
Morphology and Histology of the Orbital Region and Eye of the Asiatic Black Bear (Ursus thibetanus)—Similarities and Differences within the Caniformia Suborder



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Collection of Specimen and Conservation Status
2.2. Ethical Statement
2.3. Gross Macroscopy Analysis
- Axial eye diameter: from the corneal vertex to the root of the optic nerve;
- Maximum transverse (equatorial) eye diameter;
- Minimum transverse (equatorial) eye diameter;
- Maximum and minimum corneal diameter;
- Corneal axial thickness and corneal peripheral thickness;
- Lens axial length;
- Aqueous chamber depth;
- Vitreous chamber depth;
- Tapetum lucidum length;
- Tapetum lucidum thickness.
2.4. Light Microscopic Studies
3. Results
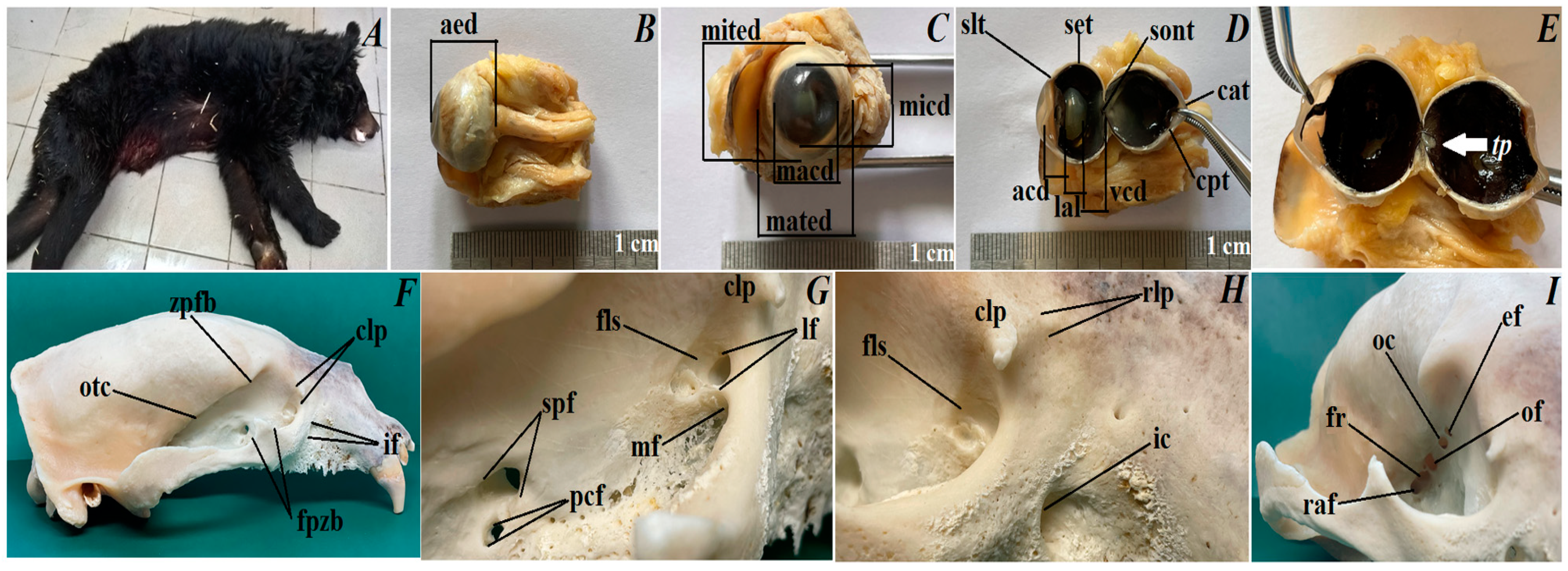
3.1. The Eyeball Morphometry and Orbital Region Description
3.2. Macroscopic Observations of the Eyelids and Orbital Glands
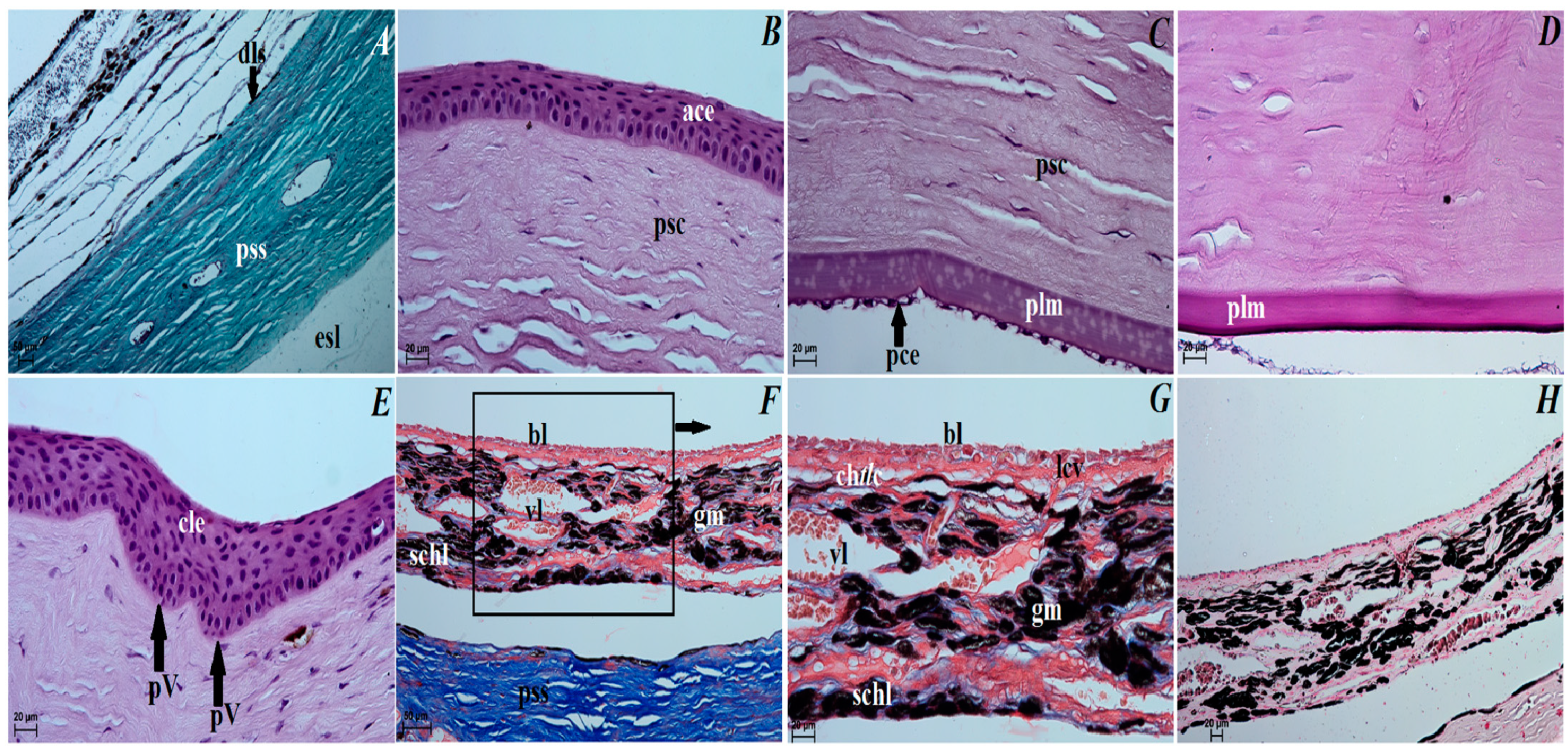
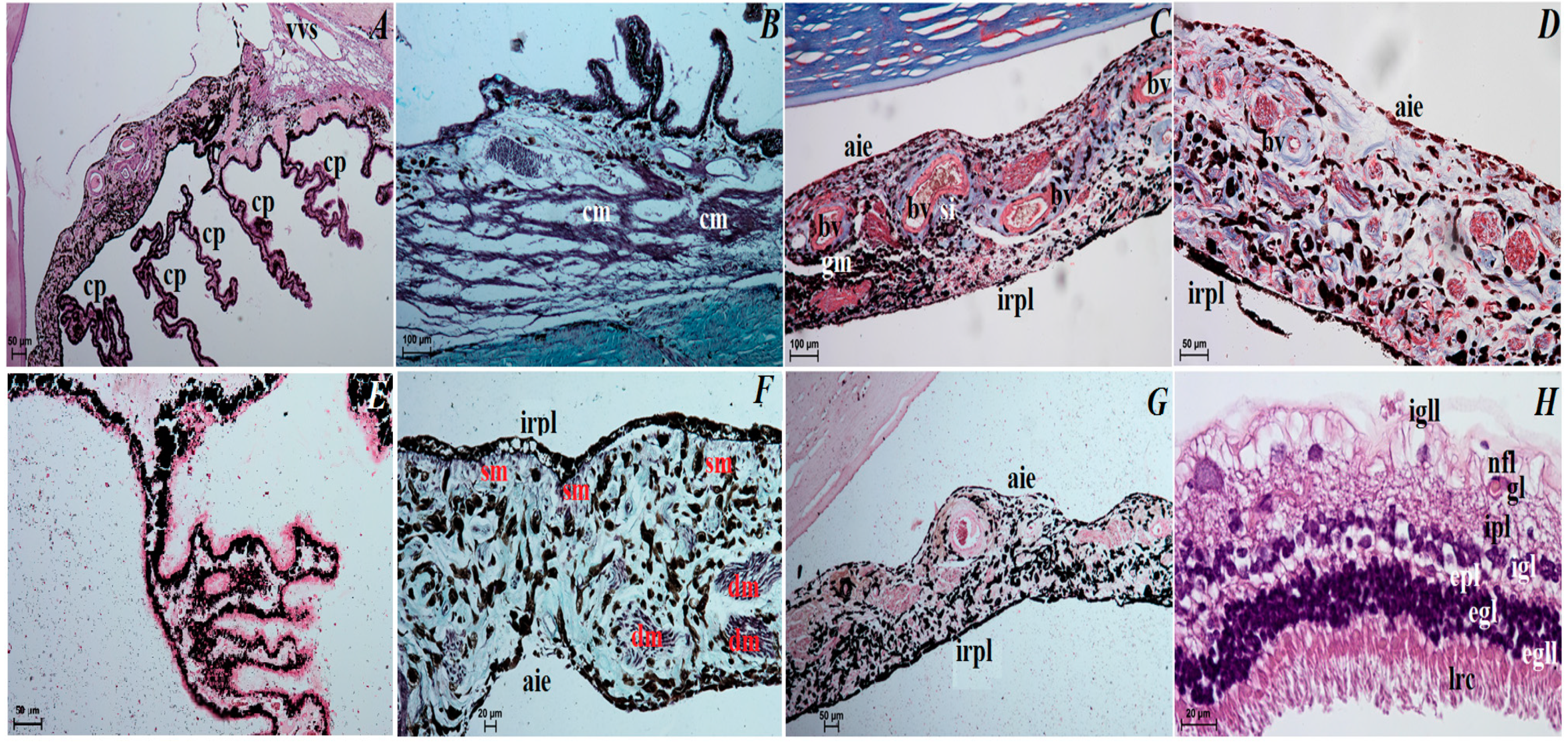
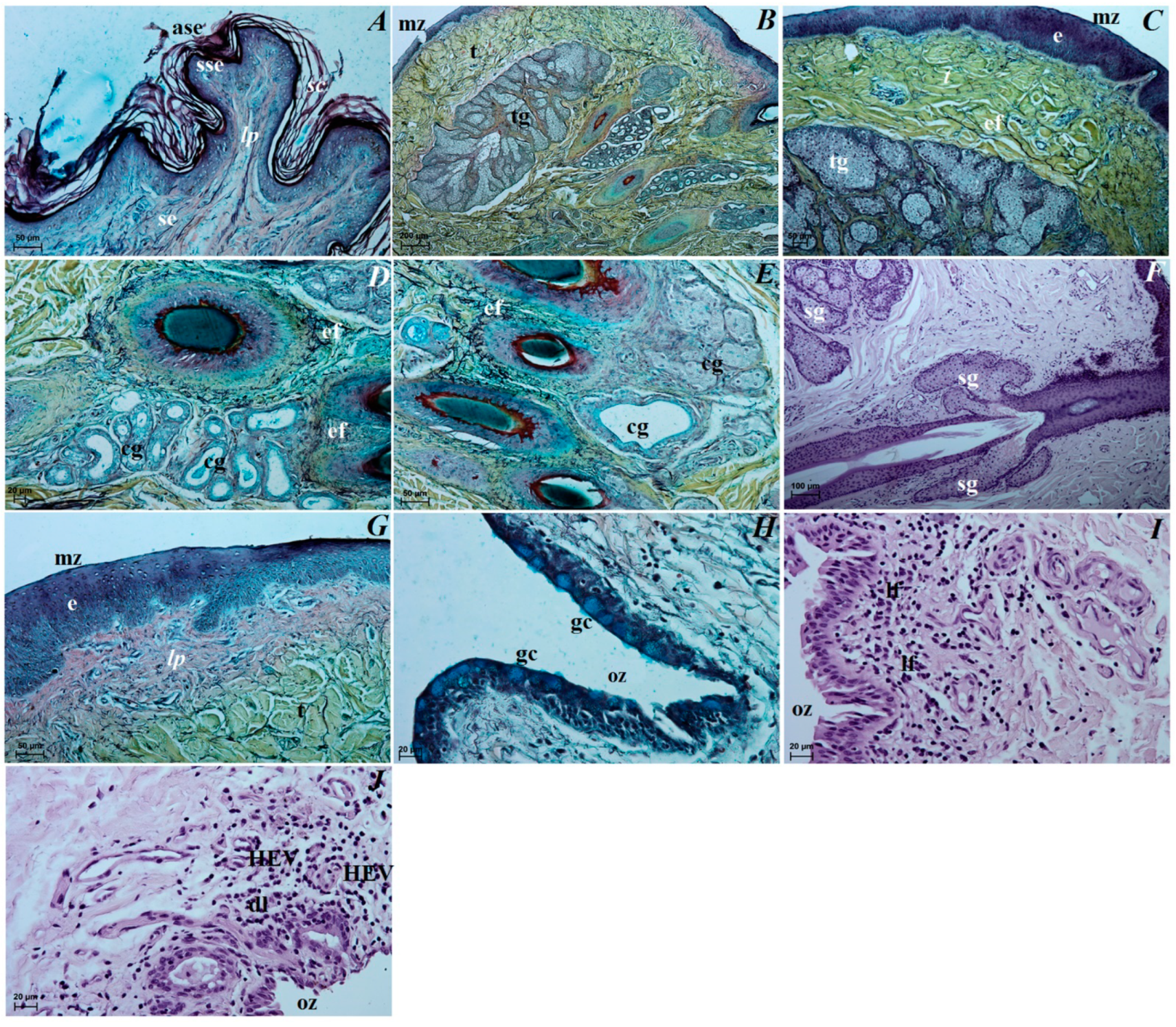
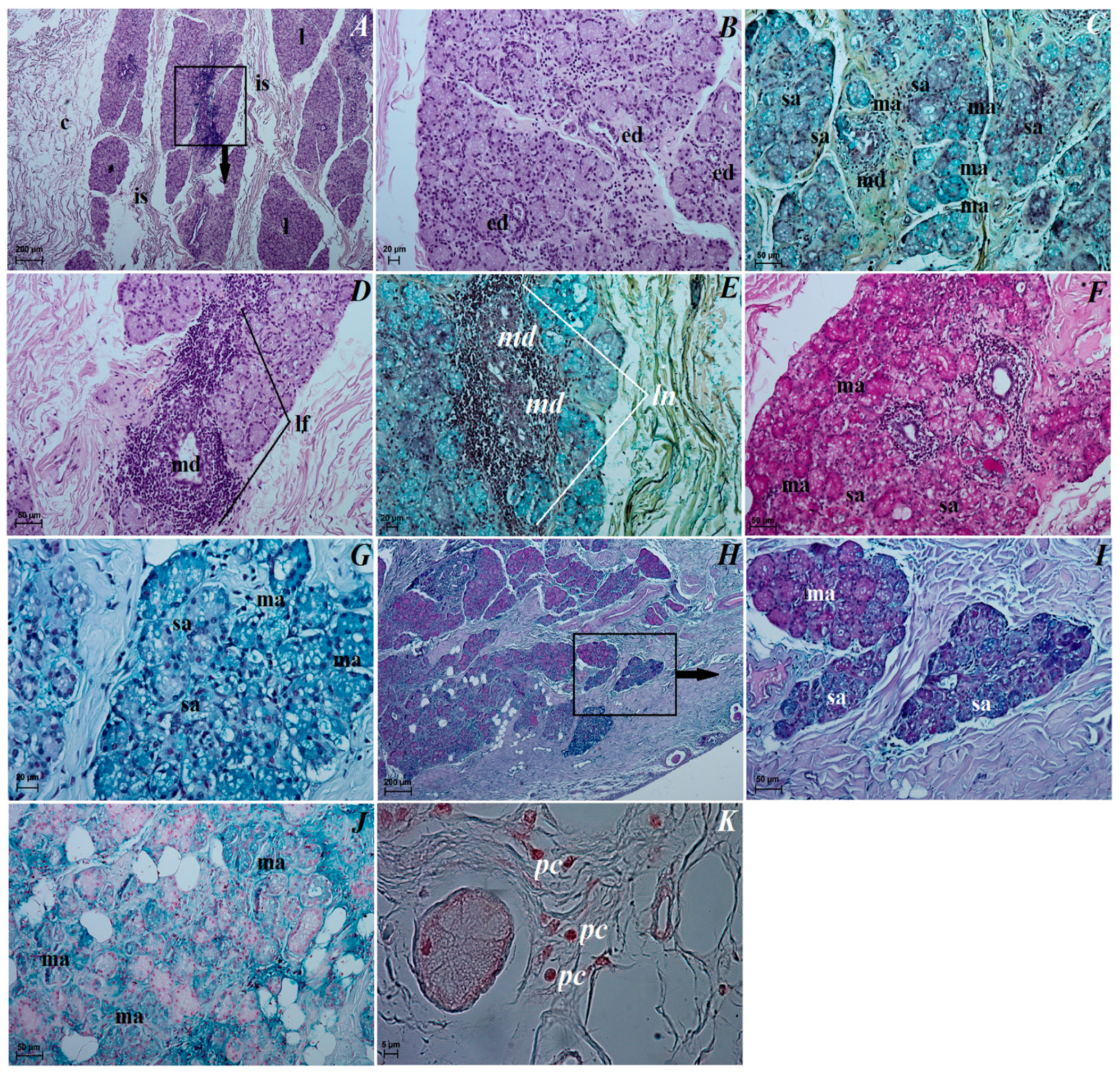
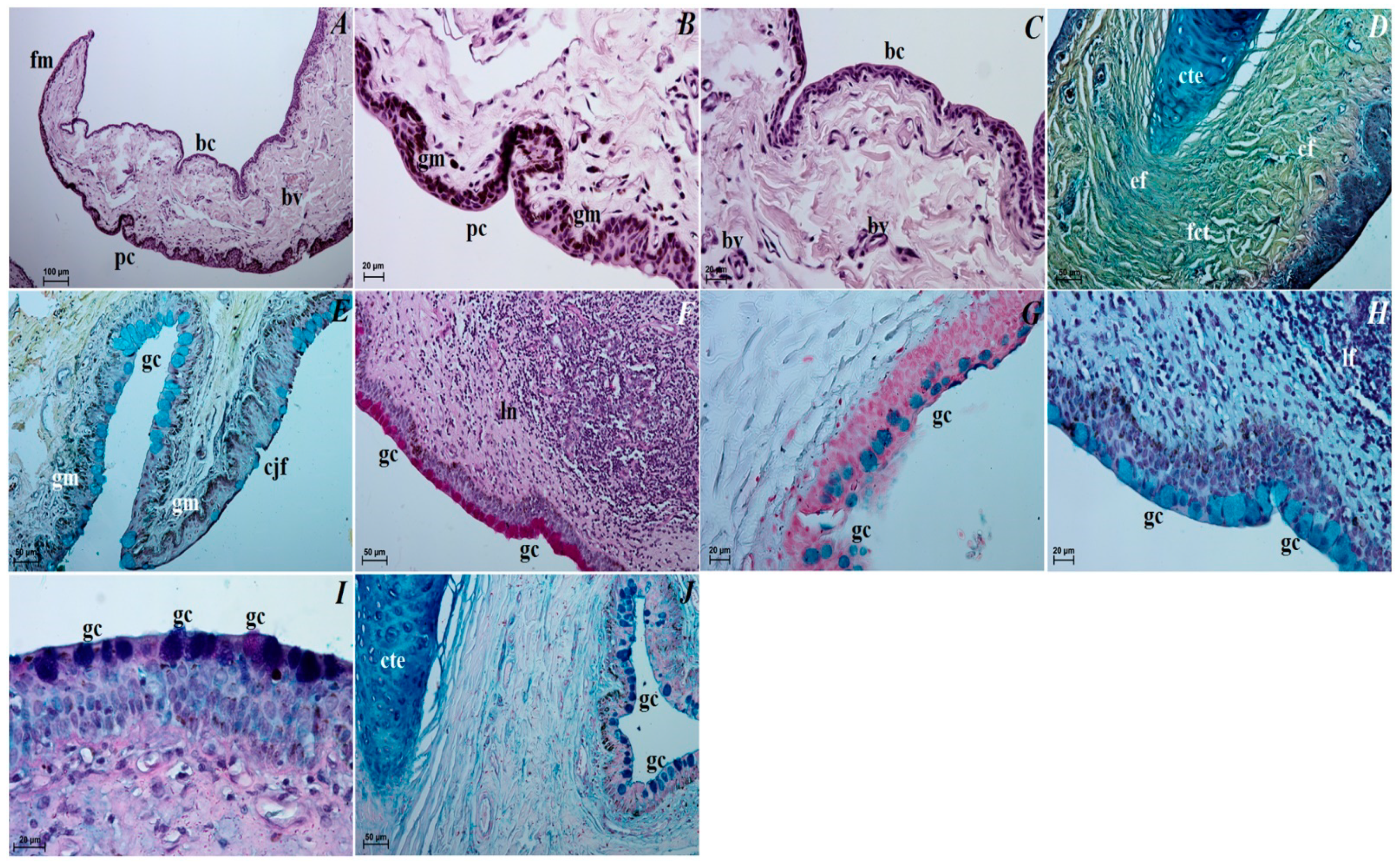
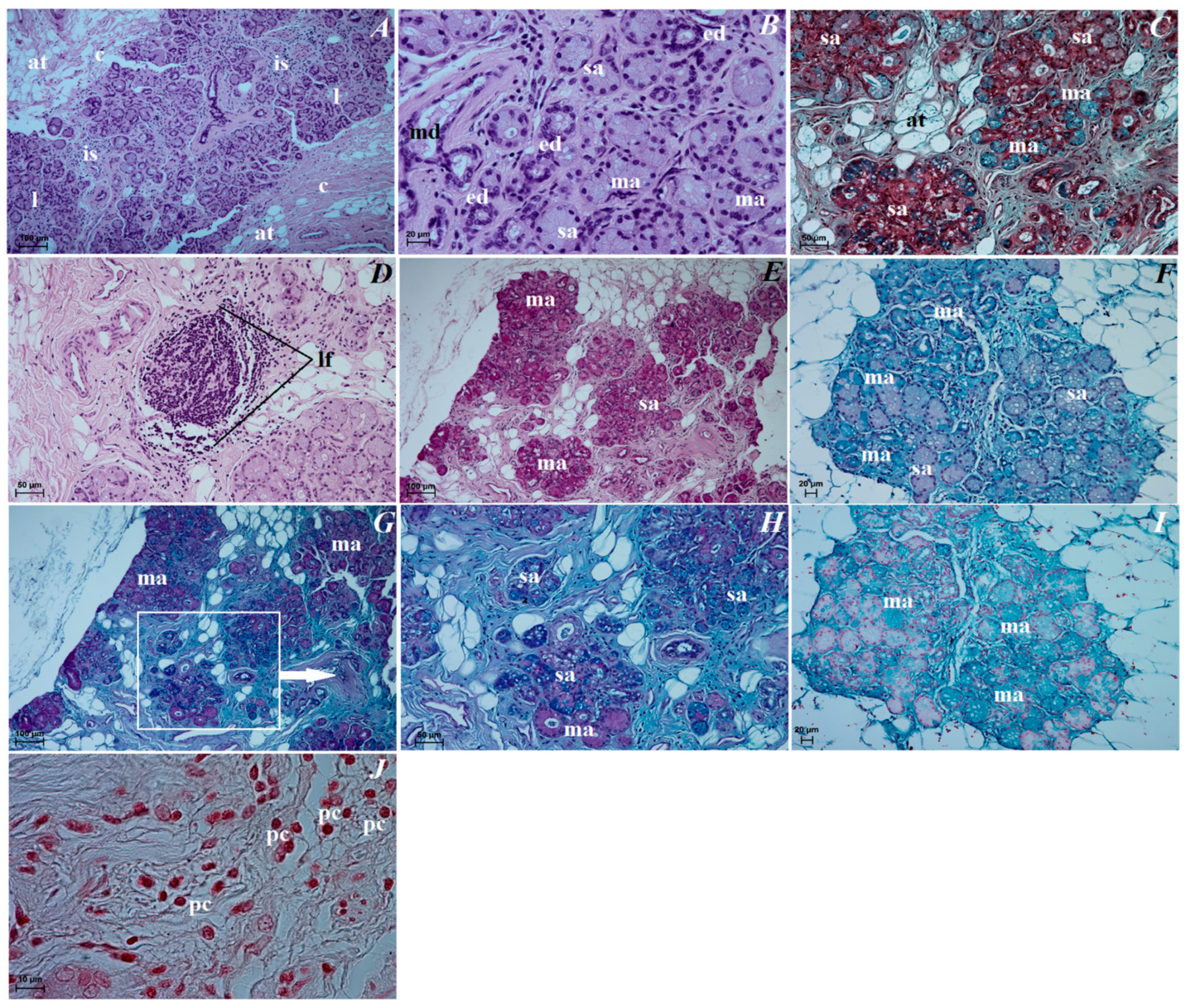
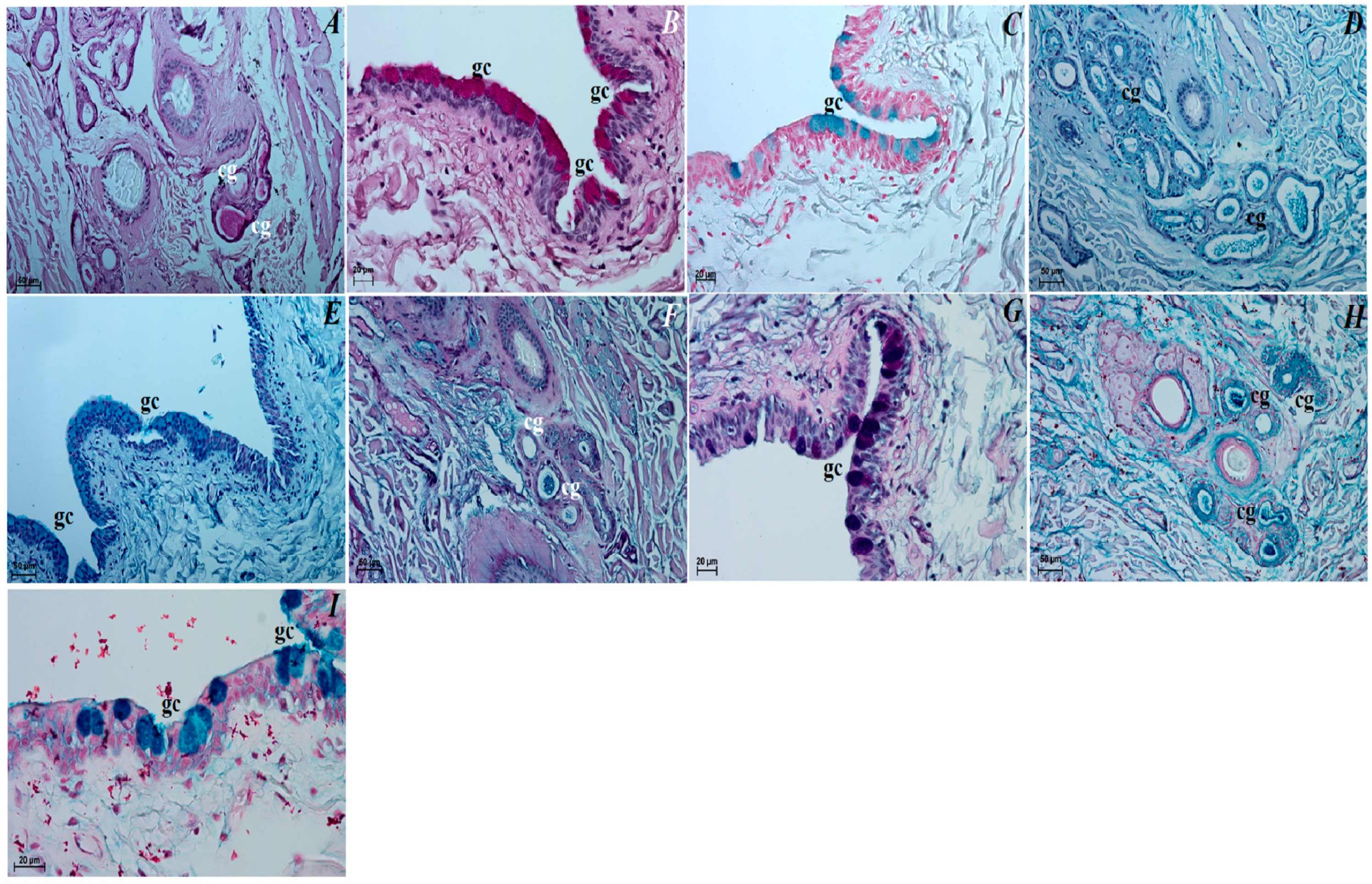
3.3. Histological Observations of the Eye Tunics, Eyelids and Orbital Glands
3.4. Histochemical Observation of the Eyelids and Orbital Glands
4. Discussion
4.1. The Eyeball and Eye Tunics
4.2. The Orbital Region
4.3. The Upper and Lower Eyelids
4.4. The Superficial Gland of the Third Eyelid with the Third Eyelid
4.5. The Lacrimal Gland
5. Conclusions
- -
- The eyeball was a spherical shape, while the periorbita was funnel/conical-shaped and the orbit was an open type;
- -
- The cornea in Asiatic black bear did not include Bowman’s membrane;
- -
- The tapetum lucidum was cellulosum, like other Canidae;
- -
- The individual orbital openings were arranged in a manner typical for Asian black bears (knowledge of their exact location is associated with properly performed local anesthesia, if necessary);
- -
- In the anterior palpebral margin of the upper eyelid, delicate and short eyelashes were observed, while in the lower eyelids they were absent, just like in Canidae;
- -
- The presence of lymphoid follicles, diffuse lymphocytes and high endothelial venules (HEV) within the eyelids as well as in Canidae and from 7–10 conjunctival folds with numerous goblet cells characteristic only to Asiatic black bear;
- -
- The marginal part of the third eyelids was very pigmented and was thick. The third eyelid was T-shaped and present were lymphoid cells that formed the subepithelial conjunctival lymphoid follicle (CALT) characterized to Canidae and Asiatic black bear;
- -
- The superficial gland of the third eyelid was a multilobar vesicular branched complex with seromucous nature and numerous lymphoid cells formed lymph nodes concentrated around the main ducts;
- -
- The lacrimal gland was very small and had a multilobar acinar branched complex structure, producing a muco-serous discharge, a small single lymphatic papule, and numerous plasma cells located in the glandular interstitium.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Cichocki, W.; Ważna, A.; Cichocki, J.; Rajska, E.; Jasiński, A.; Bogdanowicz, W. Polskie Nazewnictwo Ssaków Świata; Muzeum i Instytut Zoologii PAN: Warszawa, Poland, 2015; p. 152. ISBN 978-83-88147-15-9. [Google Scholar]
- Wilson Don, E.; Reeder Dee, A.M. Ursus thibetanus. Mammal Species of the World a Taxonomic and Geographic Reference, 3rd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2005. [Google Scholar]
- Brown, G. The Bear Almanac. A Comprehensive Guide to the Bears of the World, 2nd ed.; The Lyons Press: Guilford, CT, USA, 2009. [Google Scholar]
- Brown, G. The Great Bear Almanac; Lyons & Burford: New York, NY, USA, 1993; ISBN 978-1-55821-210-7. [Google Scholar]
- Stirling, I. Bears: Majestic Creatures of the Wild; Harper Collins: New York, NY, USA, 1993. [Google Scholar]
- Herrero, S.; Peyton, B.; Servheen, C. Bears: Status Survey and Conservation Action Plan. IUCN/SSC Action Plans for the Conservation of Biological Diversity; IUCN: Gland, Switzerland, 1999. [Google Scholar]
- Heptner, V.G.; Naumov, N.P. Mammals of the Soviet Union; Brill, A.E.J., Ed.; Smithsonian Institution Libraries and National Science Foundation: New York, NY, USA, 1992; Volume 2, Pt 1. [Google Scholar]
- Nowak, R. Walker’s Mammals of the World, 5th ed.; Johns Hopkins University Press: Baltimore, MD, USA; London, UK, 1991. [Google Scholar]
- Reid, D.; Jiang, M.; Teng, Q.; Qin, Z.; Hu, J. Ecology of the Asiatic black bear Ursus thibetanus in Sichuan China. Mammalia 1991, 55, 221–237. [Google Scholar] [CrossRef]
- Garshelis, D.L.; Steinmetz, R. “Ursus thibetanus”; IUCN Red List of Threatened Species: Gland, Switzerland, 2020. [Google Scholar]
- Hunter, L. Carnivores of the World. Princeton Field Guides; Princeton University Press: Prniceton, NJ, USA; Oxford, UK, 2011. [Google Scholar]
- Seton, E.T. The Black Bear. In Lives of Game Animals; Charles T. Branford Co.: Boston, MA, USA, 1929; Volume 2. [Google Scholar]
- Skinner, M.P. Bears in the Yellowstone; A. C. McClurg and Co.: Chicago, IL, USA, 1925. [Google Scholar]
