
Temperature and Diet Acclimation Modify the Acute Thermal Performance of the Largest Extant Amphibian

 , ,
, ,  ,
, 

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Acclimation
2.2. Respiration Rate
2.3. Measurement of Thermal Limits
2.4. Statistical Analyses
3. Results
3.1. Influences of Temperature and Diet Acclimation on Larvae Metabolism
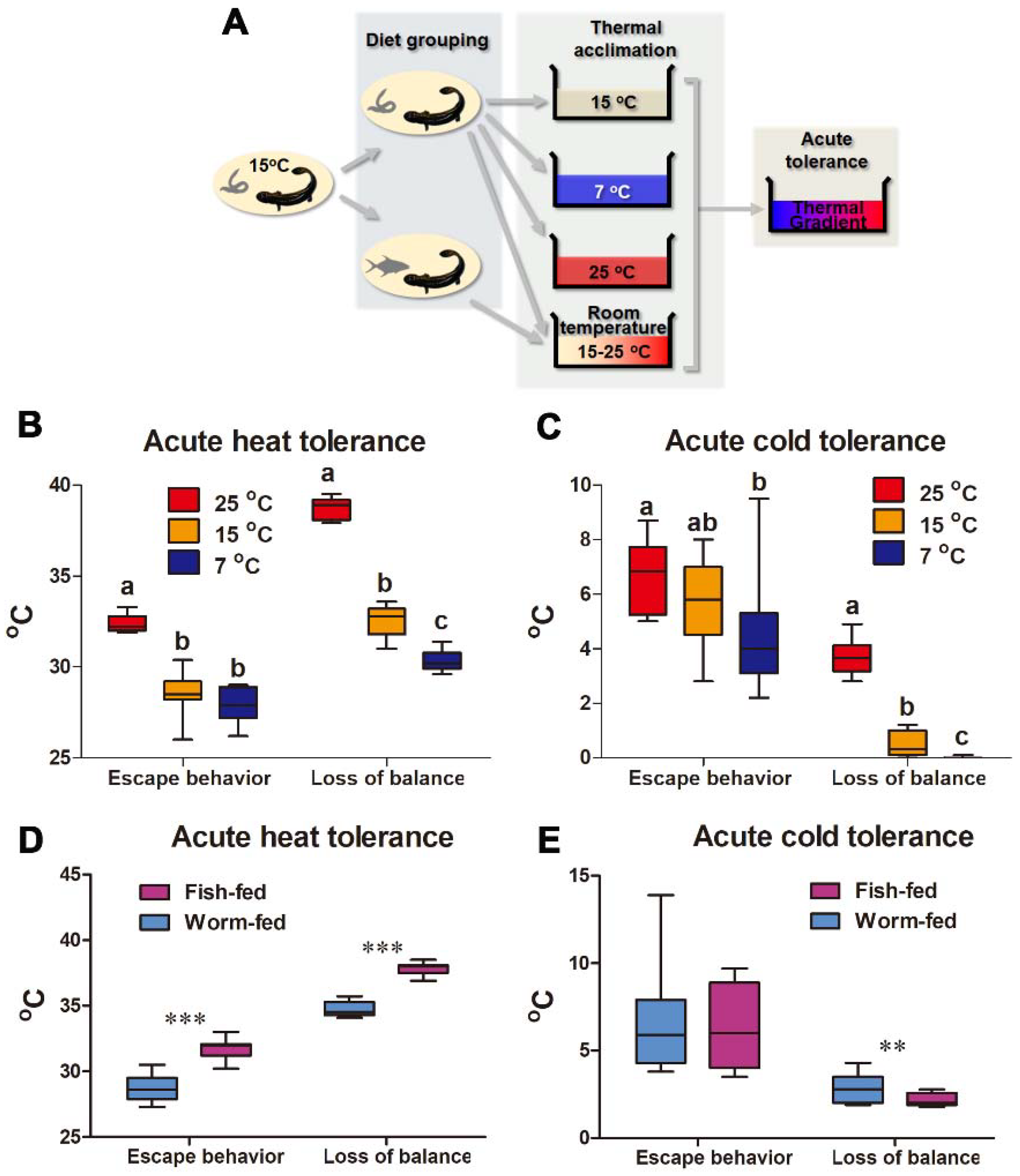
3.2. Influences of Temperature and Diet Acclimation on the Acute Thermal Tolerance Window
4. Discussion
4.1. Temperature-Induced Shift of Thermal Limits
4.2. Diet-Induced Shift of Thermal Limits
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Galloy, V.; Denoël, M. Detrimental effect of temperature increase on the fitness of an amphibian (Lissotriton helveticus). Acta Oecol. 2010, 36, 179–183. [Google Scholar] [CrossRef] [Green Version]
- Rohr, J.R.; Raffel, T.R. Linking global climate and temperature variability to widespread amphibian declines putatively caused by disease. Proc. Natl. Acad. Sci. USA 2010, 107, 8269–8274. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alan Pounds, J.; Bustamante, M.R.; Coloma, L.A.; Consuegra, J.A.; Fogden, M.P.L.; Foster, P.N.; La Marca, E.; Masters, K.L.; Merino-Viteri, A.; Puschendorf, R.; et al. Widespread amphibian extinctions from epidemic disease driven by global warming. Nature 2006, 439, 161–167. [Google Scholar] [CrossRef]
- Dahlke, F.T.; Wohlrab, S.; Butzin, M.; Pörtner, H.O. Thermal bottlenecks in the life cycle define climate vulnerability of fish. Science 2020, 369, 65. [Google Scholar] [CrossRef] [PubMed]
- Stillman, J.H. Acclimation capacity underlies susceptibility to climate change. Science 2003, 301, 65. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutterschmidt, W.I.; Hutchison, V.H. The critical thermal maximum: History and critique. Can. J. Zool. 1997, 75, 1561–1574. [Google Scholar] [CrossRef]
- Calosi, P.; Bilton, D.T.; Spicer, J.I.; Votier, S.C.; Atfield, A. What determines a species’ geographical range? Thermal biology and latitudinal range size relationships in european diving beetles (coleoptera: Dytiscidae). J. Anim. Ecol. 2010, 79, 194–204. [Google Scholar] [CrossRef]
- Kellermann, V.; Overgaard, J.; Hoffmann, A.A.; Fløjgaard, C.; Svenning, J.C.; Loeschcke, V. Upper thermal limits of Drosophila are linked to species distributions and strongly constrained phylogenetically. Proc. Natl. Acad. Sci. USA 2012, 109, 16228–16233. [Google Scholar] [CrossRef] [Green Version]
- Andersen, J.L.; Manenti, T.; Sørensen, J.G.; MacMillan, H.A.; Loeschcke, V.; Overgaard, J. How to assess drosophila cold tolerance: Chill coma temperature and lower lethal temperature are the best predictors of cold distribution limits. Funct. Ecol. 2015, 29, 55–65. [Google Scholar] [CrossRef]
