
An Updated Review of Hypotheses Regarding Bat Attraction to Wind Turbines

 , and
, and 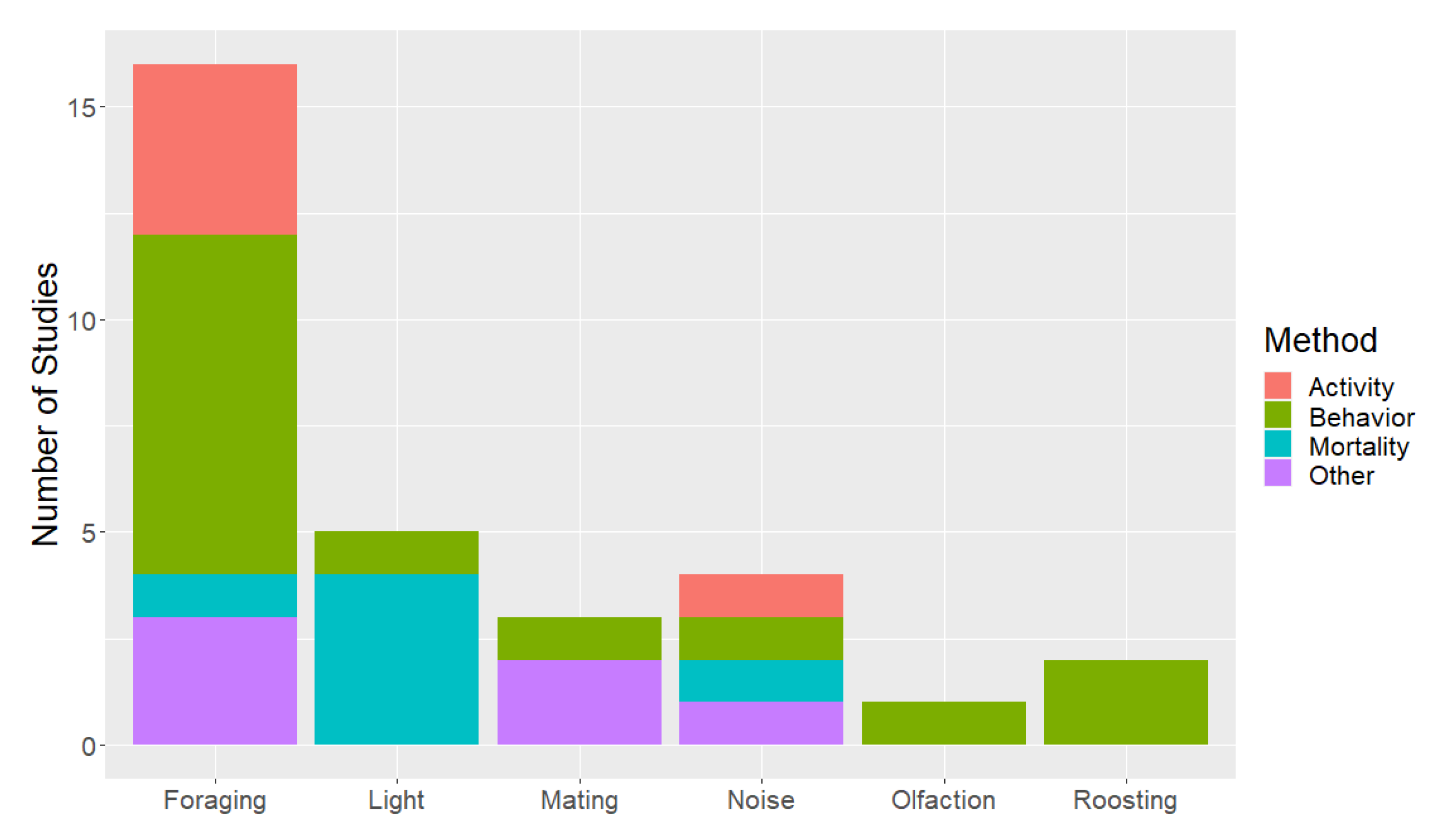{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results and Discussion
3.1. Noise
3.2. Roosting
3.3. Foraging and Water
3.4. Mating
3.5. Lights
3.6. Olfaction
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- International Energy Agency Renewables 2020: Analysis and Forecast to 2025. Available online: https://www.iea.org/reports/renewables-2020/wind (accessed on 8 March 2021).
