
Rubbing Salt in the Wound: Molecular Evolutionary Analysis of Pain-Related Genes Reveals the Pain Adaptation of Cetaceans in Seawater

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
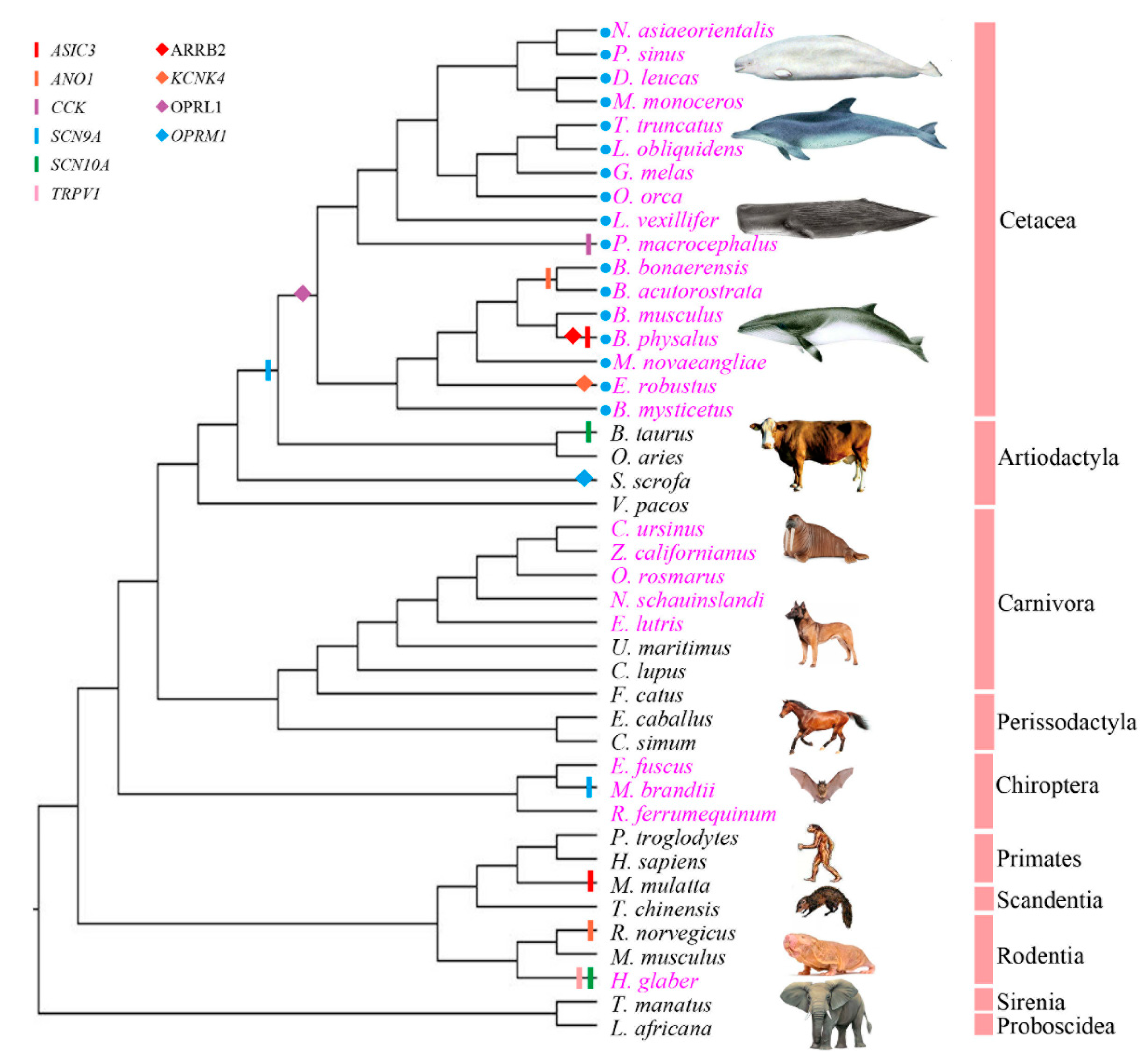
2.1. Selected Species, Candidate Genes and Sequence Acquisition
2.2. Screening for and Validating Pseudogenes
2.3. Selective Pressure Analysis
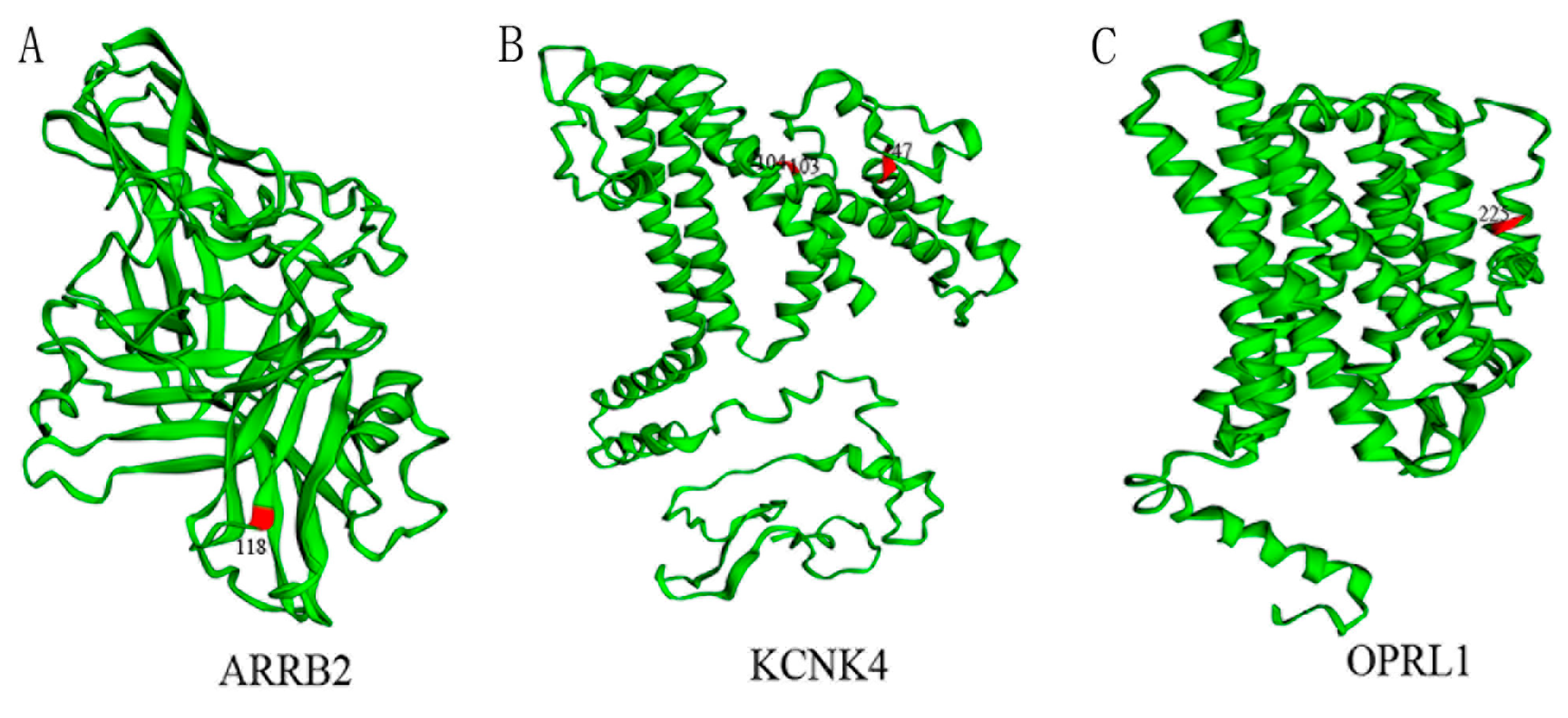
2.4. Labeling Positive Selection Sites on the Three-Dimensional Structure of Proteins
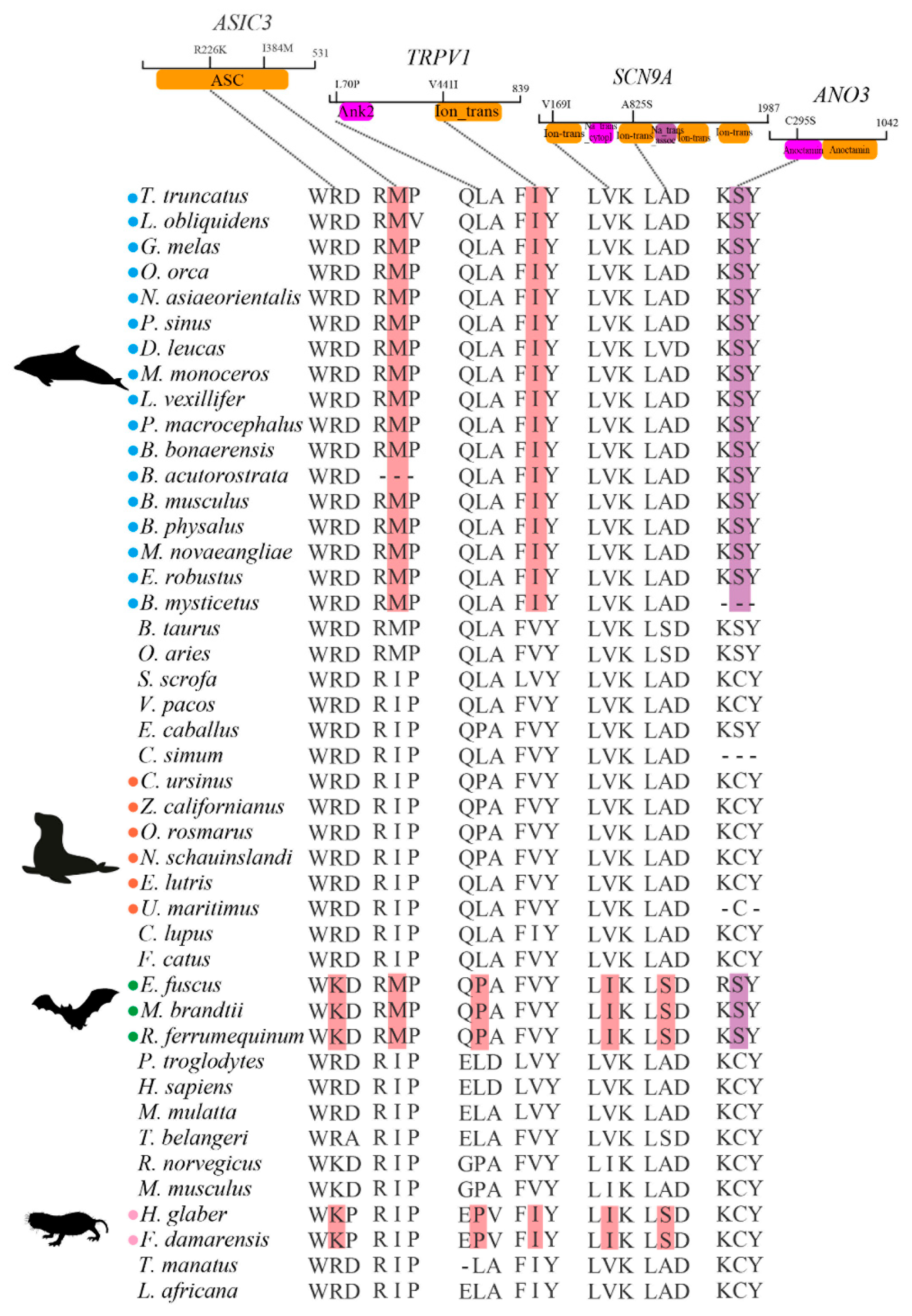
2.5. Identification of Convergent Amino Acids and Cetaceans-Specific Amino Acid Substitutions
2.6. Predicting the Effect of Cetacean-Specific Amino Acid Substitutions on Protein Function
3. Results
3.1. Pseudogenization of ASCI4 in Toothed Whales
3.2. Relaxed Selection in ASIC4 Detected in Toothed Whales
3.3. Cetacean Lineages Displayed Stronger Positive Selection than Unreported Pain-Adaption Branches and Reported Pain-Adaption Non-Cetacean Branches
3.4. Potential Molecular Convergence among Species Adapted to Pain
3.5. Cetaceans-Specific Amino Acid Substitutions
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tracey, I.; Woolf, C.J.; Andrews, N.A. Composite Pain Biomarker Signatures for Objective Assessment and Effective Treatment. Neuron 2019, 101, 783–800. [Google Scholar] [CrossRef] [Green Version]
- Apkarian, A.V.; Bushnell, M.C.; Treede, R.-D.; Zubieta, J.-K. Human brain mechanisms of pain perception and regulation in health and disease. Eur. J. Pain 2005, 9, 463. [Google Scholar] [CrossRef]
- Dubin, A.E.; Patapoutian, A. Nociceptors: The sensors of the pain pathway. J. Clin. Investig. 2010, 120, 3760–3772. [Google Scholar] [CrossRef] [Green Version]
- Ji, R.-R.; Xu, Z.-Z.; Gao, Y.-J. Emerging targets in neuroinflammation-driven chronic pain. Nat. Rev. Drug Discov. 2014, 13, 533–548. [Google Scholar] [CrossRef] [Green Version]
- Moore, C.; Gupta, R.; Jordt, S.-E.; Chen, Y.; Liedtke, W.B. Regulation of Pain and Itch by TRP Channels. Neurosci. Bull. 2018, 34, 120–142. [Google Scholar] [CrossRef] [Green Version]
- King, G.F.; Vetter, I. No Gain, No Pain: NaV1.7 as an Analgesic Target. ACS Chem. Neurosci. 2014, 5, 749–751. [Google Scholar] [CrossRef]
