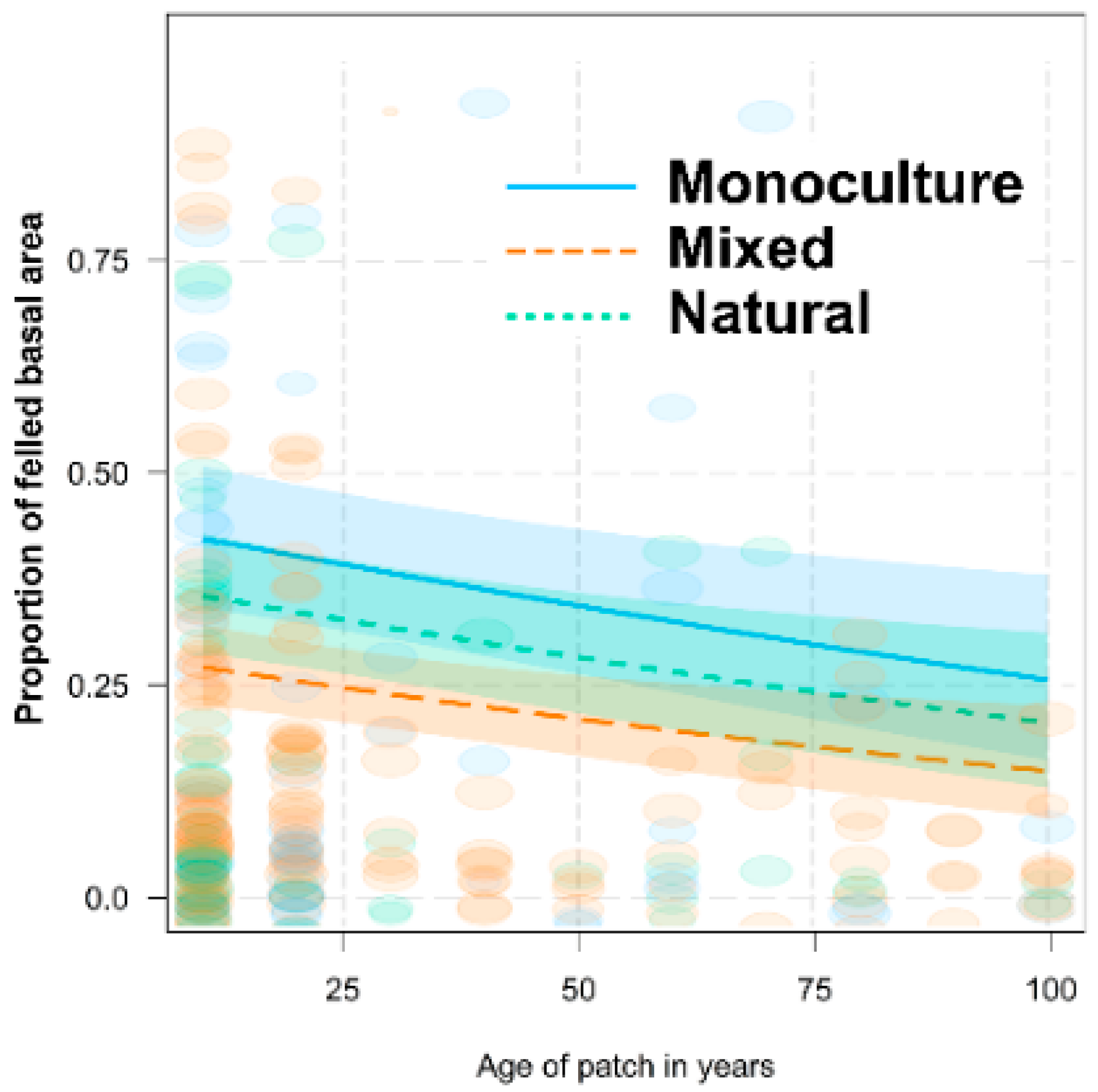
3.3. Differences among Forest Types in Foraging Behavior
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

References
- Nolet, B.A.; Rosell, F. Comeback of the beaver Castor fiber: An overview of old and new conservation problems. Biol. Conserv. 1998, 83, 165–173. [Google Scholar] [CrossRef] [Green Version]
- Halley, D.J.; Saveljev, A.P.; Rosell, F. Population and distribution of beavers Castor fiber and Castor canadensis in Eurasia. Mammal Rev. 2021, 51, 1–24. [Google Scholar] [CrossRef]
- Wróbel, M. Population of Eurasian beaver (Castor fiber) in Europe. Glob. Ecol. 2020, 23, e01046. [Google Scholar] [CrossRef]
- AOPK, CR—Finding Database of Nature Protection. Occurrence and Distribution of Species of Animals, Plants and Fungi in the Czech Republic. Available online: http://portal.nature.cz (accessed on 20 November 2020).
