

Lysolecithin Improves Broiler Growth Performance through Upregulating Growth-Related Genes and Nutrient Transporter Genes Expression Independent of Experimental Diet Nutrition Level


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design and Animals
2.2. Growth Performance and Apparent Ileal Digestibility
2.3. RNA Extraction and qRT-PCR
2.4. Statistical Analysis
3. Results
3.1. Growth Performance and Ileal Digestibility
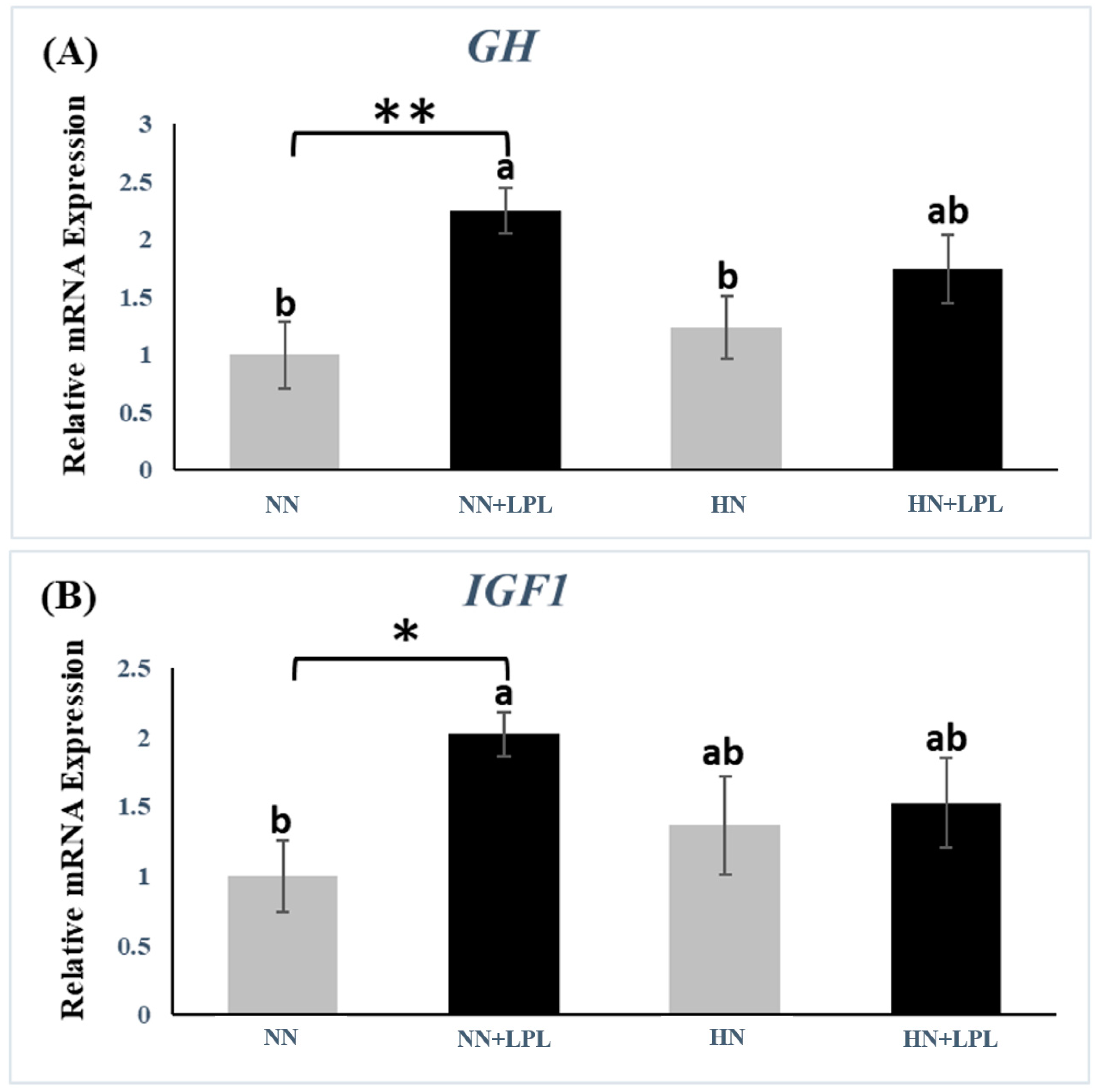
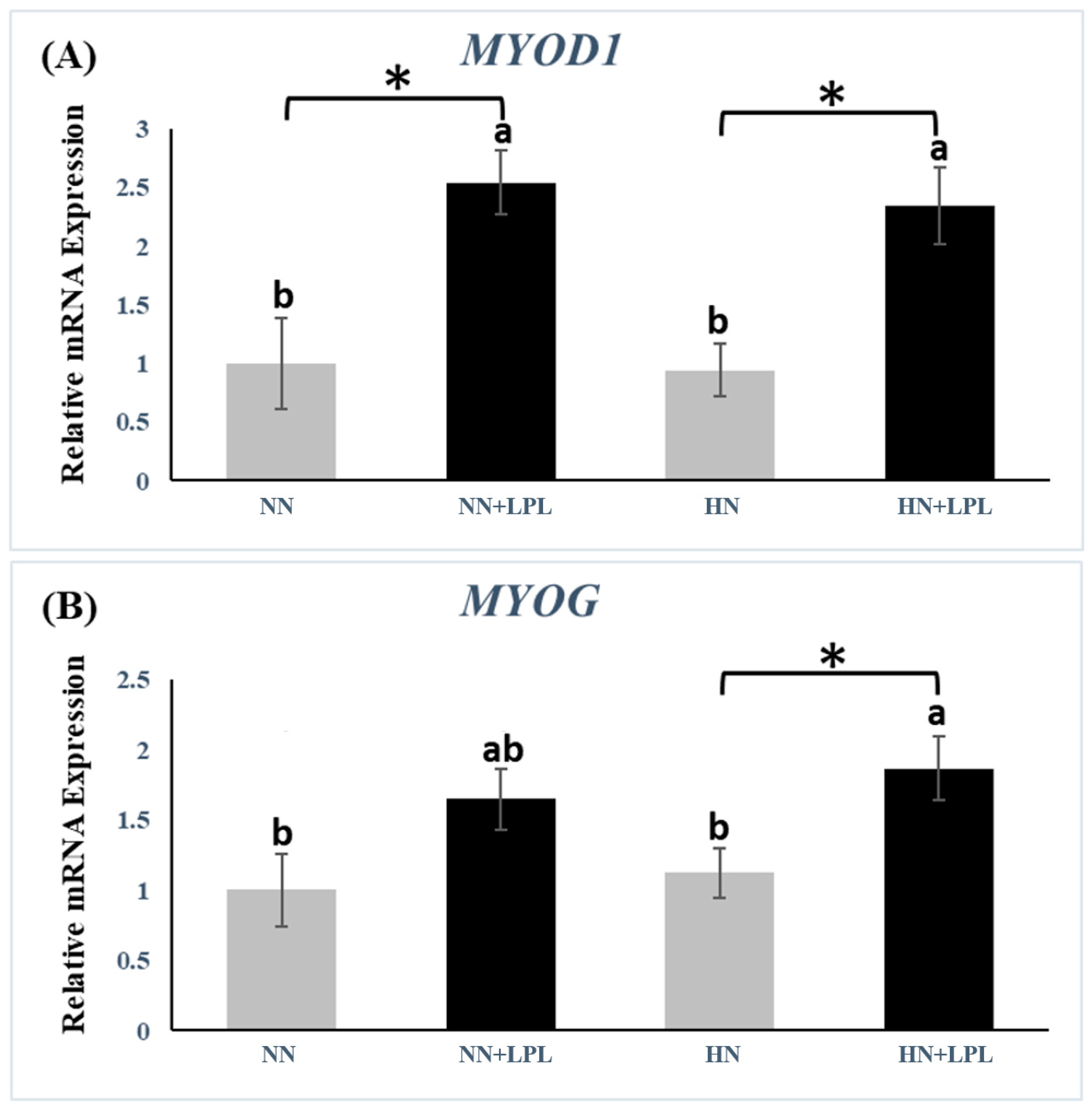
3.2. Expression of Growth-Related Genes
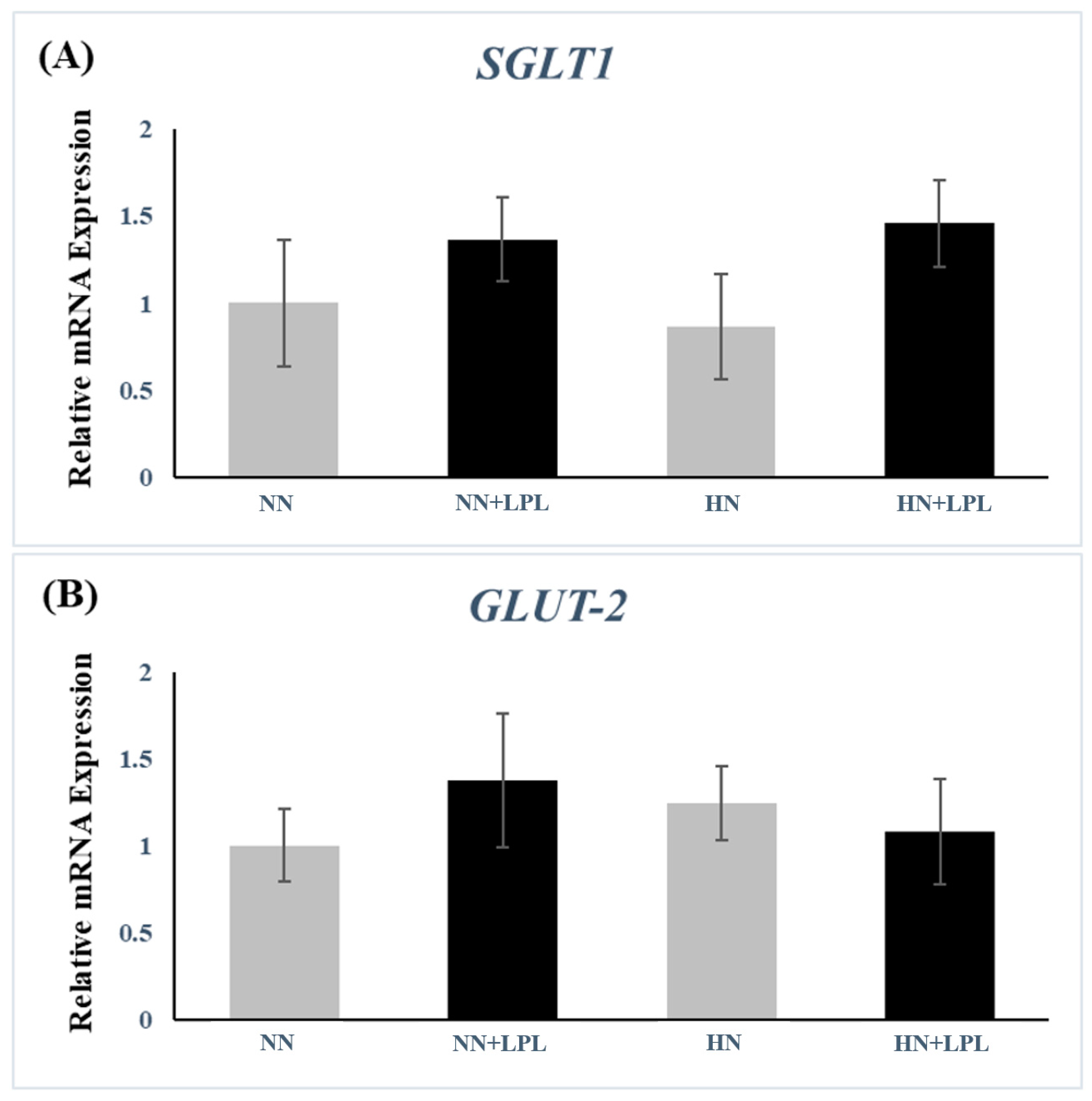
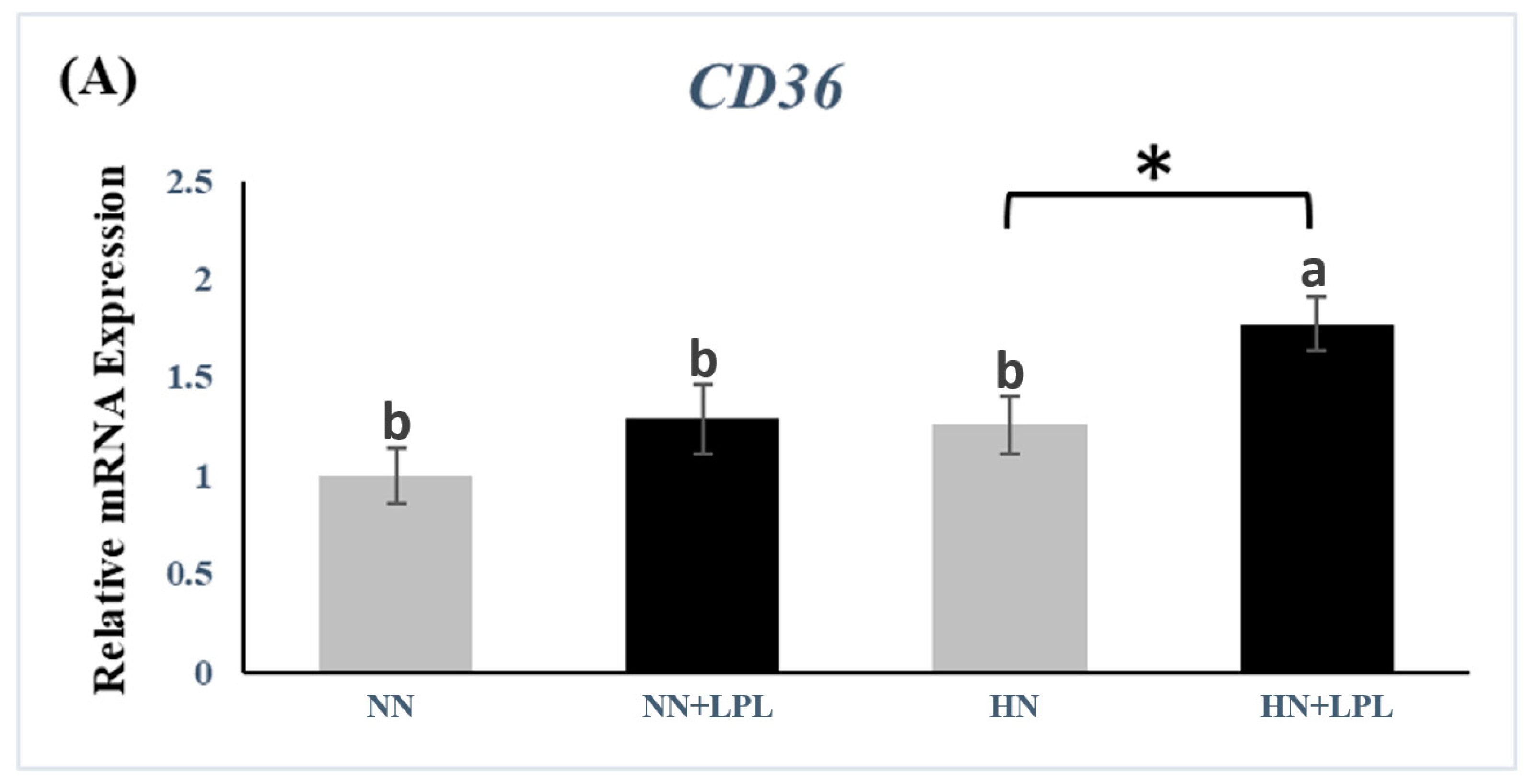
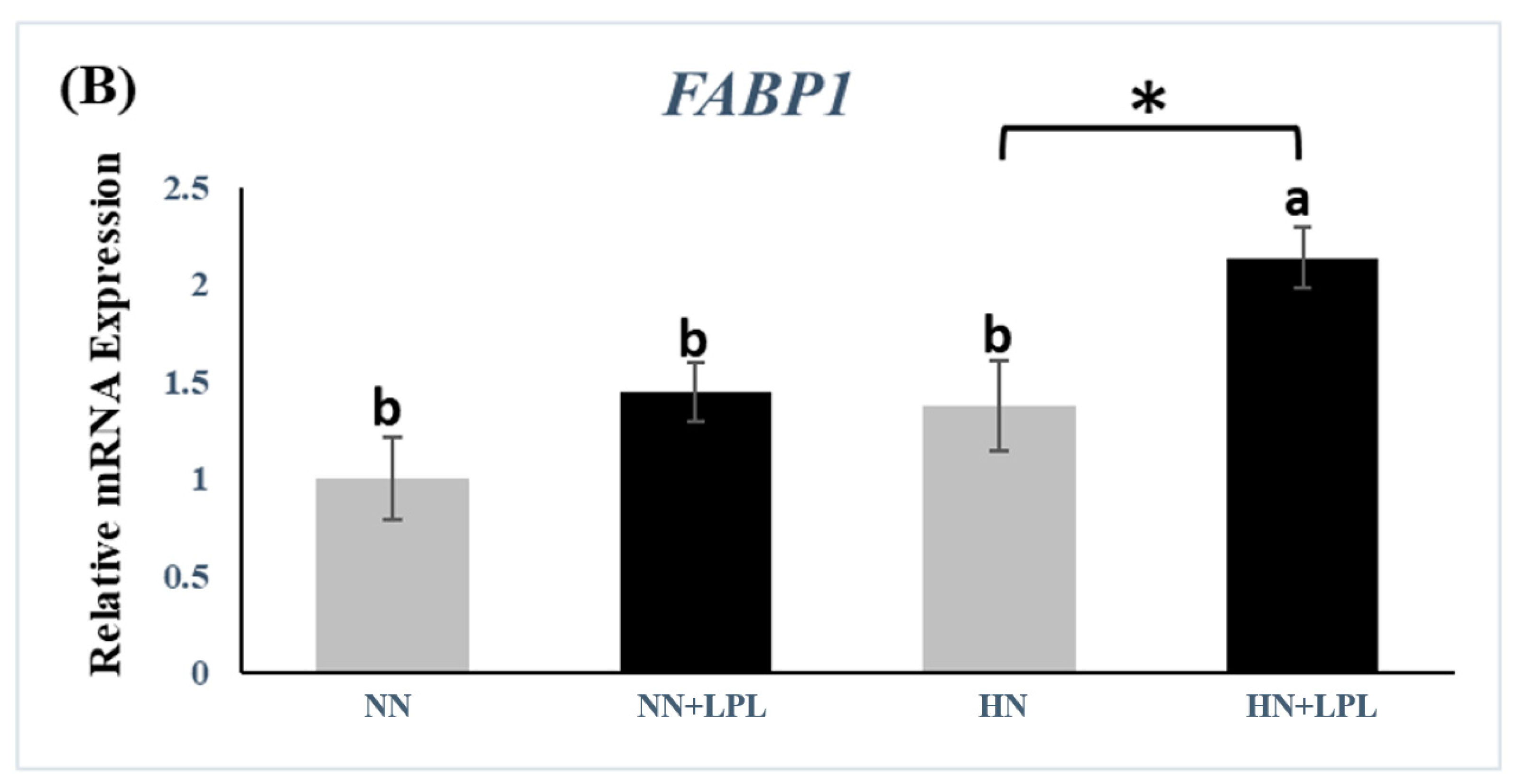
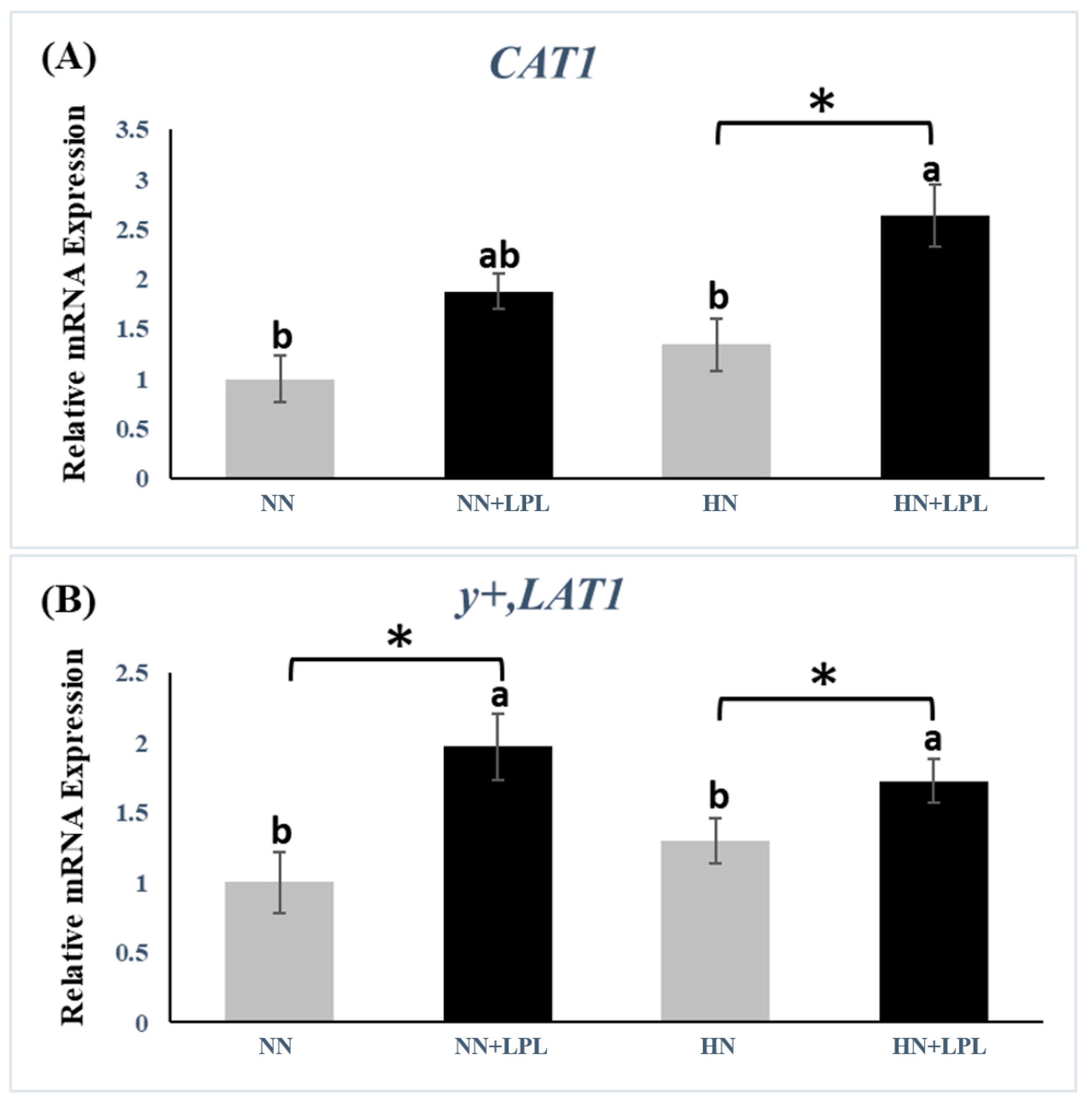
3.3. Expression of Nutrient Transporter Genes
4. Discussion
4.1. Growth Performance and Ileal Digestibility of Animo Acids
4.2. Growth-Related Genes
4.3. Nutrient Transporter Genes
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Haetinger, V.S.; Dalmoro, Y.K.; Godoy, G.L. Optimizing cost, growth performance, and nutrient absorption with a bio-emulsifier based on lysophospholipids for broiler chickens. Poult. Sci 2021, 100, 101025. [Google Scholar] [CrossRef]
- Joshi, A.; Paratkar, S.G.; Thorat, B.N. Modification of lecithin by physical, chemical and enzymatic methods. Eur. J. Lipid Sci. 2006, 108, 363–373. [Google Scholar] [CrossRef]