- Bacon, E.S. Investigation on Perception and Behavior of the American Black Bear (Ursus americanus). Ph.D. Dissertation, University of Tennessee, Knoxville, TN, USA, 1973. [Google Scholar]
- Kuckuk, E. Tierpsychologische beobachtungen an zwei jungen braun baren. Z. Für Vgl. Physiol. 1937, 24, 14–41. [Google Scholar] [CrossRef]
- Couturier, M.A.J. L’Ours Brun; L’imprimerie Allier: Grenoble, France, 1954. [Google Scholar]
- Shepard, P.; Sanders, B. The Sacred Paw: The Bear in Nature, Myth, and Literature; Viking: New York, NY, USA, 1985. [Google Scholar]
- Kelling, A.S.; Snyder, R.J.; Jackson Marr, M.; Bloomsmith, M.A.; Gardner, W.; Maple, T.L. Color vision in the giant panda (Ailuropoda melanoleuca). Learn. Behav. 2006, 34, 154–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bacon, E.S.; Burghardt, G.M. Learning and color discrimination in the American black bear. In Bears—Their Biology and Management; Pelton, M.R., Lenfer, J.W., Folks, G.E., Jr., Eds.; International Union for the Conservation of Nature & Natural Resources: Morges, Switzerland, 1976; pp. 27–36. [Google Scholar]
- Ducker, G. Colour-vision in mammals. J. Bombay Nat. Hist. Soc. 1964, 61, 572–586. [Google Scholar]
- Jacobs, G.H.; Deegan, I.I.J.F.; Crognale, M.A.; Fenwick, J.A. Photopigments of dogs and foxes and their implications for canid vision. Vis. Neurosci. 1993, 10, 173–180. [Google Scholar] [CrossRef]
- Striling, I. Polar Bears: The Natural History of a Threatened Species; Bloomsbury: London, UK, 2012. [Google Scholar]
- Heyward, J.L.; Reynolds, B.D.; Foster, M.L.; Archibald, K.E.; Stoskopf, M.K.; Mowat, F.M. Retinal cone photoreceptor distribution in the American black bear (Ursus americanus). Anat. Rec. 2020, 304, 662–672. [Google Scholar] [CrossRef] [PubMed]
- Johnson, G.L. Contributions to the comparative anatomy of the mammalian eye. Chiefly based on ophthalmoscopic examination. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1901, 194, 1–82. [Google Scholar]
- Kirk, E.C. Eye morphology in cathemeral lemurids and other mammals. Folia Primatol. 2006, 77, 27–49. [Google Scholar] [CrossRef]
- Kirbas Dogan, G.; Koral Tasci, S.; Dalga, S.; Ilhan Aksu, S. Anatomical and histological studies on the eye of brown bear (Ursus arctos horribilis). Turk. J. Vet. Anim. Sci. 2020, 44, 871–878. [Google Scholar] [CrossRef]
- Peichl, L. Diversity of mammalian photoreceptor properties: Adaptations to habitat and lifestyle? Anat. Rec. Part A Discov. Mol. Cell. Evol. Biol. 2005, 287, 1001–1012. [Google Scholar] [CrossRef]
- Blendea, A.; Gudea, A.; Irimescu, I.; Crisan, M.; Dezdrobitu, C.; Martonos, C.; Bardas, S.; Damian, A. Studies regarding comparative aspects of the splachnocranium in the brown bear (Ursus arctros) and lion (Panthera leo). Bull. Univ. Agric. Sci. Vet. Med. Cluj Napoca Vet. Med. 2013, 70, 7–15. [Google Scholar]
- Nezami, B.; Eagdari, S. Allometric growth pattern of skull on brown bear (Ursus arctos Linnaeus, 1758) of the Alborz Mountain. J. Appl. Biol. Sci. 2014, 8, 52–58. [Google Scholar]
- Mihaylov, R.; Dimitrov, R.; Raichev, E.; Kostov, D.; Stamatova-Yiovcheva, K.; Zlatanova, D.; Bivolarski, B. Morphometrical features of the head skeleton in brown bear (Ursus arctos) in Bulgaria. Bulg. J. Agric. Sci. 2013, 19, 331–337. [Google Scholar]
- Yousefi, M.H. Anatomical study of the Iranian brown bear’s skull (Ursus arctos); A case report. Iran. J. Vet. Med. 2016, 10, 237–244. [Google Scholar]
- Kalita, P.C.; Kalita, H.C.; Sarma, K. Anatomy of the skull of sloth bear (Melursus ursinus). Indian J. Anim. Sci. 2006, 76, 225–227. [Google Scholar]
- Rajashailesha, N.M.; Prasad, R.V.; Sha, A.A.; Jamuna, K.V.; Satyanarayana, M.L.; Ganga Naik, S. Gross anatomy of bony orbit and eyeball of sloth bear (Melursus ursinus). Indian J. Vet. Anat. 2018, 30, 85–87. [Google Scholar]
- Blake, C.N.; Collins, D. Captive Ursids: Results of A Multi-Institutional Survey; Cambridge University Press: Cambridge, UK, 2002; pp. 21–26. [Google Scholar]
- Boedeker, N.C.; Walsh, T.; Murray, S.; Bromberg, N. Medical and surgical management of severe inflammation of the nictitating membrane in a giant panda (Ailuropoda melanoleuca). Vet. Ophthalmol. 2010, 13, 109–115. [Google Scholar] [CrossRef]
- Caruso, K.A.; Koch, S.; Reynolds, B.D.; McCarthy, P.M.G.; Whittaker, C.J. Progressive visual loss and severe retinal degeneration in a captive Kodiak bear (Ursus arctos mittendorfi). Vet. Rec. Case Rep. 2020, 8, e001115. [Google Scholar] [CrossRef]
- Dombrowski, E.; McGregor, G.F.; Bauer, B.S.; Parker, D.; Grahn, B.H. Blindness in a wild American black bear cub (Ursus americanus). Vet. Ophthalmol. 2016, 19, 340–346. [Google Scholar] [CrossRef]
- Hartley, C. Ocular disease in rescued moon bears. Vet. Times 2009, 39, 12–15. [Google Scholar]
- Hartley, C.; Donaldson, D.; Bacon, H.; Officer, K.; Bando, M.; O’Dwyer, J.; Reynard, J.; Leadbeater, W.; Field, N.; Nelson, C.; et al. Ocular findings in Asiatic black bears (Ursus thibetanus), Malayan sun bears (Helarctos malayanus), Eurasian brown bears (Ursus arctos arctos), and a Tibetan brown bear (Ursus arctos pruinosus) rescued from the bile farming industry & wildlife trade in Asia. Vet. Ophthalmol. 2014, 16, E36. [Google Scholar]