- Fitzgerald, W.S.; Thompson, R.A. Problems associated with beaver in stream or floodway management. Vertebr. Pest Conf. 1988, 13, 190–195. [Google Scholar]
- Mikulka, O.; Homolka, M.; Drimaj, J.; Kamler, J. European beaver (Castor fiber) in open agricultural landscapes: Crop grazing and the potential for economic damage. Eur. J. Wildl. Res. 2020, 66, 101. [Google Scholar] [CrossRef]
- Harkonen, S. Forest damage caused by the Canadian beaver (Castor canadensis) in South Savo. Finland. Silva Fenn. 1999, 33, 247–259. [Google Scholar] [CrossRef] [Green Version]
- Gorczyca, E.; Krzemień, K.; Sobucki, M.; Jarzyna, K. Can beaver impact promote river renaturalization? The example of the Raba River, southern Poland. Sci. Total Environ. 2018, 615, 1048–1060. [Google Scholar] [CrossRef]
- Osipov, V.V.; Bashinskiy, I.V.; Podshivalina, V.N. Influence of the activity of the Eurasian beaver Castor fiber (Castoridae, Mammalia) on the ecosystem biodiversity of small Rivers in the Forest-steppe zone. Biol. Bull 2018, 45, 1139–1148. [Google Scholar] [CrossRef]
- Stringer, A.P.; Gaywood, M.J. The impacts of beavers Castor spp. on biodiversity and the ecological basis for their reintroduction to Scotland, UK. Mammal Rev. 2016, 46, 270–283. [Google Scholar] [CrossRef]
- Wilsson, L. Observations and Experiments on the Ethology of the European Beaver (Castor fiber L.): A study in the Development of Phylogenetically Adapted Behaviour in a Highly Specialized Mammal. Dissertation Thesis, Stockholm University, Stockholm, Sweden, 1971. [Google Scholar]
- Müller–Schwarze, D.; Sun, L. The Beaver—Natural History of a Wetlands Engineer; Cornell University Press: Ithaca, NY, USA, 2003; 190p. [Google Scholar]
- Jenkins, S.H. Seasonal and year-to-year differences in food selection by beavers. Oecologia 1979, 44, 112–116. [Google Scholar] [CrossRef]
- Busher, P.E.; Mayer, M.; Ulevičius, A.; Samus, A.; Hartman, G.; Rosell, F. Food caching behavior of the Eurasian beaver in northern Europe. Wildl. Biol. 2020, 2020, 1–10. [Google Scholar] [CrossRef]
- Jackowiak, M.; Busher, P.; Krauze-Gryz, D. Eurasian Beaver (Castor fiber) Winter Foraging Preferences in Northern Poland—The Role of Woody Vegetation Composition and Anthropopression Level. Animals 2020, 10, 1376. [Google Scholar] [CrossRef] [PubMed]
- Busher, P.E. Food caching behavior of beavers (Castor canadensis): Selection and use of woody species. Am. Midl. Nat. 1996, 135, 343–348. [Google Scholar] [CrossRef]
- Haarberg, O.; Rosell, F. Selective foraging on woody plant species by the Eurasian beaver (Castor fiber) in Telemark, Norway. J. Zool. 2006, 270, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Nolet, B.A.; Hoekstra, A.; Ottenheim, M.M. Selective foraging on woody species by the beaver Castor fiber and its impact on a riparian willow forest. Biol. Con. 1994, 70, 117–128. [Google Scholar] [CrossRef]
- Novak, M.; Baker, J.A.; Obbard, M.E.; Malloch, B. Wild Furbearer Management and Conservation in North America; Ontario Ministry of Natural Resources: Toronto, ON, Canada, 1987. [Google Scholar]
- Parker, H.; Haugen, A.; Kristensen, Ø.; Myrum, E.; Kolsing, R.; Rosell, F. Landscape Use and Economic Value of Eurasian Beaver (Castor fiber) on Large Forest in Southeast Norway; European-American Beaver Congress: Kazan, Russia, 1999. [Google Scholar]
- Jenkins, S.H. A size-distance relation in food selection by beavers. Ecology 1980, 61, 740–746. [Google Scholar] [CrossRef] [Green Version]
- Jenkins, S.H. Food selection by beavers. Oecologia 1975, 21, 157–173. [Google Scholar] [CrossRef]