- Philips, H.C.; Jahanshahi, M. The components of pain behaviour report. Behav. Res. Ther. 1986, 24, 117–125. [Google Scholar] [CrossRef] [PubMed]
- Philips, H.C. Avoidance behaviour and its role in sustaining chronic pain. Behav. Res. Ther. 1987, 25, 273–279. [Google Scholar] [CrossRef]
- Gigliuto, C.; De Gregori, M.; Malafoglia, V.; Raffaeli, W.; Compagnone, C.; Visai, L.; Petrini, P.; Avanzini, M.A.; Muscoli, C.; Viganò, J.; et al. Pain assessment in animal models: Do we need further studies? J. Pain Res. 2014, 7, 227–236. [Google Scholar] [CrossRef] [Green Version]
- Cao, Y.Q.; Mantyh, P.W.; Carlson, E.J.; Gillespie, A.-M.; Epstein, C.J.; Basbaum, A.I. Primary afferent tachykinins are required to experience moderate to intense pain. Nature 1998, 392, 390–394. [Google Scholar] [CrossRef]
- Park, T.J.; Lu, Y.; Jüttner, R.; Smith, E.S.J.; Hu, J.; Brand, A.; Wetzel, C.; Milenkovic, N.; Erdmann, B.; Heppenstall, P.A.; et al. Selective inflammatory pain insensitivity in the African naked mole-rat (Heterocephalus glaber). PLoS Biol. 2008, 6, e13. [Google Scholar] [CrossRef] [PubMed]
- Omerbašić, D.; Smith, E.S.J.; Moroni, M.; Homfeld, J.; Eigenbrod, O.; Bennett, N.C.; Reznick, J.; Faulkes, C.G.; Selbach, M.; Lewin, G.R. Hypofunctional TrkA Accounts for the Absence of Pain Sensitization in the African Naked Mole-Rat. Cell Rep. 2016, 17, 748–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gurevitz, M. Mapping of scorpion toxin receptor sites at voltage-gated sodium channels. Toxicon 2012, 60, 502–511. [Google Scholar] [CrossRef]
- Hopp, B.H.; Arvidson, R.S.; Adams, M.E.; Razak, K.A. Arizona bark scorpion venom resistance in the pallid bat, Antrozous pallidus. PLoS ONE 2017, 12, e0183215. [Google Scholar] [CrossRef] [Green Version]
- Ahern, G.P.; Brooks, I.M.; Miyares, R.L.; Wang, X.-b. Extracellular Cations Sensitize and Gate Capsaicin Receptor TRPV1 Modulating Pain Signaling. J. Neurosci. 2005, 25, 5109. [Google Scholar] [CrossRef] [Green Version]
- Longhurst, H.; Cicardi, M. Hereditary angio-oedema. Lancet 2012, 379, 474–481. [Google Scholar] [CrossRef] [Green Version]