- Papadopoulos, E.; Komnenou, A.; Karamanlidis, A.A.; Bezerra-Santos, M.A.; Otranto, D. Zoonotic Thelazia callipaeda eyeworm in brown bears (Ursus arctos): A new host record in Europe. Transbound. Emerg. Dis. 2021, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Kaya, M.; Dorrestein, G.M. Dental and eye diseases of Istanbul bears. Proc. Int. Conf. Asp. Bear Conserv. 1994, 1, 143–148. [Google Scholar]
- Klos, H.-G.; Lang, E.M. Handbook of Zoo Medicine: Diseases and Treatments of Wild Animals in Zoos, Games Parks, Circuses and Private Collections; Van Nostrand Reinhold Company: New York, NY, USA, 1982; pp. 123–130. [Google Scholar]
- Lopez, M.; Talalvera, C.; Rest, J.R.; Taylor, D. Hemangiosarcoma of the conjunctiva of a giant panda. Vet. Rec. 1996, 138, 24. [Google Scholar]
- Mainka, S.A.; Christmas, R. Ocular nodular fasciitis in an Asiatic. J. Vet. Med. Sci. 2020, 82, 740–744. [Google Scholar]
- Maehara, S.; Matsumoto, N.; Takiyama, N.; Itoh, Y.; Kitamura, Y.; Yamashita, K.; Sano, T.; Itami, T.; Oyama, N.; Hayashi, M.; et al. Surgical removal of cataract in an Asiatic black bear (Ursus thibetanus) by phacoemulsification and aspiration black bear (Selenarctos thibetanus). J. Zoo Anim. Med. 1987, 18, 56–158. [Google Scholar]
- McLean, I.W.; Bodman, M.G.; Montali, R.J. Retinal astrocytic hamartomas: Unexpected findings in a giant panda. Arch. Ophthalmol. 2003, 121, 1786–1790. [Google Scholar] [CrossRef] [Green Version]
- Miller, S.N.; Colitz, C.M.H.; Dubielzig, R.R. Anatomy of the California sea lion globe. Vet. Ophthalmol. 2010, 13, 63–71. [Google Scholar] [CrossRef]
- Reynard, J.; Bando, M.; Hartley, C.; Donaldson, D.; Bacon, H.; Robinson, J.; Field, N. Eye pathologies and treatments in bears rescued from the bear bile farming industry in China. In Proceedings of the International Conference on Diseases of Zoo and Wild Animals, Beekse Bergen, The Netherlands, 20–24 May 2013; pp. 71–74. [Google Scholar]
- Sakai, H.; Goto, M.; Komatsu, T. Basal cell adenocarcinoma in the gland of the third eyelid of a brown bear (Ursus arctos). J. Vet. Med. Sci. 2017, 79, 1348–1351. [Google Scholar] [CrossRef] [Green Version]
- Seruca, C.; Lobo, T.; Leiva, M.; Costa, D.; Bernardino, R.; Pena, T. Use of modified semicircular flap combined with advancement for medial lower eyelid reconstruction in a brown bear (Ursus arctos). Annu. Sci. Meet. Eur. Coll. Vet. Ophthalmol. 2014, 17, 463–5216. [Google Scholar]
- Stades, F.C.; Dorrestein, G.M.; Boeve, M.H.; van de Sandt, R.R.D.M. Eye lesions in Turkish dancing bears. Vet. Q. 1995, 17, 45–46. [Google Scholar] [CrossRef]
- Wolff, P. Selected medical aspects of the spectacled bear (Tremarctos ornatus). In Proceedings of the First International Symposium on the Spectacled Bear, Chicago, IL, USA, 14–15 October 1988; pp. 313–317. [Google Scholar]
- IUCN Red List of Threatened Species. 2020. Available online: https://www.iucnredlist.org (accessed on 10 February 2022).
- Nomina Anatomica Veterinaria, 6th ed.; Editoral Committee: Hanover, Germany; Ghent, Belgium; Columbia, MO, USA; Rio de Janeiro, Brazil, 2017.
- Nomina Histologica Veterinaria; The International Committee on Veterinary Histological Nomenclature (ICVHN) to the World Association Veterinary Anatomists: Knoxville, TN, USA, 2017.
- Hermanson, J.W.; de Lahunta, A.; Evans, H.E. Miller and Evans’ Anatomy of The Dog, 5th ed.; Elsevier Inc.: Amsterdam, The Netherlands, 2020. [Google Scholar]
- Nickel, R.; Schummer, A.; Seiferle, E. Lehrbuch der Anatomie der Haustiere; Verlag Paul Parey: Berlin/Hamburg, Germany, 2014; Volume I. [Google Scholar]
- Bishop, J.A.; Nelson, A.M.; Merz, W.G.; Askin, F.B.; Riedel, S. Evaluation of the detection of melanin by the Fontana-Masson silver stain in tissue with a wide range of organisms including Cryptococcus. Hum. Pathol. 2012, 43, 898–903. [Google Scholar] [CrossRef] [PubMed]
- Burck, N.C. Technika Histologiczna; PZWL: Warszawa, Poland, 1975. [Google Scholar]
- Movat, H.Z. Demonstration of all connective tissue elements in a single section. AMA Arch. Pathol. 1955, 60, 289. [Google Scholar]
- Spicer, S.C.; Henson, J.G. Methods for localizing mucosubstances in epithelial and connective tissue. In Series on Methods and Achievements in Experimental Pathology; Bajusz, E., Jamin, F., Eds.; Karger Press: Basal, Switzerland, 1967; Volume 2, pp. 78–112. [Google Scholar]
- Bancroft, J.D.; Gamble, M. Theory and Practice of Histological Techniques, 6th ed.; Churchill Livingstone Elsevier: Oxford, UK, 2008; pp. 173–174. [Google Scholar]
- Carson, F. Histotechnology A Self-Instructional Text, 1st ed.; ASCP: Chicago, IL, USA, 1990; pp. 126–127. [Google Scholar]
- Munakata, H.; Isemura, M.; Yosizawa, Z. An application of the high-iron diamine staining for detection of sulfated glycoproteins (glycopeptides) in electrophoresis on cellulose acetate membrane. Tohoku J. Exp. Med. 1985, 145, 251–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sheehan, D.C.; Hrapchak, B.B. Theory and Practice Histotechnology, 2nd ed.; CV Mosby: St. Louis, MO, USA, 1980; Volume 52, pp. 164–167. [Google Scholar]
- Heard-Booth, A.N.; Kirk, E.C. The influence of maximum running speed on eye size: A test of Leuckart’s law in mammals. Anat. Rec. 2012, 295, 1053–1062. [Google Scholar] [CrossRef]