- Lee, J.; Zhao, F. Global Wind Report; GWEC: Brussels, Belgium, 2019. [Google Scholar]
- Allison, T.D.; Diffendorfer, J.E.; Baerwald, E.F.; Beston, J.A.; Drake, D.; Hale, A.M.; Hein, C.D.; Huso, M.M.; Loss, S.R.; Lovich, J.E.; et al. Impacts to wildlife of wind energy siting and operation in the United States. Issues Ecol. 2019, 21, 2–18. [Google Scholar]
- O’Shea, T.J.; Cryan, P.M.; Hayman, D.T.S.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mammal Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef] [PubMed]
- Arnett, E.B.; Erickson, W.P.; Kerns, J.; Horn, J. Relationships between bats and wind turbines in Pennsylvania and West Virginia: An assessment of bat fatality search protocols, patterns of fatality, and behavioral interactions with wind turbines. In A Final Report Submitted to the Bats and Wind Energy Cooperative; Bat Conservation International: Austin, TX, USA, 2005. [Google Scholar]
- Zimmerling, J.R.; Francis, C.M. Bat mortality due to wind turbines in Canada. J. Wildl. Manag. 2016, 80, 1360–1369. [Google Scholar] [CrossRef]
- American Wind Wildlife Institute Summary of Bat Fatality Monitoring Data Contained in AWWIC (2nd Edition). Available online: https://awwi.org/resources/awwic-bat-technical-report/ (accessed on 16 April 2021).
- Choi, D.Y.; Wittig, T.W.; Kluever, B.M. An evaluation of bird and bat mortality at wind turbines in the Northeastern United States. PLoS ONE 2020, 15, e0238034. [Google Scholar] [CrossRef]
- Endangered Species Recovery Committee and State of Hawaii Department of Land and Natural Resources Division of Forestry and Wildlife. Hawaiian Hoary Bat Guidance for Wind Energy Projects; (First edition September 2015); Endangered Species Recovery Committee and State of Hawaii Department of Land and Natural Resources Division of Forestry and Wildlife: Honolulu, HI, USA, 2021.
- Erickson, R.A.; Thogmartin, W.E.; Diffendorfer, J.E.; Russell, R.E.; Szymanski, J.A. Effects of wind energy generation and white-nose syndrome on the viability of the indiana bat. PeerJ 2016, 4, e2830. [Google Scholar] [CrossRef] [PubMed]
- Gorresen, P.M.; Cryan, P.M.; Tredinnick, G. Hawaiin Hoary Bat (Lasiurus cinereus semotus) Behavior at Wind Turbines on Maui; University of Hawai ‘i at Hilo: Hilo, HI, USA, 2020. [Google Scholar]
- Arnett, E.B.; Baerwald, E.F.; Mathews, F.; Rodrigues, L.; Rodriguez-Duran, A.; Rydell, J.; Villegas-Patraca, R.; Voigt, C.C. Impacts of wind energy development on bats: A global perspective. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer Nature: Berlin/Heidelberg, Germany, 2016; pp. 295–323. ISBN 978-3-319-25220-9. [Google Scholar]
- Kunz, T.H.; Arnett, E.B.; Cooper, B.M.; Erickson, W.P.; Larkin, R.P.; Mabee, T.; Morrison, M.L.; Strickland, M.D.; Szewczak, J.M. Assessing impacts of wind-energy development on nocturnally active birds and bats: A guidance document. J. Wildl. Manag. 2007, 71, 2449–2486. [Google Scholar] [CrossRef]