- Walker, K.A. Behavioural and Physiological Responses of Steller SEA Lions to Invasive Marking Techniques: Evidence of Post-operative Pain. Ph.D. Thesis, University of British Columbia, Vancouver, BC, Canada, 2010. [Google Scholar]
- Walker, K.A.; Mellish, J.E.; Weary, D.M. Effects of hot-iron branding on heart rate, breathing rate and behaviour of anaesthetised Steller sea lions. Vet. Rec. 2011, 169, 363. [Google Scholar] [CrossRef]
- Zasloff, M. Observations on the Remarkable (and Mysterious) Wound-Healing Process of the Bottlenose Dolphin. J. Investig. Dermatol. 2011, 131, 2503–2505. [Google Scholar] [CrossRef] [Green Version]
- Xue, Y.; Chidiac, C.; Herault, Y.; Gaveriaux-Ruff, C. Pain behavior in SCN9A (Nav1.7) and SCN10A (Nav1.8) mutant rodent models. Neurosci. Lett. 2021, 753, 135844. [Google Scholar] [CrossRef]
- Landy, M.A.; Goyal, M.; Casey, K.M.; Liu, C.; Lai, H.C. Loss of Prdm12 during development, but not in mature nociceptors, causes defects in pain sensation. Cell Rep. 2021, 34, 108913. [Google Scholar] [CrossRef]
- Liu, Y.; Liu, Z.; Wang, K. The Ca2+-activated chloride channel ANO1/TMEM16A: An emerging therapeutic target for epithelium-originated diseases? Acta Pharm. Sin. B 2021, 11, 1412–1433. [Google Scholar] [CrossRef] [PubMed]
- McRoberts, J.W. Cholecystokinin and pain: A review. Anesthesia Prog. 1986, 33, 87–90. [Google Scholar]
- Yu, L.-N.; Sun, L.-H.; Wang, M.; Yan, M. Research progress of the role and mechanism of extracellular signal-regulated protein kinase 5 (ERK5) pathway in pathological pain. J. Zhejiang Univ. Sci. B 2016, 17, 733–741. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Obata, K.; Noguchi, K. MAPK activation in nociceptive neurons and pain hypersensitivity. Life Sci. 2004, 74, 2643–2653. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Stifani, S. Roles of Runx Genes in Nervous System Development. In RUNX Proteins in Development and Cancer; Groner, Y., Ito, Y., Liu, P., Neil, J.C., Speck, N.A., van Wijnen, A., Eds.; Springer: Singapore, 2017; pp. 103–116. [Google Scholar]
- Testa, G.; Cattaneo, A.; Capsoni, S. Understanding pain perception through genetic painlessness diseases: The role of NGF and proNGF. Pharmacol. Res. 2021, 169, 105662. [Google Scholar] [CrossRef]