- Honda, K.; Kamisoyama, H.; Isshiki, Y.; Hasegawa, S. Effects of dietary fat levels on nutrient digestibility at different sites of chicken intestines. JPSA 2009, 46, 291–295. [Google Scholar] [CrossRef] [Green Version]
- Lundbæk, J.A.; Andersen, O.S. Lysophospholipids modulate channel function by altering the mechanical properties of lipid bilayers. J. Gen. Physiol. 1994, 104, 645–673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maingret, F.; Patel, A.J.; Lesage, F.; Lazdunski, M.; Honoré, E. Lysophospholipids open the two-pore domain mechanogated K(+) channels TREK-1 and TRAAK. J. Biol. Chem. 2000, 275, 10128–10133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Renaville, R.; Hammadi, M.; Portetelle, D. Role of the somatotropic axis in the mammalian metabolism. Domest. Anim. Endocrinol. 2002, 23, 351–360. [Google Scholar] [CrossRef] [PubMed]
- Kühn, E.R.; Vleurick, L.; Edery, M.; Decuypere, E.; Darras, V.M. Internalization of the chicken growth hormone receptor complex and its effect on biological functions. Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2002, 132, 299–308. [Google Scholar] [CrossRef]
- Anh, N.T.L.; Kunhareang, S.; Duangjinda, M. Association of Chicken Growth Hormones and Insulin-like Growth Factor Gene Polymorphisms with Growth Performance and Carcass Traits in Thai Broilers. Asian-Australas. J. Anim. Sci. 2015, 28, 1686–1695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.H.; Lee, S.I.; Kim, I.H. The effect of protease on growth performance, nutrient digestibility, and expression of growth-related genes and amino acid transporters in broilers. J. Anim. Sci. Technol. 2020, 62, 614–627. [Google Scholar] [CrossRef] [PubMed]
- Saxena, R.; Saxena, V.K.; Tripathi, V.; Mir, A.; Dev, K.; Begum, J.; Agarwal, R.; Goel, A. Dynamics of gene expression of hormones involved in the growth of broiler chickens in response to the dietary protein and energy changes. Gen. Comp. Endocrinol. 2020, 288, 113377. [Google Scholar] [CrossRef] [PubMed]
- Juntanapum, W.; Bunchasak, C.; Poeikhampha, T.; Rakangthong, C. Poungpong, K; Effects of supplementation of lysophosphatidylcholine (LPC) to lying hens on production performance, fat digestibility, blood lipid profile, and gene expression related to nutrients transport in small intestine. J. Anim. Feed Sci. 2020, 29, 258–265. [Google Scholar] [CrossRef]
- Brautigan, D.L.; Li, R.; Kubicka, E.; Turner, S.D.; Garcia, J.S.; Weintraut, M.L.; Wong, E.A. Lysolecithin as feed additive enhances collagen expression and villus length in the jejunum of broiler chickens. Poult. Sci. 2017, 96, 2889–2898. [Google Scholar] [CrossRef] [PubMed]