- Frick, W.F.; Cheng, T.L.; Langwig, K.E.; Hoyt, J.R.; Janicki, A.F.; Parise, K.L.; Foster, J.T.; Kilpatrick, A.M. Pathogen dynamics during invasion and establishment of white-nose syndrome explain mechanisms of host persistence. Ecology 2017, 98, 624–631. [Google Scholar] [CrossRef] [Green Version]
- Russell, R.E.; Tinsley, K.; Erickson, R.A.; Thogmartin, W.E.; Szymanski, J. Estimating the spatial distribution of wintering little brown bat populations in the Eastern United States. Ecol. Evol. 2014, 4, 3746–3754. [Google Scholar] [CrossRef]
- Frick, W.F.; Baerwald, E.F.; Pollock, J.F.; Barclay, R.M.R.; Szymanski, J.A.; Weller, T.J.; Russell, A.L.; Loeb, S.C.; Medellin, R.A.; McGuire, L.P. Fatalities at wind turbines may threaten population viability of a migratory bat. Biol. Conserv. 2017, 209, 172–177. [Google Scholar] [CrossRef]
- Horn, J.W.; Arnett, E.B.; Kunz, T.H. Behavioral responses of bats to operating wind turbines. J. Wildl. Manag. 2008, 72, 123–132. [Google Scholar] [CrossRef]
- Cryan, P.M.; Gorresen, P.M.; Hein, C.D.; Schirmacher, M.R.; Diehl, R.H.; Huso, M.M.; Hayman, D.T.S.; Fricker, P.D.; Bonaccorso, F.J.; Johnson, D.H.; et al. Behavior of bats at wind turbines. Proc. Natl. Acad. Sci. USA 2014, 111, 15126–15131. [Google Scholar] [CrossRef] [Green Version]
- Goldenberg, S.Z.; Cryan, P.M.; Gorresen, P.M.; Fingersh, L.J. Behavioral patterns of bats at a wind turbine confirm seasonality of fatality risk. Ecol. Evol. 2021, 11, 4843–4853. [Google Scholar] [CrossRef] [PubMed]
- Richardson, S.M.; Lintott, P.R.; Hosken, D.J.; Economou, T.; Mathews, F. Peaks in bat activity at turbines and the implications for mitigating the impact of wind energy developments on bats. Sci. Rep. 2021, 11, 3636. [Google Scholar] [CrossRef] [PubMed]
- Arnett, E.B.; Brown, W.K.; Erickson, W.P.; Fiedler, J.K.; Hamilton, B.L.; Henry, T.H.; Jain, A.; Johnson, G.D.; Kerns, J.; Koford, R.R.; et al. Patterns of bat fatalities at wind energy facilities in North America. J. Wildl. Manag. 2008, 72, 61–78. [Google Scholar] [CrossRef]
- Arnett, E.B.; Baerwald, E.F. Impacts of Wind Energy Development on Bats: Implications for Conservation; Adams, R.A., Pedersen, S.C., Eds.; Springer: New York, NY, USA, 2013; ISBN 978-1-4614-7396-1. [Google Scholar]
- Barclay, R.M.R.; Baerwald, E.F.; Gruver, J.C. Variation in bat and bird fatalities at wind energy facilities: Assessing the effects of rotor size and tower height. Can. J. Zool. 2007, 85, 381–387. [Google Scholar] [CrossRef]
- Georgiakakis, P.; Kret, E.; Cárcamo, B.; Doutau, B.; Kafkaletou-Diez, A.; Vasilakis, D.; Papadatou, E. Bat fatalities at wind farms in North-Eastern Greece. Acta Chiropterologica 2012, 14, 459–468. [Google Scholar] [CrossRef]