- Indo, Y. Genetics of congenital insensitivity to pain with anhidrosis (CIPA) or hereditary sensory and autonomic neuropathy type IV. Clin. Auton. Res. 2002, 12, I20–I32. [Google Scholar] [CrossRef]
- Coutelier, M.; Blesneac, I.; Monteil, A.; Monin, M.-L.; Ando, K.; Mundwiller, E.; Brusco, A.; Le Ber, I.; Anheim, M.; Castrioto, A.; et al. A Recurrent Mutation in CACNA1G Alters Cav3.1 T-Type Calcium-Channel Conduction and Causes Autosomal-Dominant Cerebellar Ataxia. Am. J. Hum. Genet. 2015, 97, 726–737. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.B.; Gray, J.; Gunthorpe, M.J.; Hatcher, J.P.; Davey, P.T.; Overend, P.; Harries, M.H.; Latcham, J.; Clapham, C.; Atkinson, K.; et al. Vanilloid receptor-1 is essential for inflammatory thermal hyperalgesia. Nature 2000, 405, 183–187. [Google Scholar] [CrossRef]
- Wirkner, K.; Sperlagh, B.; Illes, P. P2X3 Receptor Involvement in Pain States. Mol. Neurobiol. 2007, 36, 165–183. [Google Scholar] [CrossRef]
- Zhang, W.-J.; Luo, H.-L.; Zhu, Z.-M. The role of P2X4 receptors in chronic pain: A potential pharmacological target. Biomed. Pharmacother. 2020, 129, 110447. [Google Scholar] [CrossRef]
- Chauhan, S.A.; Sahoo, C.G.; Dikhit, R.M.; Das, P. Acid-Sensing Ion Channels Structural Aspects, Pathophysiological Importance and Experimental Mutational Data Available Across Various Species to Target Human ASIC1. Curr. Drug Targets 2019, 20, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Wemmie, J.A.; Taugher, R.J.; Kreple, C.J. Acid-sensing ion channels in pain and disease. Nat. Rev. Neurosci. 2013, 14, 461–471. [Google Scholar] [CrossRef] [PubMed]
- Fu, E.S.; Erasso, D.M.; Zhuang, G.Z.; Upadhyay, U.; Ozdemir, M.; Wiltshire, T.; Sarantopoulos, K.D.; Smith, S.B.; Maixner, W.; Martin, E.R.; et al. Impact of human CA8 on thermal antinociception in relation to morphine equivalence in mice. Neuroreport 2017, 28, 1215–1220. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Heijnen, C.J.; van Velthoven, C.T.J.; Willemen, H.L.D.M.; Ishikawa, Y.; Zhang, X.; Sood, A.K.; Vroon, A.; Eijkelkamp, N.; Kavelaars, A. Balancing GRK2 and EPAC1 levels prevents and relieves chronic pain. J. Clin. Investig. 2013, 123, 5023–5034. [Google Scholar] [CrossRef] [Green Version]