- Eliason, E.J.; Clark, T.D.; Hague, M.J.; Hanson, L.M.; Gallagher, Z.S.; Jeffries, K.M.; Gale, M.K.; Patterson, D.A.; Hinch, S.G.; Farrell, A.P. Differences in thermal tolerance among sockeye salmon populations. Science 2011, 332, 109–112. [Google Scholar] [CrossRef] [Green Version]
- Pinsky, M.L.; Eikeset, A.M.; McCauley, D.J.; Payne, J.L.; Sunday, J.M. Greater vulnerability to warming of marine versus terrestrial ectotherms. Nature 2019, 569, 108–111. [Google Scholar] [CrossRef] [PubMed]
- Sunday, J.; Bennett, J.M.; Calosi, P.; Clusella-Trullas, S.; Gravel, S.; Hargreaves, A.L.; Leiva, F.P.; Verberk, W.C.E.P.; Olalla-Tárraga, M.Á.; Morales-Castilla, I. Thermal tolerance patterns across latitude and elevation. Philos. Trans. R. Soc. London. Ser. B Biol. Sci. 2019, 374, 20190036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pörtner, H.O.; Knust, R. Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 2007, 315, 95–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pörtner, H.O. Climate variations and the physiological basis of temperature dependent biogeography: Systemic to molecular hierarchy of thermal tolerance in animals. Comp. Biochem. Physiol. A 2002, 132, 739–761. [Google Scholar] [CrossRef]
- Martin, B.T.; Dudley, P.N.; Kashef, N.S.; Stafford, D.M.; Reeder, W.J.; Tonina, D.; Del Rio, A.M.; Scott Foott, J.; Danner, E.M. The biophysical basis of thermal tolerance in fish eggs. Proc. Biol. Sci. 2020, 287, 20201550. [Google Scholar] [CrossRef]
- Zhu, W.; Meng, Q.; Zhang, H.; Wang, M.L.; Li, X.; Wang, H.T.; Zhou, G.L.; Miao, L.; Qin, Q.L.; Zhang, J.H. Metabolomics reveals the key role of oxygen metabolism in heat susceptibility of an alpine-dwelling ghost moth, Thitarodes xiaojinensis (lepidoptera: Hepialidae). Insect Sci. 2019, 26, 695–710. [Google Scholar] [CrossRef]
- Javal, M.; Roques, A.; Roux, G.; Laparie, M. Respiration-based monitoring of metabolic rate following cold-exposure in two invasive Anoplophora species depending on acclimation regime. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2018, 216, 20–27. [Google Scholar] [CrossRef]
- Coughlin, D.J.; Shiels, L.P.; Nuthakki, S.; Shuman, J.L. Thermal acclimation to cold alters myosin content and contractile properties of rainbow smelt, Osmerus mordax, red muscle. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2016, 196, 46–53. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Chown, S.L.; Clusella-Trullas, S.; Fox, C. Upper thermal limits in terrestrial ectotherms: How constrained are they? Funct. Ecol. 2013, 27, 934–949. [Google Scholar] [CrossRef]