- Baerwald, E.F.; Barclay, R.M.R. Geographic variation in activity and fatality of migratory bats at wind energy facilities. J. Mammal. 2009, 90, 1341–1349. [Google Scholar] [CrossRef]
- Electric Power Research Institute. Relationship between Bat Fatality Rates and Turbine Size at Wind Farms across the Continental U.S. and Southern Canada: An Investigation into the Occurrence of Bat Fatalities in Relation to Turbine Size; Electric Power Research Institute: Palo Alto, CA, USA, 2020. [Google Scholar]
- Thompson, M.; Beston, J.A.; Etterson, M.; Diffendorfer, J.E.; Loss, S.R. Factors associated with bat mortality at wind energy facilities in the United States. Biol. Conserv. 2017, 215, 241–245. [Google Scholar] [CrossRef]
- Fiedler, J.K.; Henry, T.H.; Tankersley, R.D.; Nicholson, C.P. Results of Bat and Bird Mortality Monitoring at the Expanded Buffalo Mountain Wind Farm, 2005; Tennessee Valley Authority: Knoxville, TN, USA, 2007. [Google Scholar]
- Huso, M.; Conkling, T.; Dalthorp, D.; Davis, M.; Smith, H.; Fesnock, A.; Katzner, T. Relative energy production determines effect of repowering on wildlife mortality at wind energy facilities. J. Appl. Ecol. 2021, 58, 1284–1290. [Google Scholar] [CrossRef]
- Lintott, P.R.; Richardson, S.M.; Hosken, D.J.; Fensome, S.A.; Mathews, F. Ecological impact assessments fail to reduce risk of bat casualties at wind farms. Curr. Biol. 2016, 26, R1135–R1136. [Google Scholar] [CrossRef] [Green Version]
- Solick, D.; Pham, D.; Nasman, K.; Bay, K. Bat activity rates do not predict bat fatality rates at wind energy facilities. Acta Chiropterologica 2020, 22, 135. [Google Scholar] [CrossRef]
- Cryan, P.M.; Brown, A.C. Migration of bats past a remote island offers clues toward the problem of bat fatalities at wind turbines. Biol. Conserv. 2007, 139, 1–11. [Google Scholar] [CrossRef]
- Cryan, P.M.; Barclay, R.M.R. Causes of bat fatalities at wind turbines: Hypotheses and predictions. J. Mammal. 2009, 90, 1330–1340. [Google Scholar] [CrossRef] [Green Version]
- Buchler, E.R.; Childs, S.B. Orientation to distant sounds by foraging big brown bats (Eptesicus fuscus). Anim. Behav. 1981, 29, 428–432. [Google Scholar] [CrossRef]
- Ahlen, I. Wind Turbines and Bats—A Pilot Study. Report Prepared for the Swedish National Energy Administration. 2003. Available online: https://docs.wind-watch.org/Ahlen-windturbines-bats-2003.pdf (accessed on 15 May 2020).
- Szewczak, J.; Arnett, E. Ultrasound Emissions from Wind Turbines as a Potential Attractant to Bats: A Preliminary Investigation. Report by Bat Conservation International; 2006. Available online: https://tethys.pnnl.gov/publications/ultrasound-emissions-wind-turbines-potential-attractant-bats-preliminary-investigation (accessed on 15 May 2020).