- Ozberk, D.; Haywood, A.; Sutherland, H.G.; Yu, C.; Albury, C.L.; Zunk, M.; George, R.; Good, P.; Griffiths, L.R.; Hardy, J.; et al. Association of polymorphisms in ARRB2 and clinical response to methadone for pain in advanced cancer. Pharmacogenomics 2022, 23, 281–289. [Google Scholar] [CrossRef]
- Imbrici, P.; Altamura, C.; Gualandi, F.; Mangiatordi, G.F.; Neri, M.; De Maria, G.; Ferlini, A.; Padovani, A.; D’Adamo, M.C.; Nicolotti, O.; et al. A novel KCNA1 mutation in a patient with paroxysmal ataxia, myokymia, painful contractures and metabolic dysfunctions. Mol. Cell. Neurosci. 2017, 83, 6–12. [Google Scholar] [CrossRef]
- Peng, Y.; Zhang, Q.; Cheng, H.; Yan, G.; Xing, C. Upregulation of ubiquitin conjugating enzyme E2B (Ube2b) ameliorates neuropathic pain by regulating Kcna2 (potassium voltage-gated channel subfamily A member 2) in primary afferent neurons. Bioengineered 2021, 12, 7470–7480. [Google Scholar] [CrossRef]
- Noël, J.; Zimmermann, K.; Busserolles, J.; Deval, E.; Alloui, A.; Diochot, S.; Guy, N.; Borsotto, M.; Reeh, P.; Eschalier, A.; et al. The mechano-activated K+ channels TRAAK and TREK-1 control both warm and cold perception. EMBO J. 2009, 28, 1308–1318. [Google Scholar] [CrossRef] [Green Version]
- Guo, Y.-P.; Zhi, Y.-R.; Liu, T.-T.; Wang, Y.; Zhang, Y. Global Gene Knockout of Kcnip3 Enhances Pain Sensitivity and Exacerbates Negative Emotions in Rats. Front. Mol. Neurosci. 2019, 12, 5. [Google Scholar] [CrossRef] [Green Version]
- Mannerak, M.A.; Lashkarivand, A.; Eide, P.K. Trigeminal neuralgia and genetics: A systematic review. Mol. Pain 2021, 17, 17448069211016139. [Google Scholar] [CrossRef]
- Ślęczkowska, M.; Almomani, R.; Marchi, M.; de Greef, B.T.A.; Sopacua, M.; Hoeijmakers, J.G.J.; Lindsey, P.; Salvi, E.; Bönhof, G.J.; Ziegler, D.; et al. Peripheral Ion Channel Gene Screening in Painful- and Painless-Diabetic Neuropathy. Int. J. Mol. Sci. 2022, 23, 7190. [Google Scholar] [CrossRef] [PubMed]