- Moreno-Navarrete, J.M.; Catalan, V.; Whyte, L.; Diaz-Arteaga, A.; Vazquez-Martinez, R.; Rotellar, F. The L-alpha-lysophosphatidylinositol/GPR55 system and its potential role in human obesity. Diabetes 2012, 61, 281–291. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Metz, S.A. Lysophosphatidylinositol, but not lysophosphatidic acid, stimulates insulin release. A possible role for phospholipase A2 but notde novosynthesis of lysophospholipid in pancreatic islet function. Biochem. Biophys. Res. Commun. 1986, 138, 720–727. [Google Scholar] [CrossRef] [PubMed]
- Menon, D.; Salloum, D.; Bernfeld, E.; Gorodetsky, E.; Akselrod, A.; Frias, M.A.; Sudderth, J.; Chen, P.H.; DeBerardinis, R.; Foster, D.A. Lipid sensing by mTOR complexes via de novo synthesis of phosphatidic acid. J. Biol. Chem. 2017, 292, 6303–6311. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoon, M.S. The Role of Mammalian Target of Rapamycin (mTOR) in Insulin Signaling. Nutrients 2017, 9, 1176. [Google Scholar] [CrossRef]
- Oka, S.; Nakajima, K.; Yamashita, A.; Kishimoto, S.; Sugiura, T. Identification of GPR55 as a lysophosphatidylinositol receptor. Biochem. Biophys. Res. Commun. 2007, 362, 928–934. [Google Scholar] [CrossRef]