- Aleksiuk, M. The seasonal food regime of arctic beavers. Ecology 1970, 51, 264–270. [Google Scholar] [CrossRef]
- Vorel, A.; Válková, L.; Hamšíková, L.; Maloň, J.; Korbelová, J. Beaver foraging behaviour: Seasonal foraging specialization by a choosy generalist herbivore. Behav. Ecol. 2015, 69, 1221–1235. [Google Scholar] [CrossRef]
- Urban, J.; Suchomel, J.; Dvořák, J. Contribution to the knowledge of woods preferences of European beaver (Castor fiber L. 1758) in bank vegetation on non-forest land in the forest district Soutok (Czech Republic). Acta Univ. Agric. Silvic. 2014, 56, 289–294. [Google Scholar] [CrossRef] [Green Version]
- Dvořák, J. Diet preference of Eurasian Beaver (Castor fiber L. 1758) in the environment of Oderské vrchy and its influence on the tree species composition of river bank stands. Acta Univ. Agric. Silvic. 2013, 61, 1637–1643. [Google Scholar] [CrossRef]
- John, F.; Baker, S.; Kostkan, V. Habitat selection of an expanding beaver (Castor fiber) population in central and upper Morava River basin. Eur. J. Wildl. Res. 2010, 56, 663–671. [Google Scholar] [CrossRef] [Green Version]
- Janiszewski, P.; Hermanowska, Z. Damage caused by the European beaver (Castor fiber L.) in agricultural and forest farms in view of selected atmospheric factors and animal behavior. Appl. Ecol. Environ. Res. 2019, 17, 15633–15642. [Google Scholar] [CrossRef]
- Fustec, J.; Lodé, T.; Le Jacques, D.; Cormier, J.P. Colonization. riparian habitat selection and home range size in a reintroduced population of European beavers in the Loire. Freshw. Biol. 2001, 46, 1361–1371. [Google Scholar] [CrossRef]
- Margaletić, J.; Grubešić, M.; Dušak, V.; Konjević, D. Activity of European beavers (Castor fiber L.) in young pedunculate oak (Quercus robur L.) forests. Vet. Arch. 2006, 76, 167–175. [Google Scholar]
- Schley, L. Characteristics of trees and shrubs felled by a Eurasian beaver. Bull Soc. Nat. Luxemb. 2004, 105, 133–136. [Google Scholar]
- Kamczyc, J.; Bielachowicz, M.; Pers–Kamczyc, E. Damages caused by European beaver (Castor fiber L. 1758) in broadleaved stands. For. Lett. 2016, 109, 7–10. [Google Scholar]
- Vorel, A.; Šafář, J.; Šimůnková, K. Recentní rozšíření bobra evropského (Castor fiber) v České republice v letech 2002–2012 (Rodentia: Castoridae). Lynx 2012, 43, 169–179. (in Czech). [Google Scholar]
- Quantum, G.I.S. Development Team, 2014. In QGIS Geographic Information System; Open Source Geospatial Foundation Project: Chicago, IL, USA, 2019. [Google Scholar]
- Lesprojekt. Forest Management Plan Židlochovice (1 January 2010––31 December 2019); Lesprojekt: Brno, Czech Republic, 2010. (in Czech) [Google Scholar]
- Taxonia. Forest Management Plan Pomoraví (1 January 2010–31 December 2019); Lesprojekt: Olomouc, Czech Republic; Taxonia: Olomouc, Czech Republic, 2010. (in Czech) [Google Scholar]
- R Core Team, R. A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2022. [Google Scholar]
- Venables, W.N.; Ripley, B.D. Modern Applied Statistics with S, 4th ed.; Springer: New York, NY, USA, 2002. [Google Scholar]
- Brooks, M.E.; Kristensen, K.; van Benthem, K.J.; Magnusson, A.; Berg, C.W.; Nielsen, A.; Skaug, H.J.; Maechler, M.; Bolker, B.M. glmmTMB Balances Speed and Flexibility among Packages for Zero-I Flated Generalized Linear Mixed Modeling. R J. 2017, 9, 378–400. [Google Scholar] [CrossRef] [Green Version]
- Bates, D.; Maechler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- Fox, J.; Weisberg, S. An {R} Companion to Applied Regression, Third Edition. Thousand Oaks CA: Sage. 2019. Available online: https://socialsciences.mcmaster.ca/jfox/Books/Companion/ (accessed on 27 September 2018).