- Long, C.V. The Interaction of Bats (Microchiroptera) with Wind Turbines: Bioacoustic and Other Investigations; Loughborough University: Loughborough, UK, 2011. [Google Scholar]
- Rydell, J.; Wickman, A. Bat activity at a small wind turbine in the Baltic Sea. Acta Chiropterologica 2015, 17, 359–364. [Google Scholar] [CrossRef]
- Cryan, P.M. Mating behavior as a possible cause of bat fatalities at wind turbines. J. Wildl. Manag. 2008, 72, 845–849. [Google Scholar] [CrossRef]
- Crampton, L.H.; Barclay, R.M.R. Selection of roosting and foraging habitat by bats in different-aged aspen mixedwood stands. Conserv. Biol. 1998, 12, 1347–1358. [Google Scholar] [CrossRef]
- Kalcounis-rüppell, M.C.; Psyllakis, J.M.; Brigham, R.M.; Kalcounis-Jlippell, M.C.; Psyllakis, J.M.; Brigham, R.M. Tree roost selection by bats: An empirical synthesis using meta-analysis. Wildl. Soc. Bull. 2005, 33, 1123–1132. [Google Scholar] [CrossRef]
- Naďo, L.; Kaňuch, P. Swarming behaviour associated with group cohesion in tree-dwelling bats. Behav. Processes 2015, 120, 80–86. [Google Scholar] [CrossRef]
- Bennett, V.J.; Hale, A.M.; Williams, D.A. When the excrement hits the fan: Fecal surveys reveal species-specific bat activity at wind turbines. Mamm. Biol. 2017, 87, 125–129. [Google Scholar] [CrossRef]
- McAlexander, A. Evidence That Bats Perceive Wind Turbine Surfaces to Be Water. M.S. Thesis, Texas Christian University, Fort Worth, TX, USA, 2013. [Google Scholar]
- Long, C.V.; Flint, J.A.; Lepper, P.A. Insect attraction to wind turbines: Does colour play a role? Eur. J. Wildl. Res. 2011, 57, 323–331. [Google Scholar] [CrossRef] [Green Version]
- Grof-Tisza, P.; Steel, Z.; Cole, E.M.; Holyoak, M.; Karban, R. Testing predictions of movement behaviour in a hilltopping moth. Anim. Behav. 2017, 133, 161–168. [Google Scholar] [CrossRef]
- Brinkmann, R.; Bontadina, F. Survey of Possible Operational Impacts on Bats by Wind Facilities in Southern Germany. Report for Administrative District of Freiburg–Department 56, Conservation and Landscape Management; 2006. Available online: https://tethys.pnnl.gov/sites/default/files/publications/Brinkmann-2006.pdf (accessed on 15 April 2020).
- Alcock, J.; Dodson, G. The diverse mating systems of hilltopping insects. Am. Entomol. 2008, 54, 80–87. [Google Scholar] [CrossRef] [Green Version]
- Skevington, J.H. Hilltopping. In Encyclopedia of Entomology; Spring: Berlin/Heidelberg, Germany, 2008; pp. 1799–1807. ISBN 9781402063596. [Google Scholar]
- Rydell, J.; Bogdanowicz, W.; Boonman, A.; Pettersson, S.; Suchecka, E.; Pomorski, J.J. Bats may eat diurnal flies that rest on wind turbines. Mamm. Biol. 2016, 81, 331–339. [Google Scholar] [CrossRef]
- Jansson, S.; Malmqvist, E.; Brydegaard, M.; Åkesson, S.; Rydell, J. A scheimpflug lidar used to observe insect swarming at a wind turbine. Ecol. Indic. 2020, 117, 106578. [Google Scholar] [CrossRef]
- McCracken, G.F.; Gillam, E.H.; Westbrook, J.K.; Lee, Y.F.; Jensen, M.L.; Balsley, B.B. Article navigation brazilian free-tailed bats (Tadarida brasiliensis: Molossidae, Chiroptera) at high altitude: Links to migratory insect populations. Integr. Comp. Biol. 2008, 48, 107–118. [Google Scholar] [CrossRef] [Green Version]
- Rydell, J.; Bach, L.; Dubourg-Savage, M.J.; Green, M.; Rodrigues, L.; Hedenström, A. Mortality of bats at wind turbines links to nocturnal insect migration? Eur. J. Wildl. Res. 2010, 56, 823–827. [Google Scholar] [CrossRef] [Green Version]