- Bujan, J.; Roeder, K.A.; Yanoviak, S.P.; Kaspari, M. Seasonal plasticity of thermal tolerance in ants. Ecology 2020, 101, e03051. [Google Scholar] [CrossRef]
- Yu, X.; Yu, K.; Huang, W.; Liang, J.; Qin, Z.; Chen, B.; Yao, Q.; Liao, Z. Thermal acclimation increases heat tolerance of the scleractinian coral Acropora pruinosa. Sci. Total Environ. 2020, 733, 139319. [Google Scholar] [CrossRef] [PubMed]
- Enriquez, T.; Colinet, H. Cold acclimation triggers major transcriptional changes in Drosophila suzukii. BMC Genom. 2019, 20, 413. [Google Scholar] [CrossRef] [PubMed]
- Guderley, H. Functional significance of metabolic responses to thermal acclimation in fish muscle. Am. J. Physiol. Regul. Integr. Comp. Physiol. 1990, 259, R245–R252. [Google Scholar] [CrossRef] [PubMed]
- Seebacher, F.; White, C.R.; Franklin, C.E. Physiological plasticity increases resilience of ectothermic animals to climate change. Nat. Clim. Change 2015, 5, 61–66. [Google Scholar] [CrossRef]
- Rohr, J.R.; Civitello, D.J.; Cohen, J.M.; Roznik, E.A.; Sinervo, B.; Dell, A.I. The complex drivers of thermal acclimation and breadth in ectotherms. Ecol. Lett. 2018, 21, 1425–1439. [Google Scholar] [CrossRef] [Green Version]
- Chevin, L.M.; Lande, R.; Mace, G.M.; Kingsolver, J.G. Adaptation, plasticity, and extinction in a changing environment: Towards a predictive theory. PLoS Biol. 2010, 8, e1000357. [Google Scholar] [CrossRef] [Green Version]
- Verberk, W.C.E.P.; Overgaard, J.; Ern, R.; Bayley, M.; Wang, T.; Boardman, L.; Terblanche, J.S. Does oxygen limit thermal tolerance in arthropods? A critical review of current evidence. Comp. Biochem. Phys. A 2016, 192, 64–78. [Google Scholar] [CrossRef] [Green Version]
- Mitchell, K.A.; Boardman, L.; Clusella-Trullas, S.; Terblanche, J.S. Effects of nutrient and water restriction on thermal tolerance: A test of mechanisms and hypotheses. Compa. Biochem. Phys. A 2017, 212, 15–23. [Google Scholar] [CrossRef]
- Stuart, S.N.; Chanson, J.S.; Cox, N.A.; Young, B.E.; Rodrigues, A.S.; Fischman, D.L.; Waller, R.W. Status and trends of amphibian declines and extinctions worldwide. Science 2004, 306, 1783–1786. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Jiang, W.; Wang, Q.J.; Zhao, H.; Zhang, H.X.; Marcec, R.M.; Willard, S.T.; Kouba, A.J. Reintroduction and post-release survival of a living fossil: The chinese giant salamander. PLoS ONE 2016, 11, e0156715. [Google Scholar] [CrossRef]
- Todd, W.P.; Fang, Y.; WANG, Y.; Theodore, P. A survey for the chinese giant salamander (Andrias davidianus; blanchard, 1871) in the qinghai province. Amphib. Reptile Conse. 2014, 8, 1–6. [Google Scholar]