- Long, J.A. Jtools: Analysis and Presentation of Social Scientific Data_. R Package Version 2.2.0. 2022. Available online: https://cran.r-project.org/package=jtools (accessed on 25 April 2022).
- Long, J.A. Interactions: Comprehensive, User-Friendly Toolkit for Probing Interactions. R package Version 1.1.0. 2019. Available online: https://cran.r-project.org/package=interactions (accessed on 2 July 2021).
- Danilov, P.I.; Kan’shiev, V.Y. State of populations and ecological characteristics of European (Castor fiber L.) and Canadian (Castor canadensis Kuhl.) beavers in the northwestern USSR. Acta Zool. Fenn. 1983, 174, 95–97. [Google Scholar]
- Doucet, C.M.; Fryxell, J.M. The effect of nutritional quality on forage preference by beavers. Oikos 1993, 67, 201–208. [Google Scholar] [CrossRef]
- Klich, D. Selective bark stripping of various tree species by Polish horses in relation to bark detachability. For. Ecol. Manag. 2017, 384, 65–71. [Google Scholar] [CrossRef]
- Basey, J.M.; Jenkins, S.H. Influences off predation risk and energy maximization on food selection by beavers (Castor canadensis). Can. J. Zool. 1995, 73, 2197–2208. [Google Scholar] [CrossRef]
- Boege, K.; Marquis, R.J. Facing herbivory as you grow up: The ontogeny of resistance in plants. Trends Ecol. Evol. 2005, 20, 441–448. [Google Scholar] [CrossRef]
- Basey, J.M.; Jenkins, S.H.; Miller, G.C. Food selection by beavers in relation to inducible defenses of Populus tremuloides. Oikos 1990, 59, 57–62. [Google Scholar] [CrossRef]
- Basey, J.M.; Jenkins, S.H.; Busher, P.E. Optimal central-place foraging by beavers: Tree-size selection in relation to defensive chemicals of quaking aspen. Oecologia 1988, 76, 278–282. [Google Scholar] [CrossRef]
- Rosell, F.; Czech, A. Responses of foraging Eurasian beavers Castor fiber to predator odours. Wildl. Biol. 2000, 6, 13–21. [Google Scholar] [CrossRef] [Green Version]
- Severud, W.J.; Belant, J.L.; Bruggink, J.G.; Windels, S.K. Predator cues reduce American beaver use of foraging trails. Hum. Wildl. Interact. 2011, 5, 296–305. [Google Scholar]
- Doucet, C.M.; Adams, I.T.; Fryxell, J.M. Beaver dam and cache composition: Are woody species used differently? Ecoscience 1994, 1, 268–270. [Google Scholar] [CrossRef]
- Tajchman, K.; Czyżowski, P.; Drozd, L. Food selectivity of European beaver (Castor fiber L.) occurring in the area Lubaczow and Chotylow Forest Districts. Pol. J. Nat. Sci. 2018, 33, 533–543. [Google Scholar]
- Ważna, A.; Cichocki, J.; Bojarski, J.; Gabryś, G. Selective foraging on tree and shrub species by the European beaver Castor fiber in lowland and highland habitats in western Poland. Pol. J. Ecol. 2018, 66, 286–300. [Google Scholar] [CrossRef]
- Mahoney, M.J.; Stella, J.C. Stem size selectivity is stronger than species preferences for beaver. a central place forager. For. Ecol. Man. 2020, 475, 118331. [Google Scholar] [CrossRef]