- Lantyer-Araujo, N.L.; Nascimento Silva, D.; Estrela-Lima, A.; Muramoto, C.; de Azevedo Libório, F.; da Silva, E.A.; Pontes Oriá, A. Anatomical, histological and computed tomography comparision of the eye and adnexa of crab-eating fox (Cerdocyon thous) to domestic dogs. PLoS ONE 2019, 14, e0224245. [Google Scholar] [CrossRef] [PubMed]
- Paszta, W.; Klećkowska-Nawrot, J.E.; Goździewska-Harłajczuk, K. Anatomical and morphometric evaluation of the orbit, eye tunics, eyelids and orbital glands of the captive females of the South African painted dog (Lycaon pictus pictus Temminck, 1820) (Caniformia: Canidae). PLoS ONE 2021, 16, e0249368. [Google Scholar] [CrossRef]
- Kemp, A.D.; Kirk, E.C. Eye size and visual acuity influence vestibular anatomy in mammals. Anat. Rec. 2014, 297, 781–790. [Google Scholar] [CrossRef]
- Murphy, C.; Bellhorn, R.W.; Williams, T.; Burns, M.S.; Schaeffel, F.; Howland, H.C. Refractive state, ocular anatomy, and accommodative range of the sea otter (Enhydra lutris). Vis. Res. 1990, 30, 23–32. [Google Scholar] [CrossRef]
- Iriarte-Díaz, J. Differential scaling of locomotor performance in small and large terrestrial mammals. J. Exp. Biol. 2002, 205, 2897–2908. [Google Scholar] [CrossRef]
- Hughes, A. The topography of vision in mammals of contrasting life style: Comparative optics and retinal organization. In Handbook of Sensory Physiology VII/5: The Visual System in Vertebrates; Crescitell, F., Ed.; Springer: Berlin, Germany, 1977; pp. 613–756. [Google Scholar]
- Brooke, M.L.; Hanley, S.; Laughlin, S.B. The scaling of eye size with body mass in birds. Proc. R. Soc. Lond. B 1999, 266, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Hall, M.I.; Heesy, C.P. Eye size, flight speed and Leuckart’s Law in birds. J. Zool. 2011, 283, 291–297. [Google Scholar] [CrossRef]
- Davis, R.W. Marine Mammals. In Sensory Systems, Adaptations for an Aquatic Life; Springer: Berlin/Heidelberg, Germany, 2019; pp. 177–217. [Google Scholar]
- Kastelein, R.A.; Zwypfenning, R.C.V.J.; Spekreijse, H.; Dubbeldam, J.L.; Born, E.W. The Anatomy of the walrus head (Odobenus rosmarus). Part 3: The eyes and their function in Walrus ecology. Aquat. Mamm. 1993, 19, 61–92. [Google Scholar]
- Piggins, D.J. Refraction of the harp seal, Phagophilus groenlandicus (Erxleben 1777). Nature 1970, 227, 78–79. [Google Scholar] [CrossRef] [PubMed]
- Sivak, J.G. Accommodation in vertebrates: A contemporarysurvey. Curr. Top. Eye Res. 1980, 3, 281–330. [Google Scholar] [PubMed]
- Hanke, F.D.; Dehnhardt, G.; Chaeffel, F.; Hanke, W. Corneal topography, refractive state, and accommodationin harbor seals (Phoca vitulina). Vis. Res. 2006, 46, 837–847. [Google Scholar] [CrossRef] [Green Version]
- Jamieson, G.S.; Fisher, H.D. The retina of the harbor seal Phoca vitulina. Can. J. Zool. 1971, 49, 19–23. [Google Scholar] [CrossRef]
- Jamieson, G.S.; Fisher, H.D. The pinniped eye: A review. In Functional Anatomy of Marine Mammals I; Harrison, R.J., Ed.; Academic Press: London, UK; New York, NY, USA, 1972; pp. 255–257. [Google Scholar]
- Walls, G.L. The Vertebrate Eye and Its Adaptive Radiation; Hafner Press: New York, NY, USA, 1963. [Google Scholar]
- Mass, A.M.; Supin, A.Y. Adaptive features of aquatic mammals’ eye. Anat. Rec. 2007, 290, 701–715. [Google Scholar] [CrossRef]
- Merindano, M.D.; Costa, J.; Canals, M.; Potau, J.M.; Ruano, D. A comparative study of Bowman’s layer in some mammals: Relationships with other constituent corneal structures. Eur. J. Anat. 2002, 6, 133–139. [Google Scholar]
- Nautscher, N.; Baurer, A.; Steffl, M.; Amselgruber, W.M. Comparative morphological evaluation of domestic animal cornea. Vet. Ophthalmol. 2016, 19, 297–304. [Google Scholar] [CrossRef]
- Merindano, M.D.; Canals, M.; Potau, J.M.; Costa, J.; Ruano, D. Morphomterical features of the corneal epithelium in mammals. Anat. Histol. Embryol. 1998, 27, 105–110. [Google Scholar] [CrossRef] [PubMed]
- Renzo, R. Parâmetros oftálmicos em cachorro-do-mato (Cerdocyon thous, Linnaeus, 1766). Ph.D. Thesis, Universidade Estadual Paulista–UNESP Faculdade De Ciências Agrárias E Veterinárias Câmpus De Jaboticabal, São Paulo, Brazil, 2015. [Google Scholar]
- Gwin, R.M.; Lerner, I.; Warren, J.K.; Gum, G. Decrease in canine endothelial cell density and increase in corneal thickness with age. Investig. Ophthalmol. Vis. Sci. 1982, 22, 267–271. [Google Scholar]
- Mass, A.M.; Supin, A.Y. Eye Optics in Semiaquatic Mammals for Aerial and Aquatic Vision. Brain Behav. Evol. 2018, 92, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Gaiddon, J.; Rosolen, S.G.; Steru, L.; Cook, S.C.; Peiffer, R., Jr. Use of biometry and keratometry for determining optimal power for intraocular lens implants in dogs. Am. J. Vet. Res. 1991, 52, 781–783. [Google Scholar] [PubMed]
- Miller, S.; Whelan, N.; Hope, K.; Nogueira Marmolejo, M.G.; Knightly, F.; Sutherland-Smith, M.; Rivera, S. Survey of clinical ophthalmic disease in the giant panda (Ailuropoda Melanoleuca) among North American Zoological Institutions. J. Zoo Wildl. Med. 2019, 50, 837–844. [Google Scholar] [CrossRef]
- Welsch, U.; Ramdohr, S.; Riedelsheimer, B.; Hebel, R.; Eisert, R.; Plötz, J. Microscopic anatomy of the eye of the deep-diving Antarctic Weddell seal (Leptonychotes weddellii). J. Morphol. 2001, 248, 165–174. [Google Scholar] [CrossRef] [PubMed]