- Ross, R.A. L-α-Lysophosphatidylinositol meets GPR55: A deadly relationship. Trends Pharmacol. Sci 2011, 32, 265–269. [Google Scholar] [CrossRef]
- Kimball, S.R.; Jefferson, L. New functions for amino acids: Effects on gene transcription and translation. Am. J. Clin. Ntr. 2006, 83, 500S–507S. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- CVB. CVB Table Booklet Feeding of Poultry. Voluntary Feed Intake in Pigs Feeding Standards, Feeding Advices and Nutritional Values of Feed Ingredients for Poultry; Wageningen Academic Publications: Wageningen, The Netherlands, 2018. [Google Scholar]
- Sales, J.; Janssens, G.P.J. The use of markers to determine energy metabolizability and nutrient digestibility in avian species. World’s Poult. Sci. J. 2003, 59, 314–327. [Google Scholar] [CrossRef]
- Mountzouris, K.C.; Tsitrsikos, P.; Palamidi, I.; Arvaniti, A.; Mohnl, M.; Schatzmayr, G.; Fegero, K. Effects of probiotic inclusion levels in broiler nutrition on growth performance, nutrient digestibility, plasma immunoglobulins, and cecal microflora composition. Poult. Sci. 2010, 89, 58–67. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis, 19th ed.; AOAC Int 2012; Association of Official Analytical Chemists: Washington, DC, USA, 2012.