- Fryxell, J.M.; Doucet, C.M. Provisioning time and central-place foraging in beavers. Can. J. Zool. 1991, 69, 1308–1313. [Google Scholar] [CrossRef]
- Salandre, J.A.; Beil, R.; Loehr, J.A. Sundell J Foraging decisions of North American beaver (Castor canadensis) are shaped by energy constraints and predation risk. Mamm. Res. 2017, 62, 229–239. [Google Scholar] [CrossRef]
- Baker, S.; Fraser, D.; Kostkan, V. A modified method for appraising the suitability of urban sites in Great Britain, for use by the Eurasian (European) beaver (Castor fiber). J. Practical. Ecol. Conservat. 2006, 5, 22–37. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monoculture | Mixed | Natural | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Genus Species | Specific Species | Available | Fell | Available | Fell | Available | Fell | Available | Fell |
| Quercus spp. [comm] | robur, petraea, cerris | 70.63 | 18.31 | 44.05 | 8.27 | 6.66 | 1.03 | 37.63 | 8.07 |
| Fraxinus spp. [comm] | excelsior, augustifolia | 25.38 | 2.39 | 15.57 | 1.26 | 3.10 | 0.24 | 13.63 | 1.17 |
| Populus spp. | tremula, alba, nigra | 0.01 | 0.00 | 9.87 | 3.72 | 14.28 | 2.17 | 9.14 | 2.41 |
| Tilia cordata | 0.57 | 0.00 | 5.32 | 0.38 | 5.49 | 0.03 | 4.32 | 0.18 | |
| Acer campestre | 1.28 | 0.40 | 4.52 | 0.51 | 4.67 | 1.22 | 3.85 | 0.72 | |
| Ulmus laevis | 0.10 | 0.00 | 3.88 | 0.38 | 3.99 | 0.23 | 3.08 | 0.25 | |
| Salix spp. | alba, euxina, cinerea, caprea, triandra | 0.00 | 0.00 | 3.40 | 0.93 | 23.71 | 6.58 | 9.33 | 2.57 |
| Acer pseudoplatanus | 0.00 | 0.00 | 2.34 | 0.42 | 11.91 | 3.97 | 4.97 | 1.49 | |
| Sambucus nigra | 0.15 | 0.00 | 1.88 | 0.22 | 1.17 | 0.00 | 1.26 | 0.10 | |
| Carpinus betulus | 0.01 | 0.00 | 1.79 | 0.20 | 0.93 | 0.00 | 1.12 | 0.09 | |
| Alnus glutinosa | 0.02 | 0.01 | 1.54 | 0.10 | 11.56 | 0.92 | 4.50 | 0.35 | |
| Crataegus spp. | monogyna, laevigata | 0.23 | 0.01 | 1.20 | 0.01 | 0.18 | 0.01 | 0.65 | 0.01 |
| Prunus avium | 0.26 | 0.01 | 1.09 | 0.06 | 0.47 | 0.02 | 0.70 | 0.04 | |
| Acer negundo | 0.00 | 0.00 | 0.59 | 0.06 | 2.18 | 0.26 | 0.98 | 0.11 | |
| Betula pendula | 0.00 | 0.00 | 0.56 | 0.06 | 2.12 | 0.38 | 0.95 | 0.15 | |
| Prunus padus | 0.89 | 0.44 | 0.54 | 0.00 | 2.74 | 0.25 | 1.34 | 0.17 | |
| Acer platanoides | 0.09 | 0.00 | 0.43 | 0.10 | 2.81 | 1.46 | 1.14 | 0.52 | |
| Robinia pseudoacacia | 0.00 | 0.00 | 0.41 | 0.00 | 0.00 | 0.00 | 0.18 | 0.00 | |
| Viburnum opulus | 0.00 | 0.00 | 0.30 | 0.03 | 0.11 | 0.04 | 0.17 | 0.03 | |
| Cornus spp. | sanguinea, mas | 0.25 | 0.03 | 0.27 | 0.04 | 0.09 | 0.02 | 0.21 | 0.03 |
| Pinus spp. | sylvestris, strobus | 0.00 | 0.00 | 0.24 | 0.00 | 0.16 | 0.00 | 0.16 | 0.00 |
| Corylus avellana | 0.05 | 0.00 | 0.08 | 0.00 | 1.01 | 0.59 | 0.38 | 0.19 | |
| Prunus avium | 0.00 | 0.00 | 0.07 | 0.00 | 0.00 | 0.00 | 0.03 | 0.00 | |
| Euonymus europaeus | 0.00 | 0.00 | 0.04 | 0.00 | 0.00 | 0.00 | 0.02 | 0.00 | |