- Wang, J.; Zhang, H.; Xie, F.; Wei, G.; Jiang, J.P. Genetic bottlenecks of the wild chinese giant salamander in karst caves. Asian Herpetol. Res. 2017, 8, 174–183. [Google Scholar]
- Liang, G.; Geng, B.R.; Zhao, E.M. Andrias davidianus. In The IUCN Red List of Threatened Species; IUCN: Gland, Switzerland, 2004; e.T1272A3375181. [Google Scholar]
- Zhao, T.; Zhang, W.Y.; Zhou, J.; Zhao, C.L.; Liu, X.K.; Liu, Z.D.; Shu, G.S.; Wang, S.S.; Li, C.; Xie, F.; et al. Niche divergence of evolutionarily significant units with implications for repopulation programs of the world’s largest amphibians. Sci. Total Environ. 2020, 738, 140269. [Google Scholar] [CrossRef] [PubMed]
- Jiang, J.P.; Xie, F.; Zang, C.X.; Cai, L.; Li, C.; Wang, B.; Li, J.T.; Wang, J.; Hu, J.H.; Wang, Y.; et al. Assessing the threat status of amphibians in china. Biodivers. Sci. 2016, 24, 588–597. [Google Scholar] [CrossRef]
- Zhang, Z.; Mammola, S.; Liang, Z.; Capinha, C.; Wei, Q.; Wu, Y.; Zhou, J.; Wang, C. Future climate change will severely reduce habitat suitability of the critically endangered chinese giant salamander. Freshwater Biol. 2020, 65, 971–980. [Google Scholar] [CrossRef]
- Hu, Q.; Tian, H.; Xiao, H. Effects of temperature and sex steroids on sex ratio, growth, and growth-related gene expression in the chinese giant salamander andrias davidianus. Aquat. Biol. 2019, 28, 79–90. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Kouba, A.; Wang, Q.J.; Zhao, H.; Jiang, W.; Willard, S.; Zhang, H.X. The effect of water temperature on the growth of captive chinese giant salamanders (Andrias davidianus) reared for reintroduction: A comparison with wild salamander body condition. Herpetologica 2014, 70, 369–377. [Google Scholar] [CrossRef]
- Chen, X.; Shen, J. The effect of water temperature on intake of Andrias davidianus (in chinese). Fish. Sci. 1999, 1, 20–22. [Google Scholar]
- Mu, H.m.; Li, Y.; Yao, J.j.; Ma, S. A review: Current research on biology of chinese giant salamander. Fisheries Sci. 2011, 30, 513–516. [Google Scholar]
- Wang, H.W. The artificial culture of chinese giant salamander. J. Aquacult. 2004, 25, 41–44. [Google Scholar]
- Fu, S.J.; Xie, X.J.; Cao, Z.D. Effect of dietary composition on specific dynamic action in southern catfish Silurus meridionalis chen. Aquac. Res. 2005, 36, 1384–1390. [Google Scholar] [CrossRef]
- Layne, J.R.; Claussen, D.L. Seasonal variation in the thermal acclimation of critical thermal maxima (ctmax) and minima (ctmin) in the salamander eurycea bislineata. J. Therm. Biol. 1982, 7, 29–33. [Google Scholar] [CrossRef]
- Wickham, H. Ggplot2: Elegant Graphics for Data Analysis; Springer Publishing Company, Incorporated: New York, NY, USA, 2009. [Google Scholar]