- Campa, D.; Gioia, A.; Tomei, A.; Poli, P.; Barale, R. Association of ABCB1/MDR1 and OPRM1 Gene Polymorphisms With Morphine Pain Relief. Clin. Pharmacol. Ther. 2008, 83, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Seo, E.-J.; Efferth, T.; Panossian, A. Curcumin downregulates expression of opioid-related nociceptin receptor gene (OPRL1) in isolated neuroglia cells. Phytomedicine 2018, 50, 285–299. [Google Scholar] [CrossRef] [PubMed]
- Ho, K.W.D.; Wallace, M.R.; Staud, R.; Fillingim, R.B. OPRM1, OPRK1, and COMT genetic polymorphisms associated with opioid effects on experimental pain: A randomized, double-blind, placebo-controlled study. Pharm. J. 2020, 20, 471–481. [Google Scholar] [CrossRef]
- Nielsen, L.M.; Christrup, L.L.; Sato, H.; Drewes, A.M.; Olesen, A.E. Genetic Influences of OPRM1, OPRD1 and COMT on Morphine Analgesia in a Multi-Modal, Multi-Tissue Human Experimental Pain Model. Basic Clin. Pharmacol. Toxicol. 2017, 121, 6–12. [Google Scholar] [CrossRef] [Green Version]
- Scornavacca, C.; Belkhir, K.; Lopez, J.; Dernat, R.; Delsuc, F.; Douzery, E.J.P.; Ranwez, V. OrthoMaM v10: Scaling-Up Orthologous Coding Sequence and Exon Alignments with More than One Hundred Mammalian Genomes. Mol. Biol. Evol. 2019, 36, 861–862. [Google Scholar] [CrossRef] [Green Version]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Green, M.R.; Sambrook, J. Molecular Cloning: A Laboratory Manual, 4th ed.; Cold Spring Harbor Laboratory Pr: New York, NY, USA, 2012; Volume 3. [Google Scholar]
- Yang, Z. PAML 4: Phylogenetic Analysis by Maximum Likelihood. Mol. Biol. Evol. 2007, 24, 1586–1591. [Google Scholar] [CrossRef] [Green Version]
- Zhao, H.; Zhou, Y.; Pinto, C.M.; Charles-Dominique, P.; Galindo-González, J.; Zhang, S.; Zhang, J. Evolution of the sweet taste receptor gene Tas1r2 in bats. Mol. Biol. Evol. 2010, 27, 2642–2650. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Suleski, M.; Hedges, S.B. TimeTree: A Resource for Timelines, Timetrees, and Divergence Times. Mol. Biol. Evol. 2017, 34, 1812–1819. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Society. Ser. B Methodol. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Yang, Z.; Wong, W.S.W.; Nielsen, R. Bayes Empirical Bayes Inference of Amino Acid Sites Under Positive Selection. Mol. Biol. Evol. 2005, 22, 1107–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Y.B. FasParser: A package for manipulating sequence data. Zool. Res. 2017, 38, 110–112. [Google Scholar] [PubMed] [Green Version]
