
The High Testosterone Concentrations of the Bucks Used in the “Male Effect” Is Not a Prerequisite for Obtaining High Ovarian Activity in Goats from Mediterranean Latitudes

 , , ,
, , ,  and
and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Conditions
2.2. Experiment 1
2.2.1. Treatment of Bucks
2.2.2. Experimental Design
2.3. Experiment 2
2.3.1. Preparation of Bucks
2.3.2. Experimental Design
- (1)
- 14 does exposed to the bucks of the Photo group.
- (2)
- 13 does exposed to the bucks of the Photo-Ago Long group.
- (3)
- 15 does exposed to the bucks of the Photo-Ago Short group.
- (4)
- 13 does exposed to the bucks of the Natural-Ago Short group.
- (5)
- 15 does exposed to the bucks of the Natural group.
2.3.3. Buck Odor and Scrotal Circumference
2.4. Methodology Common to Both Experiments
2.4.1. Management
2.4.2. Detection of Estrous Behavior and Ovulation
2.4.3. Analysis of Plasma Progesterone
2.4.4. Fecundity, Fertility, Prolificacy, and Productivity
2.4.5. Buck Plasma Testosterone
2.4.6. Sexual Behavior of the Bucks
2.5. Statistical Analyses
3. Results
3.1. Experiment 1
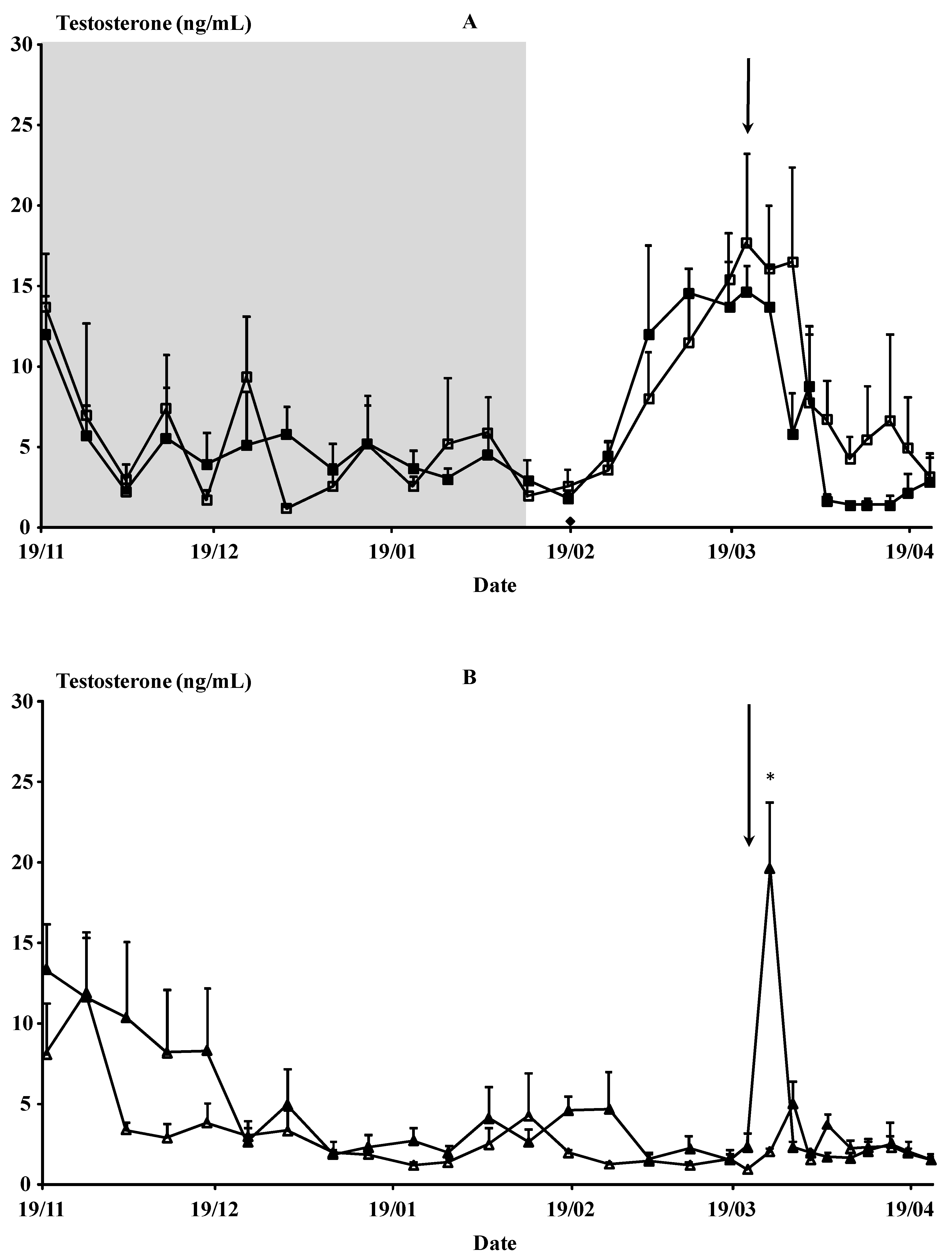
3.1.1. Testosterone Concentrations and Sexual Behavior
3.1.2. Female Reproductive Response
3.2. Experiment 2
3.2.1. Testosterone Concentrations, Odor, Scrotal Circumference, and Sexual Interactions with Bucks
3.2.2. Female Reproductive Response
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zarazaga, L.A.; Celi, I.; Guzmán, J.L.; Malpaux, B. The effect of nutrition on the neural mechanisms potentially involved in melatonin-stimulated LH secretion in female Mediterranean goats. J. Endocrinol. 2011, 211, 263–272. [Google Scholar] [CrossRef] [PubMed]
- Zarazaga, L.A.; Celi, I.; Guzmán, J.L.; Malpaux, B. The response of luteinizing hormone secretion to photoperiod is modified by the level of nutrition in female Mediterranean goats. Anim. Reprod. Sci. 2011, 126, 83–90. [Google Scholar] [CrossRef] [PubMed]
- Zarazaga, L.A.; Celi, I.; Guzmán, J.L.; Malpaux, B. The role of nutrition in the regulation of luteinizing hormone secretion by the opioidergic, dopaminergic and serotoninergic system in female Mediterranean goat. Biol. Reprod. 2011, 84, 447–454. [Google Scholar] [CrossRef] [PubMed]
- Chemineau, P.; Malpaux, B.; Brillard, J.P.; Fostier, A. Seasonality of reproduction and production in farm fishes, birds and mammals. Animal 2007, 1, 419–432. [Google Scholar] [CrossRef] [Green Version]