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- SAS Institute. SAS User’s Guide: Statistics, Version 7.0 ed.; 1SAS Institute Inc.: Cary, NC, USA, 1998. [Google Scholar]
- Park, J.H.; Nguyen, D.H.; Kim, I.H. Effects of Exogenous Lysolecithin Emulsifier Supplementation on the Growth Performance, Nutrient Digestibility, and Blood Lipid Profiles of Broiler Chickens. J. Poult. Sci. 2018, 55, 190–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, P.Y.; Kim, I.H. Effect of diets with different energy and lysophospholipids levels on performance, nutrient metabolism, and body composition in broilers. Poult. Sci. 2017, 96, 1341–1347. [Google Scholar] [CrossRef]
- Wealleans, A.L.; Jansen, M.; Benedetto, M.D. The addition of lysolecithin to broiler diets improves growth performance across fat levels and sources: A meta-analysis of 33 trials. Br. Poult. Sci. 2019, 61, 51–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lundbæk, J.A.; Collingwood, S.A.; Ingólfsson, H.I.; Kapoor, R.; Andersen, O.S. Lipid bilayer regulation of membrane protein function: Gramicidin channels as molecular force probes. J. R. Soc. Interface 2010, 7, 373–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boontiam, W.; Jung, B.; Kim, Y.Y. Effects of lysophospholipid supplementation to lower nutrient diets on growth performance, intestinal morphology, and blood metabolites in broiler chickens. Poult. Sci. 2017, 96, 593–601. [Google Scholar] [CrossRef]
- Boontiam, W.; Hyun, Y.K.; Jung, B.; Kim, Y.Y. Effects of lysophospholipid supplementation to reduced energy, crude protein, and amino acid diets on growth performance, nutrient digestibility, and blood profiles in broiler chickens. Poult. Sci. 2019, 98, 6693–6701. [Google Scholar] [CrossRef]
- Giustina, A.; Veldhuis, J.D. Pathophysiology of the neuroregulation of growth hormone secretion in experimental animals and the human. Endocr. Rev. 1998, 19, 717–797. [Google Scholar]
- Frago, L.M.; Paneda, C.; Dickson, S.L.; Hewson, A.K.; Argente, J.; Chowen, J.A. Growth hormone (gh) and gh-releasing peptide-6 increase brain insulin-like growth factor-i expression and activate intracellular signaling pathways involved in neuroprotection. Endocrinology 2002, 143, 4113–4122. [Google Scholar] [CrossRef] [Green Version]
- Froesch, E.R.; Guler, H.P.; Schmid, C.; Zapf, J. Therapeutic potential of insulin-like growth factor 1. Trends. Endocrinol. Metab. 1990, 19, 254–260. [Google Scholar]
- Nam, S.Y.; Lee, E.J.; Kim, K.R.; Cha, B.S.; Song, Y.D.; Lim, S.K.; Lee, H.C.; Huh, K.B. Effect of obesity on total and free insulin-like growth factor (IGF)-1, and their relationship to IGF-binding protein (BP)-1, IGFBP-2, IGFBP-3, insulin, and growth hormone. Int. J. Obes. 1997, 21, 355–359. [Google Scholar] [CrossRef]
- Fekete, S.G.; Brown, D.L. Veterinary aspects and perspectives of nutrigenomics: A critical review. Acta Vet. Hung. 2007, 55, 229–239. [Google Scholar] [CrossRef] [PubMed]
- Lu, F.Z.; Chen, J.; Wang, X.X.; Liu, H.L. Investigation of the Insulin-like growth factor system in breast muscle during embryonic and postnatal development in langshan and arbor acres chickens subjected to different feeding regimens. Asian-Aust. J. Anim. Sci. 2009, 22, 471–482. [Google Scholar] [CrossRef]
- Megeney, L.A.; Rudnicki, M.A. Determination versus differentiation and the MyoD family of transcription factors. Biochem. Cell Biol. 1995, 73, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Devol, D.L.; Rotwein, P.; Sadow, J.L.; Novakofski, J.; Bechtel, P.J. Activation of insulin-like growth factor gene expression during work-induced skeletal muscle growth. Am. J. Physiol. Endocrinol. Metab. 1990, 259, 89–95. [Google Scholar] [CrossRef]
- Coleman, M.E.; DeMayo, F.; Yin, K.C.; Lee, H.M.; Geske, R.; Montgomery, C.; Schwartz, R.J. Myogenic vector expression of insulin-like growth factor I stimulates muscle cell differentiation and myofiber hypertrophy in transgenic mice. J. Biol. Chem. 1995, 270, 12109–12116. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.J.; Ma, J.D.; Qiu, W.L.; Zhang, J.W.; Feng, S.Y.; Zhou, X.K.; Wang, X.; Jin, L.; Long, K.; Liu, L.Y.; et al. Guanidinoacetic Acid Regulates Myogenic Differentiation and Muscle Growth Through miR-133a-3p and miR-1a-3p Co-mediated Akt/mTOR/S6K Signaling Pathway. Int. J. Mol. Sci. 2018, 19, 2837. [Google Scholar] [CrossRef] [Green Version]