- Sanchez, R.F.; Daniels, J.T. Mini-Review: Limbal Stem Cells Deficiency in Companion Animals: Time to Give Something Back? Curr. Eye Res. 2016, 41, 425–432. [Google Scholar] [CrossRef] [Green Version]
- Dua, H.S. The conjunctiva in corneal epithelial wound healing. Br. J. Ophthalmol. 1998, 82, 1407–1411. [Google Scholar] [CrossRef]
- Dua, H.S.; Saini, J.S.; Azuara-Blanco, A.; Gupta, P. Limbal stem cell deficiency: Concept, aetiology, clinical presentation, diagnosis and management. Indian J. Ophthalmol. 2000, 48, 83–92. [Google Scholar]
- Malmström, T.; Kröger, R.H.H. Pupil shapes and lens optics in the eyes of terrestrial vertebrates. J. Exp. Biol. 2006, 209, 18–25. [Google Scholar] [CrossRef] [Green Version]
- Carvalho, C.M.; Rodarte-Almeida, A.C.V.; Beanes, A.S.; Machado, M.T.S.; Galera, P.D. Ophthalmic contribution to assessing eyes of the two neotropical canids: Cerdocyon thous and Chrysocyon brachyurus. Vet. Ophthalmol. 2020, 23, 460–471. [Google Scholar] [CrossRef] [PubMed]
- Ninomiya, H.; Inomata, T.; Kanemaki, N. Microvasculature of the retina, ciliary processes and choroid in the North American raccon (Procyon lotor) Eye; A scanning electron microscopic study of corrosion casts. J. Vet. Med. Sci. 2005, 67, 547–554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKay Strobel, S.; Moore, B.A.; Freeman, K.S.; Murray, M.J.; Reichmuth, C. Adaptations for amphibious vision in sea otters (Enhydra lutris): Structural and functional observations. J. Comp. Physiol. A 2020, 206, 767–782. [Google Scholar] [CrossRef] [PubMed]
- Hu, H. Research on the Giant Panda; Shanghai Scientific and Technological Education Publishing House: Shanghai, China, 2001. [Google Scholar]
- Schaller, G.; Hu, J.; Pan, W.; Zhu, J. The Giant Pandas of Wolong; University of Chicago Press: Chicago, IL, USA, 1985. [Google Scholar]
- Banks, M.S.; Sprague, W.W.; Schmoll, J.; Parnell, J.A.Q.; Love, G.D. Why do animal eyes have pupils of different shapes? Sci. Adv. 2015, 1, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Sivak, J.G.; Howland, H.C.; West, J.; Weerheim, J. The eye of the hooded seal Cystophora cristata, in air and water. J. Comp. Physiol. A 1989, 165, 771–777. [Google Scholar] [CrossRef]
- West, J.A.; Sivak, J.G.; Murphy, C.J.; Kovacs, K.M. A comparative study of the anatomy of the iris and ciliary body in aquatic mammals. Can. J. Zool. 2011, 69, 2594–2607. [Google Scholar] [CrossRef]
- Murphy, C.J.; Samuelson, D.A.; Pollock, R.V.H. The Eye. Miller’s Anatomy of the Dog, 4th ed.; Evans, H.E., De Lahunta, A., Eds.; Elsevier Saunders: Amsterdam, The Netherlands, 2012; pp. 746–785. [Google Scholar]
- Braekevelt, C.R. Fine structure of the tapetum cellulosum of the Grey seal (Halichoerus grypus). Acta Anat. 1986, 127, 81–87. [Google Scholar] [CrossRef]
- Chijiiwa, T.; Ishibashi, T.; Inomata, H. Histological study of choroidal melanocytes in animals with tapetum lucidum cellulosum. Graefe’s Arch. Clin. Exp. Ophthalmol. 1990, 228, 161–168. [Google Scholar] [CrossRef]
- Donisa, A.; Muste, A.; Beteg, F.; Briciu, R. Morphological aspect of tapetum lucidum at some domestic animals. Bull. UASVM Vet. Med. 2008, 65, 166–170. [Google Scholar]
- Hogg, C.; Neveu, M.; Folkow, L.; Stokkan, K.-A.; Kam, J.H.; Douglas, R.H.; Jeffery, G. The eyes of the deep diving hooded seal (Cystophora cristata) enhance sensitivity to ultraviolet light. Biol. Open 2015, 4, 812–818. [Google Scholar] [CrossRef] [Green Version]
- Lesiuk, T.P.; Braekevelt, C.R. Fine structure of the canine tapetum lucidum. J. Anat. 1983, 136, 157–164. [Google Scholar] [PubMed]
- Ollivier, F.J.; Samuelson, D.A.; Brooks, D.E.; Lewis, P.A.; Kallberg, M.E.; Komáromy, A.M. Comparative morphology of the tapetum lucidum (among selected species). Vet. Ophthalmol. 2004, 7, 11–22. [Google Scholar] [CrossRef] [PubMed]
- Wen, G.Y.; Sturman, J.A.; Shek, J.W. A comparative study of the tapetum, retina and skull of the ferret, dog and cat. Lab. Anim. Sci. 1985, 35, 200–210. [Google Scholar] [PubMed]
- Yamaue, Y.; Hosaka, Y.Z.; Uehara, M. Macroscopic and Histological vartiations in the cellular tapetum in dogs. Anatomy 2014, 76, 1099–1103. [Google Scholar]
- Ahnelt, P.K.; Kolb, H. The mammalian photoreceptor mosaic-adaptive design. Prog. Retin Eye Res. 2000, 19, 711–777. [Google Scholar] [CrossRef]
- Malkemper, E.P.; Peichl, L. Retinal photoreceptor and ganglion cell types and topographies in the red fox (Vulpes vulpes) and Arctic fox (Vulpes lagopus). J. Comp. Neurol. 2018, 526, 2078–2098. [Google Scholar] [CrossRef]
- Mass, A.M. Locatization of the high-resolution area in the ganglion cell layer of the Baikal seal Pusa sibirica Gm. 1788. Dokl. Biol. Sci. 2016, 467, 51–54. [Google Scholar] [CrossRef]
- Mass, A.M.; Supin, A.Y. Retinal topography of the harp seal Pagophilus groenlandicus. Brain Behav. Evol. 2003, 62, 212–222. [Google Scholar] [CrossRef]
- Mass, A.M.; Supin, A.Y. Retinal ganglion cell layer of the Caspian seal Pusa caspica: Topography and localization of the high-resolution area. Brain Behav. Evol. 2010, 76, 144–153. [Google Scholar] [CrossRef]
- Mass, A.M.; Supin, A.Y. Retinal ganglion cell topography and retinal resolution in the Baikal seal (Pusa sibirica). Brain Behav. Evol. 2016, 88, 59–67. [Google Scholar] [CrossRef]