- Krauel, J.J.; Ratcliffe, J.M.; Westbrook, J.K.; McCracken, G.F. Brazilian free-tailed bats (Tadarida brasiliensis) adjust foraging behaviour in response to migratory moths. Can. J. Zool. 2018, 96, 513–520. [Google Scholar] [CrossRef] [Green Version]
- McGuire, L.P.; Guglielmo, C.G.; Mackenzie, S.A.; Taylor, P.D. Migratory stopover in the long-distance migrant silver-haired bat, Lasionycteris noctivagans. J. Anim. Ecol. 2012, 81, 377–385. [Google Scholar] [CrossRef]
- Samoray, S.T.; Cotham, S.N.; Gumbert, M.W. Spring migration behavior of a Perimyotis subflavus (tri-colored bat) from Tennessee. Southeast. Nat. 2019, 18, N16–N20. [Google Scholar] [CrossRef]
- Jonasson, K.A.; Guglielmo, C.G. Evidence for spring stopover refuelling in migrating silver-haired bats (Lasionycteris noctivagans). Can. J. Zool. 2019, 97, 961–970. [Google Scholar] [CrossRef]
- Voigt, C.C.; Sörgel, K.; Šuba, J.; Keišs, O.; Petersons, G. The insectivorous bat Pipistrellus nathusii uses a mixed-fuel strategy to power autumn migration. Proc. R. Soc. B Biol. Sci. 2012, 279, 3772–3778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ahlén, I.; Baagøe, H.J.; Bach, L. Behavior of scandinavian bats during migration and foraging at sea. J. Mammal. 2009, 90, 1318–1323. [Google Scholar] [CrossRef] [Green Version]
- Fiedler, J.K. Assessment of Bat Mortality and Activity at Buffalo Mountain Windfarm, Eastern Tennessee; University of Tennessee: Knoxville, TN, USA, 2004. [Google Scholar]
- Taber, A.; Butryn, R. AWWI Technical Report: A Summary of Bat Fatality Data in a Nationwide Database; American Wind and Wildlife Institute: Washington, DC, USA, 2018. [Google Scholar]
- Jameson, J.W.; Willis, C.K.R. Activity of tree bats at anthropogenic tall structures: Implications for mortality of bats at wind turbines. Anim. Behav. 2014, 97, 145–152. [Google Scholar] [CrossRef]
- Foo, C.F.; Bennett, V.J.; Hale, A.M.; Korstian, J.M.; Schildt, A.J.; Williams, D.A. Increasing evidence that bats actively forage at wind turbines. PeerJ 2017, 5, e3985. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.C.; Roeleke, M.; Marggraf, L.; Petersons, G.; Voigt-Heucke, S.L. Migratory bats respond to artificial green light with positive phototaxis. PLoS ONE 2017, 12, e0177748. [Google Scholar] [CrossRef]
- Reimer, J.P.; Baerwald, E.F.; Barclay, R.M.R.R. Echolocation activity of migratory bats at a wind energy facility: Testing the feeding-attraction hypothesis to explain fatalities. J. Mammal. 2018, 99, 1472–1477. [Google Scholar] [CrossRef]
- Melcon, M.L.; Denzinger, A.; Schnitzler, H.-U. Aerial hawking and landing: Approach behaviour in Natterer’s bats, Myotis nattereri (Kuhl 1818). J. Exp. Biol. 2007, 210, 4457–4464. [Google Scholar] [CrossRef] [Green Version]
- Griffiths, S.R. Echolocating bats emit terminal phase buzz calls while drinking on the wing. Behav. Process. 2013, 98, 58–60. [Google Scholar] [CrossRef]
- Adams, A.M. Assessing and Analyzing Bat Activity with Acoustic Monitoring: Challenges and Interpretations. Ph.D. Thesis, The University of Western Ontario, London, ON, Canada, 2013. [Google Scholar]
- Smallwood, K.S.; Bell, D.A. Relating bat passage rates to wind turbine fatalities. Diversity 2020, 12, 84. [Google Scholar] [CrossRef] [Green Version]
- Valdez, E.W.; Cryan, P.M. Insect prey eaten by hoary bats (Lasiurus cinereus) prior to fatal collisions with wind turbines. West. N. Am. Nat. 2013, 73, 516–524. [Google Scholar] [CrossRef] [Green Version]