- Adzhubei, I.A.; Schmidt, S.; Peshkin, L.; Ramensky, V.E.; Gerasimova, A.; Bork, P.; Kondrashov, A.S.; Sunyaev, S.R. A method and server for predicting damaging missense mutations. Nat. Methods 2010, 7, 248–249. [Google Scholar] [CrossRef] [Green Version]
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef] [Green Version]
- Woolf, C.J. What is this thing called pain? J. Clin. Investig. 2010, 120, 3742–3744. [Google Scholar] [CrossRef]
- Raouf, R.; Quick, K.; Wood, J.N. Pain as a channelopathy. J. Clin. Investig. 2010, 120, 3745–3752. [Google Scholar] [CrossRef]
- Reilly, M.M.; Shy, M.E. Diagnosis and new treatments in genetic neuropathies. J. Neurol. Neurosurg. Psychiatry 2009, 80, 1304. [Google Scholar] [CrossRef]
- Gründer, S.; Geisler, H.-S.; Rainier, S.; Fink, J.K. Acid-sensing ion channel (ASIC) 4 gene: Physical mapping, genomic organisation, and evaluation as a candidate for paroxysmal dystonia. Eur. J. Hum. Genet. 2001, 9, 672–676. [Google Scholar] [CrossRef]
- Donier, E.; Rugiero, F.; Jacob, C.; Wood, J.N. Regulation of ASIC activity by ASIC4—New insights into ASIC channel function revealed by a yeast two-hybrid assay. Eur. J. Neurosci. 2008, 28, 74–86. [Google Scholar] [CrossRef]
- Lingueglia, E. Acid-Sensing Ion Channels (ASICs) in pain. Biol. Aujourdhui 2014, 208, 13. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Xie, R.-G.; Gao, Y.-J.; Xu, Z.-Z.; Zhao, L.-X.; Bang, S.; Berta, T.; Park, C.-K.; Lay, M.; Chen, W.; et al. β-arrestin-2 regulates NMDA receptor function in spinal lamina II neurons and duration of persistent pain. Nat. Commun. 2016, 7, 12531. [Google Scholar] [CrossRef] [Green Version]
- Goldberg, Y.P.; MacFarlane, J.; MacDonald, M.L.; Thompson, J.; Dube, M.P.; Mattice, M.; Fraser, R.; Young, C.; Hossain, S.; Pape, T.; et al. Loss-of-function mutations in the Nav1.7 gene underlie congenital indifference to pain in multiple human populations. Clin. Genet. 2007, 71, 311–319. [Google Scholar] [CrossRef]