- Guderley, H. Metabolic responses to low temperature in fish muscle. Biol. Rev. 2004, 79, 409–427. [Google Scholar] [CrossRef] [PubMed]
- Abe, T.; Kitagawa, T.; Makiguchi, Y.; Sato, K. Chum salmon migrating upriver adjust to environmental temperatures through metabolic compensation. J. Exp. Biol. 2019, 222, jeb.186189. [Google Scholar]
- Zhu, W.; Zhang, H.; Li, X.; Meng, Q.; Shu, R.; Wang, M.; Zhou, G.; Wang, H.; Miao, L.; Zhang, J.; et al. Cold adaptation mechanisms in the ghost moth Hepialus xiaojinensis: Metabolic regulation and thermal compensation. J. Insect Physiol. 2016, 85, 76–85. [Google Scholar] [CrossRef] [Green Version]
- Storey, K.B.; Storey, J.M. Metabolic regulation and gene expression during aestivation. In Aestivation: Molecular and Physiological Aspects; Arturo Navas, C., Carvalho, J.E., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 25–45. [Google Scholar]
- Staples, J.F. Metabolic flexibility: Hibernation, torpor, and estivation. Compr. Physiol. 2016, 6, 737–771. [Google Scholar]
- Sandblom, E.; Grans, A.; Axelsson, M.; Seth, H. Temperature acclimation rate of aerobic scope and feeding metabolism in fishes: Implications in a thermally extreme future. Proceedings. Biol. Sci. 2014, 281, 20141490. [Google Scholar] [CrossRef] [Green Version]
- Chang, L.; Wang, B.; Zhang, M.; Liu, J.; Zhao, T.; Zhu, W.; Jiang, J. The effects of corticosterone and background colour on tadpole physiological plasticity. Comp. Biochem. Phys. D 2021, 39, 100872. [Google Scholar] [CrossRef]
- Yan, F.; Lü, J.C.; Zhang, B.L.; Yuan, Z.Y.; Zhao, H.P.; Huang, S.; Wei, G.; Mi, X.; Zou, D.H.; Xu, W.; et al. The chinese giant salamander exemplifies the hidden extinction of cryptic species. Curr. Biol. 2018, 28, R590–R592. [Google Scholar] [CrossRef] [Green Version]
- O’Brien, K.M.; Rix, A.S.; Egginton, S.; Farrell, A.P.; Crockett, E.L.; Schlauch, K.; Woolsey, R.; Hoffman, M.; Merriman, S. Cardiac mitochondrial metabolism may contribute to differences in thermal tolerance of red- and white-blooded antarctic notothenioid fishes. J. Exp. Biol. 2018, 221, jeb177816. [Google Scholar] [CrossRef] [Green Version]
- Takeuchi, K.; Nakano, Y.; Kato, U.; Kaneda, M.; Aizu, M.; Awano, W.; Yonemura, S.; Kiyonaka, S.; Mori, Y.; Yamamoto, D.; et al. Changes in temperature preferences and energy homeostasis in dystroglycan mutants. Science 2009, 323, 1740–1743. [Google Scholar] [CrossRef] [PubMed]
- Semsar-kazerouni, M.; Boerrigter, J.G.J.; Verberk, W.C.E.P. Changes in heat stress tolerance in a freshwater amphipod following starvation: The role of oxygen availability, metabolic rate, heat shock proteins and energy reserves. Comp. Biochem. Phys. A 2020, 245, 110697. [Google Scholar] [CrossRef] [PubMed]