- Reimer, J.P.; Baerwald, E.F.; Barclay, R.M.R. Diet of hoary (Lasiurus cinereus) and silver-haired (Lasionycteris noctivagans) bats while migrating through Southwestern Alberta in late summer and autumn. Am. Midl. Nat. 2010, 164, 230–237. [Google Scholar] [CrossRef]
- Greif, S.; Siemers, B.M. Innate recognition of water bodies in echolocating bats. Nat. Commun. 2010, 1, 107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Russo, D.; Cistrone, L.; Jones, G. Sensory ecology of water detection by bats: A field experiment. PLoS ONE 2012, 7, e48144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walsh, A.L.; Harris, S. Foraging Habitat preferences of Vespertilionid bats in Britain. J. Appl. Ecol. 1996, 33, 508–518. [Google Scholar] [CrossRef] [Green Version]
- Morris, A.D.; Miller, D.A.; Kalcounis-Rueppell, M.C. Use of forest edges by bats in a managed pine forest landscape. J. Wildl. Manag. 2010, 74, 26–34. [Google Scholar] [CrossRef]
- Johnson, G.D.; Erickson, W.P.; Strickland, M.D.; Shepherd, M.F.; Shepherd, D.A.; Sarappo, S.A. Mortality of bats at large scale wind power development Buffalo Ridge, Minnesota. Am. Midl. Nat. 2003, 150, 332–342. [Google Scholar] [CrossRef]
- Grodsky, S.M.; Behr, M.J.; Gendler, A.; Drake, D.; Dieterle, B.D.; Rudd, R.J.; Walrath, N.L. Investigating the causes of death for wind turbine-associated bat fatalities. J. Mammal. 2011, 92, 917–925. [Google Scholar] [CrossRef] [Green Version]
- Korstian, J.M.; Hale, A.M.; Bennett, V.J.; Williams, D.A. Advances in sex determination in bats and its utility in wind-wildlife studies. Mol. Ecol. Resour. 2013, 13, 776–780. [Google Scholar] [CrossRef]
- Hein, C.D.; Schirmacher, M.R. Impact of wind energy on bats: A summary of our current knowledge. Hum.-Wildl. Interact. 2016, 10, 19–27. [Google Scholar] [CrossRef]
- Chipps, A.S.; Hale, A.M.; Weaver, S.P.; Williams, D.A. Genetic approaches are necessary to accurately understand bat-wind turbine impacts. Diversity 2020, 12, 236. [Google Scholar] [CrossRef]
- Cryan, P.M.; Jameson, J.W.; Baerwald, E.F.; Willis, C.K.R.; Barclay, R.M.R.; Snider, E.A.; Crichton, E.G. Evidence of late-summer mating readiness and early sexual maturation in migratory tree-roosting bats found dead at wind turbines. PLoS ONE 2012, 7, e47586. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weaver, S.P.; Hein, C.D.; Simpson, T.R.; Evans, J.W.; Castro-Arellano, I. Ultrasonic acoustic deterrents significantly reduce bat fatalities at wind turbines. Glob. Ecol. Conserv. 2020, 24, e01099. [Google Scholar] [CrossRef]
- Keeley, A.T.H.; Keeley, B.W. The mating system of Tadarida brasiliensis (Chiroptera: Molossidae) in a large highway bridge colony. J. Mammal. 2004, 85, 113–119. [Google Scholar] [CrossRef] [Green Version]
- Voigt, C.C.; Rehnig, K.; Lindecke, O.; Pētersons, G. Migratory bats are attracted by red light but not by warm-white light: Implications for the protection of nocturnal migrants. Ecol. Evol. 2018, 8, 9353–9361. [Google Scholar] [CrossRef]
- Rowse, E.G.; Lewanzik, D.; Stone, E.L.; Harris, S.; Jones, G. Dark matters: The effects of artifical lighting on bats. In Bats in the Anthropocene: Conservation of Bats in a Changing World; Voigt, C.C., Kingston, T., Eds.; Springer Open: Berlin/Heidelberg, Germany, 2016; pp. 187–213. ISBN 978-3-319-25220-9. [Google Scholar]