- Wu, M.-T.; Huang, P.-Y.; Yen, C.-T.; Chen, C.-C.; Lee, M.-J. A novel SCN9A mutation responsible for primary erythromelalgia and is resistant to the treatment of sodium channel blockers. PLoS ONE 2013, 8, e55212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, C.; Yang, Y.; te Morsche, R.H.; Drenth, J.P.H.; Politei, J.M.; Waxman, S.G.; Dib-Hajj, S.D. Familial gain-of-function Nav1.9 mutation in a painful channelopathy. J. Neurol. Neurosurg. Psychiatry 2017, 88, 233. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Models | ω | −lnL | Models Compared | 2Δ(lnL) | p Value |
|---|---|---|---|---|---|
| One ratio ω(A) | 0.087 | −11,012.355 | |||
| One ratio ω = 1(B) | 1.000 | −12,064.363 | B vs. A | 2104.017 | 0 |
| The branches with pseudogenized ASIC4 have ω2, others ω1€ | ω1 = 0.084 ω2 = 0.200 | −11,008.339 | A vs. C | 8.031 | 0.004 |
| The branches with pseudogenized ASIC4 have ω2 = 1, others ω1(D) | ω1 = 0.084 ω2 = 1.000 | −11,022.971 | D vs. C | 29.265 | 6.31221 × 10−8 |
| Each branch has its own €(E) | −10,906.493 | C vs. E | 203.692 | 6.15578 × 10−12 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ding, X.; Yu, F.; He, X.; Xu, S.; Yang, G.; Ren, W. Rubbing Salt in the Wound: Molecular Evolutionary Analysis of Pain-Related Genes Reveals the Pain Adaptation of Cetaceans in Seawater. Animals 2022, 12, 3571. https://doi.org/10.3390/ani12243571
Ding X, Yu F, He X, Xu S, Yang G, Ren W. Rubbing Salt in the Wound: Molecular Evolutionary Analysis of Pain-Related Genes Reveals the Pain Adaptation of Cetaceans in Seawater. Animals. 2022; 12(24):3571. https://doi.org/10.3390/ani12243571
Chicago/Turabian StyleDing, Xiaoyue, Fangfang Yu, Xiaofang He, Shixia Xu, Guang Yang, and Wenhua Ren. 2022. "Rubbing Salt in the Wound: Molecular Evolutionary Analysis of Pain-Related Genes Reveals the Pain Adaptation of Cetaceans in Seawater" Animals 12, no. 24: 3571. https://doi.org/10.3390/ani12243571
APA StyleDing, X., Yu, F., He, X., Xu, S., Yang, G., & Ren, W. (2022). Rubbing Salt in the Wound: Molecular Evolutionary Analysis of Pain-Related Genes Reveals the Pain Adaptation of Cetaceans in Seawater. Animals, 12(24), 3571. https://doi.org/10.3390/ani12243571