- Bujan, J.; Kaspari, M. Nutrition modifies critical thermal maximum of a dominant canopy ant. J. Insect Physiol. 2017, 102, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Gomez Isaza, D.F.; Cramp, R.L.; Smullen, R.; Glencross, B.D.; Franklin, C.E. Coping with climatic extremes: Dietary fat content decreased the thermal resilience of barramundi (Latescalcarifer). Comp. Biochem. Phys. A 2019, 230, 64–70. [Google Scholar] [CrossRef] [PubMed]
- Turko, A.J.; Nolan, C.B.; Balshine, S.; Scott, G.R.; Pitcher, T.E. Thermal tolerance depends on season, age and body condition in imperilled redside dace clinostomus elongatus. Conserv. Physiol. 2020, 8, coaa062. [Google Scholar] [CrossRef] [PubMed]
- Wolfe, G.R.; Hendrix, D.L.; Salvucci, M.E. A thermoprotective role for sorbitol in the silverleaf whitefly, Bemisia argentifolii. J. Insect Physiol. 1998, 44, 597–603. [Google Scholar] [CrossRef]
- Kumar, N.; Minhas, P.S.; Ambasankar, K.; Krishnani, K.K.; Rana, R.S. Dietary lecithin potentiates thermal tolerance and cellular stress protection of milk fish (Chanos chanos) reared under low dose endosulfan-induced stress. J. Therm. Biol. 2014, 46, 40–46. [Google Scholar] [CrossRef]
- Kumar, N.; Ambasankar, K.; Krishnani, K.K.; Kumar, P.; Akhtar, M.S.; Bhushan, S.; Minhas, P.S. Dietary pyridoxine potentiates thermal tolerance, heat shock protein and protect against cellular stress of milkfish (Chanos chanos) under endosulfan-induced stress. Fish Shellfish Immun. 2016, 55, 407–414. [Google Scholar] [CrossRef]
- Tejpal, C.S.; Sumitha, E.B.; Pal, A.K.; Shivananda Murthy, H.; Sahu, N.P.; Siddaiah, G.M. Effect of dietary supplementation of l-tryptophan on thermal tolerance and oxygen consumption rate in cirrhinus mrigala fingerlings under varied stocking density. J. Therm. Biol. 2014, 41, 59–64. [Google Scholar] [CrossRef]
- Gupta, S.K.; Pal, A.K.; Sahu, N.P.; Dalvi, R.S.; Akhtar, M.S.; Jha, A.K.; Baruah, K. Dietary microbial levan enhances tolerance of labeo rohita (Hamilton) juveniles to thermal stress. Aquaculture 2010, 306, 398–402. [Google Scholar] [CrossRef]
- Koštál, V.; Šimek, P.; Zahradníčková, H.; Cimlová, J.; Štětina, T. Conversion of the chill susceptible fruit fly larva (Drosophila melanogaster) to a freeze tolerant organism. P. Nati. Acad. Sci. USA 2012, 109, 3270–3274. [Google Scholar] [CrossRef] [Green Version]