- Nagy, A.R.; Ronald, K. The harp seal, Pagophilus groenlandicus (Erxleben, 1777). VI. Structure of retina. Can. J. Zool. 1970, 48, 367–370. [Google Scholar] [CrossRef] [PubMed]
- Landau, D.; Dawson, W.W. The histology of retinas from the pinnipedia. Vis. Res. 1970, 10, 691–702. [Google Scholar] [CrossRef]
- Nagy, A.R.; Ronald, K. A light and electronmicroscopic study of the structure of the retina of the harp seal, Pagophilus groenlandicus (Erxleben, 1777). Rapp. Proces-Verbaux Réun. Cons. Int. Explor. Mer. 1975, 169, 92–96. [Google Scholar]
- Peichl, L.; Behrmann, G.; Kroger, R.H. For whales and seals the ocean is not blue: A visual pigment loss in marine mammals. Eur. J. Neurosci. 2001, 13, 1520–1528. [Google Scholar] [CrossRef] [Green Version]
- FitzGibbon, T.; Wingate, R.J.; Thompson, I.D. Soma and axon diameter distributions and central projections of ferret retinal ganglion cells. Vis. Neurosci. 1996, 13, 773–786. [Google Scholar] [CrossRef]
- Mass, A.M.; Supin, A.Y. Ganglion cells density and retinal resolution in the sea otter, Enhydra lutris. Brain Behav. Evol. 2000, 55, 111–119. [Google Scholar] [CrossRef]
- Jacobs, G.H.; Deegan, J.F., II. Cone photopigments in nocturnal and diurnal procyonids. J. Comp. Physiol. A 1992, 171, 351–358. [Google Scholar] [CrossRef]
- Zuoliang, F. Comparative anatomical studies on the skeleton of artic foxes. J. Econ. Anim. 2004, 8, 80–84. [Google Scholar]
- Karan, M.; Timurkaan, S.; Ozdemir, D.; Unsaldi, E. Comparative macroanatomical study of the neurocranium in some Carnivora. Anat. Histol. Embryol. 2006, 35, 53–56. [Google Scholar] [CrossRef]
- Yilmaz, S.; Dinc, G.; Toprak, B. Macro-anatomical investigations on skeletons of otter (Lutra lutra). III. Skeleton axiale. Vet. Arh. 2000, 70, 191–198. [Google Scholar]
- Hidaka, S.; Matsumoto, M.; Hiji, H.; Ohsako, S.; Nishinakagawa, H. Morphology and morphometry of skulls of Raccoon dogs Nyctereutes procyonoides and Badgers Meles meles. J. Vet. Med. Sci. 1998, 60, 161–167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debey, L.B. Osteological correlates and phylogenetic analysis of deep diving in living and extinct pinnipeds: What good are big eyes? Mar. Mammal Sci. 2013, 29, 48–83. [Google Scholar] [CrossRef]
- Kastelein, R.A.; Gerrits, N.M. The anatomy of the walrus head (Odobenus rosmarus). Part 1. The skull. Aquat. Mamm. 1990, 16, 101–119. [Google Scholar]
- King, J.E. Some Aspect of the Anatomy of the Ross Seal, Ommatophoca Rossi (Pinnipedia: Phocidae); British Antarctic Survey Scientific Reports; British Antarctic Survey: London, UK, 1963; Volume 63. [Google Scholar]
- Casares-Hidalgo, C.; Perez-Ramos, A.; Forner-Gumbau, M.; Pastor, F.J.; Figueirido, B. Taking a look into the orbit of mammalian carnivorans. J. Anat. 2019, 234, 622–636. [Google Scholar] [CrossRef] [PubMed]
- Constantinescu, G.M.; Moore, C.P. Clinical anatomy of the eyelids for small animal practitioners. Wien. Tierärztliche Mon. 1998, 85, 229–232. [Google Scholar]
- Riedman, M. The Pinnipeds. Seals, Sea Lions and Walruses; University of California Press: Berkeley, CA, USA, 1990. [Google Scholar]
- Martin, A.L.; Irizarry-Rovira, A.R.; Bevier, D.E.; Glickman, L.G.; Glickman, N.W.; Hullinger, R.L. Histology of ferret skin, preweaning to adulthood. Vet. Dermatol. 2007, 18, 401–411. [Google Scholar] [CrossRef]
- Gulland, F.M.D.; Dierauf, L.A.; Withman, K.L. CRC Handbook of Marine Mammal Medicine, 3rd ed.; CRC Press: Boca Raton, FL, USA; Taylor & Francis Inc: London, UK, 2018; pp. 517–536. [Google Scholar]
- Kelleher Davis, R.; Doane, M.G.; Knop, E.; Knop, N.; Dubielzig, R.R.; Colitz, C.M.H.; Argüeso, P.; Sullivan, D.A. Characterization of ocular gland morphology and tear composition of pinnipeds. Vet. Ophthalmol. 2013, 16, 269–275. [Google Scholar] [CrossRef] [Green Version]
- Colitz, C.M.H.; Kelleher Davis, R.; Knop, E.; Knop, N.; Dubielzig, R.R. Description of lacrimal gland of Pinnipeds. Int. Assoc. Aquat. Anim. Med. 2012; in press. [Google Scholar]
- Dartt, D.A. Formation and function of the tear film. In Adler’s Physiology of the Eye; Levin, L.A., Nilsson, S.F.E., VerHoeye, J., Wu Edinburgh, S.M., Eds.; Elsevier Saunders: Amsterdam, The Netherlands, 2011; pp. 350–362. [Google Scholar]
- Davidson, H.J.; Kuonen, V.J. The tear film and ocular mucins. Vet. Ophthalmol. 2004, 7, 71–77. [Google Scholar] [CrossRef]
- Moore, C.P.; Wilsman, N.J.; Nordheim, E.V.; Majors, L.J.; Colier, L.L. Density and distribution of canine conjunctival goblet cells. Investig. Ophthalmol. Vis. Sci. 1987, 28, 1925–1932. [Google Scholar]
- Moore, J.C.; Tiffany, J.M. Human ocular mucus. Origins and preliminary characterisation. Exp. Eye Res. 1979, 29, 291–301. [Google Scholar] [CrossRef]
- Knop, E.; Knop, N. The role of eye-associated lymphoid tissue in corneal immune protection. J. Anat. 2005, 206, 271–285. [Google Scholar] [CrossRef] [PubMed]
- Knop, E.; Knop, N.; Claus, P. Local production of secretory IgA in the eye-associated lymphoid tissue (EALT) of the normal human ocular surface. Investig. Ophthalmol. Vis. Sci. 2008, 49, 2322–2329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knop, E.; Knop, N. A functional unit for ocular surface immune defence formed by the lacrimal gland, conjunctiva and lacrimal drainage system. Adv. Exp. Med. Biol. 2002, 506, 835–844. [Google Scholar] [PubMed]