- Stone, E.L.; Jones, G.; Harris, S. Conserving energy at a cost to biodiversity? impacts of LED lighting on bats. Glob. Chang. Biol. 2012, 18, 2458–2465. [Google Scholar] [CrossRef]
- Polak, T.; Korine, C.; Yair, S.; Holderied, M.W. Differential effects of artificial lighting on flight and foraging behaviour of two sympatric bat species in a desert. J. Zool. 2011, 285, 21–27. [Google Scholar] [CrossRef]
- Stone, E.; Harris, S.; Jones, G. Impacts of artificial lighting on bats: A review of challenges and solutions. Mamm. Biol. 2015, 80, 213–219. [Google Scholar] [CrossRef]
- Seewagen, C.L.; Adams, A.M. Turning to the dark side: LED light at night alters the activity and species composition of a foraging bat assemblage in the Northeastern United States. Ecol. Evol. 2021, 11, 5635–5645. [Google Scholar] [CrossRef]
- Spoelstra, K.; van Grunsven, R.H.A.; Ramakers, J.J.C.; Ferguson, K.B.; Raap, T.; Donners, M.; Veenendaal, E.M.; Visser, M.E. Response of bats to light with different spectra: Light-shy and agile bat presence is affected by white and green, but not red light. Proc. R. Soc. B Biol. Sci. 2017, 284, 20170075. [Google Scholar] [CrossRef]
- Bennett, V.J.; Hale, A.M. Red aviation lights on wind turbines do not increase bat-turbine collisions. Anim. Conserv. 2014, 17, 354–358. [Google Scholar] [CrossRef]
- Weaver, S.P.; Morton, B.P. The secret nightlife of bats: The ultimate flash mob. Presented at the Bat Behavior and Interactions with Wind Turbines Webinar, Online, 14 December 2021; Available online: https://tethys.pnnl.gov/events/awwi-webinar-bat-behavior-interactions-wind-turbines (accessed on 14 December 2021).
- Dechmann, D.K.N.; Safi, K. Studying communication in bats. Cogn. Brain Behav. 2005, 9, 479–496. [Google Scholar]
- Chaverri, G.; Ancillotto, L.; Russo, D. Social communication in bats. Biol. Rev. 2018, 93, 1938–1954. [Google Scholar] [CrossRef] [PubMed]
- Huzzen, B.E.; Hale, A.M.; Bennett, V.J. An effective survey method for studying volant species activity and behavior at tall structures. PeerJ 2020, 2020, e8438. [Google Scholar] [CrossRef] [PubMed]
- Lewis, T. The distribution of flying insects near a low hedgerow. J. Appl. Ecol. 1969, 6, 443–452. [Google Scholar] [CrossRef]
- Pasek, J.E. Influence of wind and windbreaks on local dispersal of insects. Agric. Ecosyst. Environ. 1988, 22–23, 539–554. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guest, E.E.; Stamps, B.F.; Durish, N.D.; Hale, A.M.; Hein, C.D.; Morton, B.P.; Weaver, S.P.; Fritts, S.R. An Updated Review of Hypotheses Regarding Bat Attraction to Wind Turbines. Animals 2022, 12, 343. https://doi.org/10.3390/ani12030343
Guest EE, Stamps BF, Durish ND, Hale AM, Hein CD, Morton BP, Weaver SP, Fritts SR. An Updated Review of Hypotheses Regarding Bat Attraction to Wind Turbines. Animals. 2022; 12(3):343. https://doi.org/10.3390/ani12030343
Chicago/Turabian StyleGuest, Emma E., Brittany F. Stamps, Nevin D. Durish, Amanda M. Hale, Cris D. Hein, Brogan P. Morton, Sara P. Weaver, and Sarah R. Fritts. 2022. "An Updated Review of Hypotheses Regarding Bat Attraction to Wind Turbines" Animals 12, no. 3: 343. https://doi.org/10.3390/ani12030343
APA StyleGuest, E. E., Stamps, B. F., Durish, N. D., Hale, A. M., Hein, C. D., Morton, B. P., Weaver, S. P., & Fritts, S. R. (2022). An Updated Review of Hypotheses Regarding Bat Attraction to Wind Turbines. Animals, 12(3), 343. https://doi.org/10.3390/ani12030343