- Verberk, W.C.E.P.; Sommer, U.; Davidson, R.L.; Viant, M.R. Anaerobic metabolism at thermal extremes: A metabolomic test of the oxygen limitation hypothesis in an aquatic insect. Integr. Comp. Biol. 2013, 53, 609–619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, D.L.; Ma, Q.; Wang, J.; Li, L.Y.; Han, S.L.; Limbu, S.M.; Li, D.L.; Chen, L.Q.; Zhang, M.L.; Du, Z.Y. Fasting enhances cold resistance in fish through stimulating lipid catabolism and autophagy. J. Physiol. 2019, 597, 1585–1603. [Google Scholar] [CrossRef] [PubMed]
- Teets, N.M.; Gantz, J.D.; Kawarasaki, Y. Rapid cold hardening: Ecological relevance, physiological mechanisms and new perspectives. J. Exp. Biol. 2020, 223, jeb203448. [Google Scholar] [CrossRef] [PubMed]
- Shu, G.C.; Liu, P.; Zhao, T.; Li, C.; Hou, Y.M.; Zhao, C.L.; Wang, J.; Shu, X.X.; Chang, J.; Jiang, J.P.; et al. Disordered translocation is hastening local extinction of the chinese giant salamander. Asian Herpetol. Res. 2021, 12, 271–279. [Google Scholar]



{kind=link}
{kind=link}
| Nutrients | Worm Diet (g/100 g) | Fish Diet (g/100 g) |
|---|---|---|
| Water | 90.01 ± 0.3 | 69.61 ± 0.6 |
| Lipids | 1.21 ± 0.1 | 8.66 ± 0.8 |
| Proteins | 5.11 ± 0.2 | 17.70 ± 0.3 |
| Carbohydrates | 1.65 ± 0.2 | 0.39 ± 0.0 |
| Factors | Type III Sum of Square | df | Mean Square | F Value | Sig. |
|---|---|---|---|---|---|
| Acclimation temperature (AT) | 1023.872 | 2 | 511.936 | 10.987 | 0.002 |
| Test temperature (TT) | 12,955.612 | 1 | 12,955.612 | 278.041 | <0.001 |
| Diet | 21.827 | 1 | 21.827 | 0.468 | 0.508 |
| AT × Diet | 199.824 | 2 | 99.912 | 2.144 | 0.164 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, C.-L.; Zhao, T.; Feng, J.-Y.; Chang, L.-M.; Zheng, P.-Y.; Fu, S.-J.; Li, X.-M.; Yue, B.-S.; Jiang, J.-P.; Zhu, W. Temperature and Diet Acclimation Modify the Acute Thermal Performance of the Largest Extant Amphibian. Animals 2022, 12, 531. https://doi.org/10.3390/ani12040531
Zhao C-L, Zhao T, Feng J-Y, Chang L-M, Zheng P-Y, Fu S-J, Li X-M, Yue B-S, Jiang J-P, Zhu W. Temperature and Diet Acclimation Modify the Acute Thermal Performance of the Largest Extant Amphibian. Animals. 2022; 12(4):531. https://doi.org/10.3390/ani12040531
Chicago/Turabian StyleZhao, Chun-Lin, Tian Zhao, Jian-Yi Feng, Li-Ming Chang, Pu-Yang Zheng, Shi-Jian Fu, Xiu-Ming Li, Bi-Song Yue, Jian-Ping Jiang, and Wei Zhu. 2022. "Temperature and Diet Acclimation Modify the Acute Thermal Performance of the Largest Extant Amphibian" Animals 12, no. 4: 531. https://doi.org/10.3390/ani12040531
APA StyleZhao, C. -L., Zhao, T., Feng, J. -Y., Chang, L. -M., Zheng, P. -Y., Fu, S. -J., Li, X. -M., Yue, B. -S., Jiang, J. -P., & Zhu, W. (2022). Temperature and Diet Acclimation Modify the Acute Thermal Performance of the Largest Extant Amphibian. Animals, 12(4), 531. https://doi.org/10.3390/ani12040531