- Zeng, Y.; Irvine, R.; Hiley, C. Biased signalling might be the answer to the inconsistent pharmacology of GPR55. FASEB J. 2015, 29, 772–777. [Google Scholar] [CrossRef]
- Metayer, S.; Seiliez, I.; Collin, A.; Duchêne, S.; Mercier, Y.; Geraert, P.A.; Tesseraud, S. Mechanisms through which sulfur amino acids control protein metabolism and oxidative status. J. Nutr. Biochem. 2008, 19, 207–215. [Google Scholar] [CrossRef]
- Zeitz, J.O.; Mohrmann, S.; Käding, S.C.; Devlikamov, M.; Niewalda, I.; Whelan, R.; Helmbrecht, A.; Eder, K. Effects of methionine on muscle protein synthesis and degradation pathways in broilers. J. Anim. Physiol. Anim. Nutr. 2018, 103, 191–203. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drover, V.A.; Ajmal, M.; Nassir, F.; Davidson, N.O.; Nauli, A.M.; Sahoo, D.; Tso, P.; Abumrad, N.A. CD36 deficiency impairs intestinal lipid secretion and clearance of chylomicrons from the blood. J. Clin. Investig. 2005, 115, 1290–1297. [Google Scholar] [CrossRef] [PubMed]
- Khonyoung, D.; Yamauchi, K.; Suzuki, K. Influence of dietary fat sources and lysolecithin on growth performance, visceral organ size, and histological intestinal alteration in broiler chickens. Livest. Sci. 2015, 176, 111–120. [Google Scholar] [CrossRef]
- Hatzoglou, M.; Fernandez, J.; Yaman, I.; Closs, E. Regulation of cationic amino acid transport: The story of the CAT-1 transporter. Annu. Rev. Nutr. 2004, 24, 377–399. [Google Scholar] [CrossRef]
- Mao, Z.; Zhang, W.Z. Role of mTOR in Glucose and Lipid Metabolism. Int. J. Mol. Sci. 2018, 19, 2043. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Osmanyan, A.K.; Shahab, G.H.; Reza, M.; Fisinin, V.I.; Arkhipova, A.L.; Glazko, T.T.; Kovalchuk, S.N.; Kosovsky, G.Y. Intestinal amino acid and peptide transporters in broiler are modulated by dietary amino acids and protein. Amino Acids 2017, 50, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Garcia, V.H.; Adriana, M.T.; Araiza-Piña, B.A.; Htoo, J.K.; Cervantes-Ramírez, M. Effects of dietary protein and amino acid levels on the expression of selected cationic amino acid transporters and serum amino acid concentration in growing pigs. Arch. Anim. Nutr. 2012, 66, 257–270. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient, % | Starter 1 | Grower 1 | Finisher 1 | |||
|---|---|---|---|---|---|---|
| Normal | High | Normal | High | Normal | High | |
| Corn | 59.03 | 56.25 | 58.05 | 57.27 | 61.50 | 58.72 |
| Soymeal meal (46%) | 25.24 | 27.15 | 24.18 | 24.44 | 18.92 | 20.54 |
| Hydrolyzed Render Meal (50%) | 3.00 | 3.00 | 4.00 | 4.00 | 5.00 | 5.00 |
| Corn gluten meal (60%) | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| DDGS (Maize) | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Soy oil | 1.65 | 2.54 | 3.00 | 3.52 | 3.99 | 5.18 |
| Limestone | 0.22 | 0.24 | 0.12 | 0.13 | 0.07 | 0.08 |
| Calcium hydro-phosphate | 1.15 | 1.13 | 1.06 | 1.04 | 1.02 | 1.00 |
| Salt | 0.31 | 0.31 | 0.31 | 0.31 | 0.31 | 0.31 |
| Methionine (99%, DL-Form) | 0.43 | 0.44 | 0.40 | 0.40 | 0.33 | 0.33 |
| Lysine -HCl (98.5%) | 0.50 | 0.48 | 0.43 | 0.44 | 0.44 | 0.42 |
| Threonine (98.5%) | 0.17 | 0.16 | 0.15 | 0.15 | 0.12 | 0.12 |
| Choline (60%) | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Vitamin premix 2 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Mineral premix 3 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Total | 100 | 100 | 100 | 100 | 100 | 100 |
| Calculated composition, % | ||||||
| Crude protein | 22 | 22.5 | 21 | 21.5 | 20 | 20.5 |
| Metabolism energy (kcal/kg) | 3000 | 3050 | 3100 | 3150 | 3200 | 3250 |
| Calcium | 0.95 | 0.95 | 0.9 | 0.9 | 0.85 | 0.85 |
| Available phosphorus | 0.40 | 0.40 | 0.38 | 0.38 | 0.36 | 0.36 |
| SID-Lys | 1.23 | 1.25 | 1.15 | 1.17 | 1.05 | 1.07 |
| SID-Met + SID-Cys | 0.96 | 0.98 | 0.90 | 0.91 | 0.82 | 0.83 |
| SID-Thr | 0.65 | 0.66 | 0.61 | 0.62 | 0.56 | 0.57 |
| NO. | Gene Symbol | Accession No | Primer (5′ to 3′) | |
|---|---|---|---|---|
| Forward | Reverse | |||
| 1 | SGLT1 | NM_001293240 | TTAGAGAGGTTGGAGGGTATGA | GAATCTGCTCGAGGCGTATAG |
| 2 | GLUT2 | NM_207178 | AGAGGAAACTGTGACCCGATGA | AACGAAGAGGAAGATGGCGA |
| 3 | CD36 | NM_204192 | GAAGGTCTGAGCCCAAATGA | AGGTGTCACAAGGAGGTTTAC |
| 4 | FABP1 | NM_001293240 | ACTGGCTCCAAAGAATGACCAATG | TGTCTCCGTTGAGTTCGGTCAC |
| 5 | CAT1 | NM_001145490 | CTTGATCGCTGCCTTGGCTT | CCGTAATGAAGGCCCACAGC |
| 6 | y+ LAT1 | XM_001231336 | GCCAACTAGCCAGGCGGTTA | TATCCTGCACCCGTGTTCCC |
| 7 | IGF1 | NM_001004384 | TGCTGCTTCCAGAGTTGTGACC | TGGCATATCAGTGTGGCGCT |