| Rosa canina | 0.06 | 0.00 | 0.02 | 0.00 | 0.01 | 0.00 | 0.03 | 0.00 | |
| Pyrus pyraster | 0.00 | 0.00 | <0.01 | 0.00 | 0.00 | 0.00 | <0.00 | 0.00 | |
| Ligustrum vulgare | 0.00 | 0.00 | <0.01 | <0.01 | 0.00 | 0.00 | <0.01 | <0.01 | |
| Rubus idaeus | 0.00 | 0.00 | <0.01 | 0.00 | 0.00 | 0.00 | <0.01 | 0.00 | |
| Glossularia uva crispa | 0.00 | 0.00 | <0.01 | 0.00 | <0.01 | 0.00 | <0.01 | 0.00 | |
| Aesculus hippocastanum | 0.00 | 0.00 | 0.00 | 0.00 | 0.63 | 0.29 | 0.21 | 0.10 | |
| Fagus sylvaticus | <0.01 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | <0.01 | 0.00 | |
| Malus domestica | 0.00 | 0.00 | 0.00 | 0.00 | <0.01 | 0.00 | <0.01 | 0.00 | |
| Frangula alnus | 0.00 | 0.00 | 0.00 | 0.00 | 0.01 | 0.00 | <0.01 | 0.00 | |
| Amorpha fructicosa | 0.00 | 0.00 | 0.00 | 0.00 | <0.01 | 0.00 | <0.01 | 0.00 | |
| Total | 100.00 | 21.59 | 100.00 | 16.74 | 100.00 | 19.72 | 100.00 | 18.76 | |
| Genus | Coefficient | SE | t-Value | p-Value | Significance |
|---|---|---|---|---|---|
| Acer | −0.71 | 0.24 | −2.96 | 0.003 | ** |
| Alnus | −0.94 | 0.51 | −1.87 | 0.062 | . |
| Betula | 1.01 | 1.02 | 0.99 | 0.321 | |
| Carpinus | −1.18 | 0.91 | −2.41 | 0.016 | * |
| Cornus | −1.94 | 0.33 | −5.86 | <0.001 | *** |
| Corylus | −0.52 | 0.31 | −1.68 | 0.092 | . |
| Crataegus | −2.29 | 0.41 | −5.53 | <0.001 | *** |
| Fraxinus | −1.69 | 0.50 | −3.39 | <0.001 | *** |
| Populus | 1.24 | 0.48 | 2.60 | 0.009 | ** |
| Prunus | −1.13 | 0.28 | −3.99 | <0.001 | *** |
| Rosa | 21.41 | 7713.78 | 0.00 | 0.998 | |
| Salix | 0.45 | 0.46 | 0.99 | 0.324 | |
| Sambucus | −4.85 | 0.70 | −6.98 | <0.001 | *** |
| Tilia | −3.07 | 0.31 | −9.80 | <0.001 | *** |
| Ulmus | −1.46 | 0.52 | −2.83 | 0.005 | ** |
| Viburnum | 2.93 | 0.76 | 3.85 | <0.001 | *** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mikulka, O.; Pyszko, P.; Skoták, V.; Kamler, J.; Drimaj, J.; Plhal, R.; Homolka, M. The Influence of Forestry Management on the Selection of a Non-Vegetative Diet by the Eurasian Beaver (Castor fiber L.). Animals 2022, 12, 2949. https://doi.org/10.3390/ani12212949
Mikulka O, Pyszko P, Skoták V, Kamler J, Drimaj J, Plhal R, Homolka M. The Influence of Forestry Management on the Selection of a Non-Vegetative Diet by the Eurasian Beaver (Castor fiber L.). Animals. 2022; 12(21):2949. https://doi.org/10.3390/ani12212949
Chicago/Turabian StyleMikulka, Ondřej, Petr Pyszko, Vlastimil Skoták, Jiří Kamler, Jakub Drimaj, Radim Plhal, and Miloslav Homolka. 2022. "The Influence of Forestry Management on the Selection of a Non-Vegetative Diet by the Eurasian Beaver (Castor fiber L.)" Animals 12, no. 21: 2949. https://doi.org/10.3390/ani12212949
APA StyleMikulka, O., Pyszko, P., Skoták, V., Kamler, J., Drimaj, J., Plhal, R., & Homolka, M. (2022). The Influence of Forestry Management on the Selection of a Non-Vegetative Diet by the Eurasian Beaver (Castor fiber L.). Animals, 12(21), 2949. https://doi.org/10.3390/ani12212949