- Wenzel-Hora, B.I.; Seifert, H.M.; Grüntzig, J. Animal experimental studies of indirect lymphography of the eye, face, and neck regions using Iotasul. Lymphology 1982, 15, 32–35. [Google Scholar]
- Mastropasqua, R.; Agnifili, L.; Fasanella, V.; Nubile, M.; Gnama, A.A.; Falconio, G.; Perri, P.; Di Staso, S.; Mariotti, C. The Conjunctiva-Associated Lymphoid Tissue in chronic ocularsurface diseases. Microsc. Microanal. 2017, 23, 697–707. [Google Scholar] [CrossRef]
- Cazacu, P. Researches Concerning the Morphology of the Nictitating Gland in Dogs. PhD Thesis, IASI, Darmstadt, Germany, 2010. [Google Scholar]
- Park, S.A.; Taylor, K.T.; Zwingenberger, A.L.; Toupadakis, C.h.A.; Marfurt, C.F.; Good, K.L.; Murphy, C.J. Gross anatomy and morphometric evaluation of the canine and third eyelid glands. Vet. Ophthalmol. 2016, 19, 230–236. [Google Scholar] [CrossRef]
- Cabral, V.P.; Laus, J.L.; Dagli, M.L.Z.; PereiraIvia, G.T.; Talieri, I.C.; Monteiro, E.R.; Mamede, F.V. Canine lacrimal and third eyelid superficial glands macroscopic and morphometric characteristics. Cienc. Rural. 2005, 35, 391–397. [Google Scholar] [CrossRef]
- Tarpley, R.J.; Ridgway, S.H. Orbital gland structure and secretions in the Atlantic bottlenose dolphin (Tursiops truncatus). J. Morphol. 1991, 207, 173–184. [Google Scholar] [CrossRef]
- Funasaka, N.; Yoshioka, M.; Fujise, Y. Features of the ocular Harderian gland in three Balaenopterid species based on anatomical, histological and histochemical observations. Mammal Study 2010, 35, 9–15. [Google Scholar] [CrossRef]
- Rodrigues, F.M.; Silva, F.M.; Trompieri-Silveira, A.C.; Vergara-Parente, J.E.; Miglino, M.A.; Guimarães, J.P. Morphology of the eyeball from the Humpback whale (Megaptera novaeangliae). Mic Res. Tech. 2014, 77, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Colitz, C.M.H.; Kelleher Davis, R.; Knop, E.; Knop, N.; Sullivan, D.A.; Argueso, P. Description of the tear glands of pinnipeds. Vet. Ophthalmol. 2011, 14, 422. [Google Scholar]
- Constantinescu, G.M.; McClure, R.C. Anatomy of the orbital fasciae and the third eyelid in dogs. Am. J. Vet. Res. 1990, 51, 260–263. [Google Scholar] [PubMed]
- Goller, T.; Weyrauch, K.D. The conjunctival epithelium of the dogs. Ann. Anat. 1993, 175, 127–134. [Google Scholar]
- Zurano, J.P.; Pablo Ariel Martinez, P.A.; Canto-Hernandez, J.; Montoya-Burgos, J.I.; Costa, G.C. Morphological and ecological divergence in South American canids. J. Biogeogr. 2017, 44, 821–833. [Google Scholar] [CrossRef]
- Carpenter, W.B. Zoology, A Systematic Account; Henry, G., Ed.; British Museum: London, UK, 1857. [Google Scholar]
- Martin, C.h.L.; Pickett, J.P.; Spiess, B.M. Ophthalmic Disease in Veterinary Medicine, 2nd ed.; CRC Press Taylor & Francis Group: Boca Raton, FL, USA, 2020. [Google Scholar]
- Paule, W.J. The comparative histochemistry of the Harderian gland. PhD Dissertation, The Ohio State University, Columbus, OH, USA, 1957. [Google Scholar]
- Owen, R. On the anatomy of the Walurus. Proc. Zool. Soc. Lond. 1853, 23, 103–106. [Google Scholar]
- Pütter, A. Die Augen der Wassersaügethiere (in German). Zool. Jahrb. Jena Gustaf. Fisher Verlag. 1903, 17, 97–402. [Google Scholar]
- El-naseery, N.I.; El-behery, E.I.; El-Ghazali, H.M.; El-Hady, E. The structural characterization of the lacrimal gland in the adult dog (Canis familiaris). Benha Vet. Miedical J. 2016, 31, 106–116. [Google Scholar] [CrossRef]
- Martin, C.h.L.; Munnell, J.; Kaswan, R. Normal ultrastructure and histochemical characteristics of the canine lacrimal gland. Am. J. Vet. Res. 1988, 49, 1566–1572. [Google Scholar]
- Shaker, M.M.; Walaa, F.O.A. Anatomical and histological study of the lacrimal gland of the adult male dog (Canis familiaris). Glob. J. Bio-Sci. Biotechnol. 2016, 5, 520–524. [Google Scholar]
- Zwingenberger, A.L.; Park, A.A.; Murphy, C.J. Computed tomographic imaging characteristics of the normal canine lacrimal glands. BMC Vet. Res. 2014, 10, 116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saito, A.; Watanabe, Y.; Kotani, T. Morphologic changes of the anterior corneal epithelium caused by third eyelid removal in dogs. Vet. Ophthalmol. 2004, 7, 113–119. [Google Scholar] [CrossRef] [PubMed]
- Igado, O.O. Skull typology and morphometric of the Nigerian local dog (Canis lupus familiaris). Niger J. Physiol. Sci. 2017, 32, 153–158. [Google Scholar] [PubMed]
- Johnson, G.L. Ophthalmoscopic studies on the eyes of mammals. Philos. Trans. R. Soc. Lond. Ser. B Biol. Sci. 1968, 254, 207–220. [Google Scholar]
- Mowat, F.M.; Petersen-Jones, S.M.; Williamson, H.; Williams, D.L.; Luthert, P.J.; Ali, R.R.; Bainbridge, J.W. Topographical characterization of cone photoreceptors and the area centralis of the canine retina. Mol. Vis. 2008, 14, 2518–2527. [Google Scholar]
- Schmitt, E.; Wallace, S. Shape change and variation in the cranial morphology of wild canids (Canis lupus, Canis latrans, Canis rufus) compared to domestic dogs (Canis familiaris) using geometric morphometrics. Int. J. Osteoarcheol. 2014, 24, 42–50. [Google Scholar] [CrossRef]