| 8 | GH | NM_204359 | TACGGCCTGCTGTCCTGCTT | TGTTTTTGGTGACGGGGAGG |
| 9 | MYOD1 | NM_204214 | GGCCGCCGATGACTTCTATG | TGCTCCTCCTCGTGTGGGTT |
| 10 | MYOG | NM_204184 | AGCGATGACCAGGCAGAGGA | CCAGCTCAGTTTTGGACCCG |
| Items | Normal Nutrition | High Nutrition | SEM 2 | p-Value 3 | ||||
|---|---|---|---|---|---|---|---|---|
| NN | NN + LPL | HN | HN + LPL | Feed Effect | LPL Effect | Interaction | ||
| d 1 to 7 | . | |||||||
| BWG, g | 125 | 128 | 128 | 130 | 2 | 0.3127 | 0.1617 | 0.7677 |
| FI, g | 147 | 152 | 153 | 153 | 3 | 0.091 | 0.2916 | 0.4229 |
| FCR | 1.176 | 1.188 | 1.195 | 1.177 | 0.028 | 0.8886 | 0.8925 | 0.6745 |
| d 7 to 21 | ||||||||
| BWG, g | 657 | 660 | 657 | 668 | 7 | 0.605 | 0.3309 | 0.5947 |
| FI, g | 1008 | 1017 | 1036 | 1031 | 9 | 0.1580 | 0.8706 | 0.3651 |
| FCR | 1.534 | 1.541 | 1.577 | 1.544 | 0.018 | 0.1812 | 0.4659 | 0.2156 |
| d 21 to 35 | ||||||||
| BWG, g | 945 | 965 | 974 | 988 | 16 | 0.0430 | 0.008 | 0.8686 |
| FI, g | 1749 | 1763 | 1726 | 1742 | 14 | 0.2035 | 0.3878 | 0.9607 |
| FCR | 1.851 | 1.826 | 1.772 | 1.763 | 0.021 | 0.0070 | 0.4807 | 0.8516 |
| Overall | ||||||||
| BWG, g | 1727 | 1753 | 1760 | 1786 | 16 | 0.0445 | 0.0022 | 0.9837 |
| FI, g | 2904 | 2931 | 2915 | 2925 | 18 | 0.8963 | 0.3501 | 0.6658 |
| FCR | 1.682 | 1.673 | 1.658 | 1.6385 | 0.01 | 0.0124 | 0.2079 | 0.6299 |
| Items, % | Normal Nutrition | High Nutrition | SEM 2 | p-Value 3 | ||||
|---|---|---|---|---|---|---|---|---|
| NN | NN + LPL | HN | HN + LPL | Feed Effect | LPL Effect | Interaction | ||
| day 35 | ||||||||
| Dry matter | 70.76 | 71.26 | 72.11 | 71.99 | 0.75 | 0.394 | 0.879 | 0.800 |
| Nitrogen | 68.32 | 71.68 | 69.91 | 72.19 | 1.23 | 0.345 | 0.854 | 0.717 |
| Energy | 67.37 | 70.91 | 72.06 | 71.77 | 0.71 | 0.314 | 0.855 | 0.692 |
| Fat | 86.71 | 88.10 | 85.77 | 88.08 | 0.63 | 0.550 | 0.026 | 0.567 |
| Val | 88.42 | 91.14 | 88.84 | 90.01 | 0.86 | 0.744 | 0.078 | 0.474 |
| Met | 89.91 | 91.38 | 90.64 | 92.15 | 0.8 | 0.334 | 0.058 | 0.978 |
| Ile | 86.49 | 90.14 | 86.00 | 89.84 | 0.71 | 0.689 | 0.004 | 0.926 |
| Leu | 86.64 | 89.50 | 86.42 | 89.76 | 1.63 | 0.993 | 0.092 | 0.893 |
| Thr | 88.01 | 90.38 | 87.27 | 91.56 | 0.89 | 0.879 | 0.025 | 0.506 |
| Phe | 81.51 | 84.58 | 81.33 | 84.82 | 0.78 | 0.980 | 0.005 | 0.855 |
| His | 86.35 | 87.94 | 84.88 | 88.64 | 0.59 | 0.640 | 0.002 | 0.197 |
| Lys | 87.90 | 90.42 | 86.70 | 90.60 | 0.93 | 0.667 | 0.275 | 0.565 |
| Arg | 86.94 | 89.71 | 86.42 | 89.22 | 0.61 | 0.695 | 0.034 | 0.988 |
| Trp | 78.15 | 80.00 | 76.21 | 82.80 | 1.13 | 0.795 | 0.105 | 0.160 |
| Tyr | 88.31 | 91.42 | 87.54 | 90.97 | 0.7 | 0.619 | 0.010 | 0.897 |
| Ser | 86.10 | 88.01 | 86.03 | 88.15 | 0.95 | 0.979 | 0.938 | 0.938 |
| Glu | 85.08 | 87.60 | 84.55 | 87.87 | 0.6 | 0.905 | 0.010 | 0.716 |
| Pro | 86.20 | 88.93 | 85.46 | 88.50 | 0.86 | 0.528 | 0.003 | 0.870 |
| Gly | 86.43 | 88.83 | 86.00 | 88.61 | 0.78 | 0.771 | 0.031 | 0.926 |
| Ala | 88.82 | 91.79 | 88.27 | 92.52 | 0.56 | 0.940 | 0.005 | 0.608 |
| Cys | 86.26 | 88.99 | 83.95 | 89.96 | 1.08 | 0.594 | 0.176 | 0.197 |
| Asp | 88.85 | 91.83 | 88.15 | 91.17 | 1.02 | 0.597 | 0.245 | 0.989 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Zhang, S.; Nie, K.; Zheng, H.; Luo, Z.; Kim, I.-H. Lysolecithin Improves Broiler Growth Performance through Upregulating Growth-Related Genes and Nutrient Transporter Genes Expression Independent of Experimental Diet Nutrition Level. Animals 2022, 12, 3365. https://doi.org/10.3390/ani12233365
Zhang Z, Zhang S, Nie K, Zheng H, Luo Z, Kim I-H. Lysolecithin Improves Broiler Growth Performance through Upregulating Growth-Related Genes and Nutrient Transporter Genes Expression Independent of Experimental Diet Nutrition Level. Animals. 2022; 12(23):3365. https://doi.org/10.3390/ani12233365
Chicago/Turabian StyleZhang, Zhiming, Song Zhang, Kangkang Nie, He Zheng, Zheng Luo, and In-Ho Kim. 2022. "Lysolecithin Improves Broiler Growth Performance through Upregulating Growth-Related Genes and Nutrient Transporter Genes Expression Independent of Experimental Diet Nutrition Level" Animals 12, no. 23: 3365. https://doi.org/10.3390/ani12233365
APA StyleZhang, Z., Zhang, S., Nie, K., Zheng, H., Luo, Z., & Kim, I. -H. (2022). Lysolecithin Improves Broiler Growth Performance through Upregulating Growth-Related Genes and Nutrient Transporter Genes Expression Independent of Experimental Diet Nutrition Level. Animals, 12(23), 3365. https://doi.org/10.3390/ani12233365