- Salguero, R.; Johnson, V.; Williams, D.; Hartley, C.; Holmes, M.; Dennis, R.; Herrtage, M. CT dimensions, volume and densities of normal canine eyes. Vet. Rec. 2015, 176, 386. [Google Scholar] [CrossRef]
- Renzo, R.; Aldrovani, M.; Crivelaro, R.M.; Thiesen, R.; de Barros Sobrinho, A.A.F.; Balthazar da Silveira, C.P.; Garcia, A.P.; Campos, G.C.S.; Werther, K.; Laus, J.L. The eye of crab-eating fox (Cerdocyon thous): Anatomical characteristics and normative values of the selected diagnostic tests, morphometry of corneal tissue, and arrangements of corneal stromal collagen fibres. J. Zoo Wildl. Med. 2020, 51, 280–289. [Google Scholar] [CrossRef]
- Segura, V.; Prevosti, F. A quantitative approach of the cranial ontogeny of Lycalopex culpaeus (Carnivora: Canidae). Zoomorphology 2012, 131, 79–92. [Google Scholar] [CrossRef]
- Rexford, D. The lens as an indicator of age in the gray fox. J. Mammol. 1961, 42, 109–111. [Google Scholar]
- Peichl, L.; Dubielzig, R.R.; Kubber-Heiss, A.; Schubert, C.; Ahnelt, P.K. Retinal cone types in brown bears and the polar bear indicate dichromatic color vision (abstract). Investig. Ophthalmol. Vis. Sci. 2005, 46, 4539. [Google Scholar]
- Mass, A.M.; Supin, A.Y. Peak density, size and regional distribution of ganglion cells in the retina of the fur seal Callorhinus ursinus. Brain Behav. Evol. 1992, 39, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Turner, E.C.; Sawyer, E.K.; Kaas, J.H. Optic nerve, superior colliculus, visual thalamus, and primary visual cortex of the northern elephant seal (Mirounga angustirostris) and California sea lion (Zalophus californianus). J. Comp. Neurol. 2017, 525, 2109–2132. [Google Scholar] [CrossRef]
- Ninomiya, H.; Imamura, E.; Inomata, T. Comparative anatomy of the ophthalmic rete and its relationship to ocular blood flow in three of marine mammal. Vet. Ophthalmol. 2014, 17, 100–105. [Google Scholar] [CrossRef]
- Levenson, D.H.; Schusterman, R.J. Dark adaptation and visual sensitivity in shallow and deep diving pinnipeds. Mar. Mamm. Sci. 1999, 15, 1303–1313. [Google Scholar] [CrossRef]
- Hanke, F.D.; Hanke, W.; Scholtyssek, C.; Dehnhardt, G. Basic mechanisms in pinniped vision. Exp. Brain Res. 2009, 199, 299–311. [Google Scholar] [CrossRef]
- Ye, Y.; Doung, T.A.D.; Saito, K.; Shinmyo, Y.; Ichikawa, Y.; Higashide, T.; Kagami, K.; Fujiwara, H.; Sugiyama, K.; Kawasaki, H. Visualization of the retina in intact eyes of mice and ferrets using a tissue clearing method. Transl. Vis. Sci. Technol. 2020, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Friede, H.; Kiliardis, S. Macroscopic and roentgenographic anatomy of the skull of the ferret (Mustela putorius furo). Laboratoty Anim. 2002, 36, 86–96. [Google Scholar] [CrossRef] [Green Version]
- Hernandez-Guerra, A.M.; Rodilla, V.; Lopez-Murcia, M.M. Ocular biometry in the adult anesthetized ferret (Mustela putorius furo). Vet. Ophthalmol. 2007, 10, 50–52. [Google Scholar] [CrossRef] [PubMed]
- Tjälve, H.; Frank, A. Tapetum lucidum in the pigmented and albino ferret. Exp. Eye Res. 1984, 38, 341–351. [Google Scholar] [CrossRef]
- Braekevelt, C.R. Fine structure of the retinal epthelium and tapetum lucidum of the ranch mink Mustela vision. Acta Anat. 1989, 135, 296–302. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Examined Structure | PAS | AB pH 1.0 | AB pH 2.5 | AB pH 2.5/PAS | HDI | |
|---|---|---|---|---|---|---|
| upper and lower eyelids | tarsal glands | − | − | − | − | − |
| sebaceous glands | − | − | − | − | − | |
| ciliary glands | −/+ | − | −/+ | −/+ (blue color) | ++ | |
| goblet cells | +++ | +++ | +++ | +++ (magenta color) | +++ | |
| superficial gland of the third eyelid | serous acini | −/+ | − | −/+ | + (blue color) | − |
| mucous acini | ++ | − | −/+ | ++ (magenta color) | ++ | |
| third eyelid | goblet cells in the palpebral and bulbar conjunctiva | +++ | ++/+++ | +++ | +++ (magenta color) | +++ |
| lacrimal gland | serous acini | + | − | − | ++ (magenta color) | − |
| mucous acini | ++ | − | ++ | ++ (blue color) | + | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Paszta, W.; Goździewska-Harłajczuk, K.; Klećkowska-Nawrot, J. Morphology and Histology of the Orbital Region and Eye of the Asiatic Black Bear (Ursus thibetanus)—Similarities and Differences within the Caniformia Suborder. Animals 2022, 12, 801. https://doi.org/10.3390/ani12070801
Paszta W, Goździewska-Harłajczuk K, Klećkowska-Nawrot J. Morphology and Histology of the Orbital Region and Eye of the Asiatic Black Bear (Ursus thibetanus)—Similarities and Differences within the Caniformia Suborder. Animals. 2022; 12(7):801. https://doi.org/10.3390/ani12070801
Chicago/Turabian StylePaszta, Wojciech, Karolina Goździewska-Harłajczuk, and Joanna Klećkowska-Nawrot. 2022. "Morphology and Histology of the Orbital Region and Eye of the Asiatic Black Bear (Ursus thibetanus)—Similarities and Differences within the Caniformia Suborder" Animals 12, no. 7: 801. https://doi.org/10.3390/ani12070801
APA StylePaszta, W., Goździewska-Harłajczuk, K., & Klećkowska-Nawrot, J. (2022). Morphology and Histology of the Orbital Region and Eye of the Asiatic Black Bear (Ursus thibetanus)—Similarities and Differences within the Caniformia Suborder. Animals, 12(7), 801. https://doi.org/10.3390/ani12070801