- Gallego-Calvo, L.; Gatica, M.C.; Celi, I.; Guzmán, J.L.; Delgadillo, J.A.; Zarazaga, L.A. No previous isolation of female goats is required for novel males to induce a male effect, especially if direct physical contact is established. Theriogenology 2014, 82, 1310–1315. [Google Scholar] [CrossRef]
- Flores, J.A.; Véliz, F.G.; Pérez-Villanueva, J.A.; Martínez de la Escalera, G.; Chemineau, P.; Poindron, P.; Malpaux, B.; Delgadillo, J.A. Male reproductive condition is the limiting factor of efficiency in the male effect during seasonal anestrus in female goats. Biol. Reprod. 2000, 62, 1409–1414. [Google Scholar] [CrossRef] [Green Version]
- Rouger, Y. Etude des interactions de l’environnement et des hormones sexuelles dans la régulation du comportement sexuel des bovidae. Ph.D. Thesis, University of Rennes, Rennes, France, 1974; 197p. [Google Scholar]
- Ahmad, N.; Noakes, D.E. Seasonal variations in testis size, libido and plasma testosterone concentrations in British goats. Anim. Sci. 1995, 61, 553–559. [Google Scholar] [CrossRef]
- Delgadillo, J.A.; Flores, J.A.; Véliz, F.G.; Hernández, H.F.; Duarte, G.; Vielma, J.; Poindron, P.; Chemineau, P.; Malpaux, B. Induction of sexual activity in lactating anovulatory female goats using male goats treated only with artificially long days. J. Anim. Sci. 2002, 80, 2780–2786. [Google Scholar] [CrossRef]
- Chasles, M.; Chesneau, D.; Moussu, C.; Delgadillo, J.A.; Chemineau, P.; Keller, M. Sexually active bucks are efficient to stimulate female ovulatory activity during the anestrous season also under temperate latitudes. Anim. Reprod. Sci. 2016, 168, 86–91. [Google Scholar] [CrossRef]
- Zarazaga, L.A.; Gatica, M.C.; Celi, I.; Guzmán, J.L.; Malpaux, B. Effect of artificial long days and/or melatonin treatment on the sexual activity of Mediterranean bucks. Small. Rumin. Res. 2010, 93, 110–118. [Google Scholar] [CrossRef]
- Zarazaga, L.A.; Gatica, M.C.; Gallego-Calvo, L.; Guzmán, J.L. When using photostimulated bucks to induce the male effect in female goats living at Mediterranean latitudes, a male: Female ratio of 1:20 is optimum. J. Appl. Anim. Res. 2018, 46, 883–887. [Google Scholar] [CrossRef]
- Zarazaga, L.A.; Gatica, M.C.; Hernández, H.; Keller, M.; Chemineau, P.; Delgadillo, J.A.; Guzmán, J.L. The reproductive response to the male effect of 7- or 10-month-old female goats is improved when photostimulated males are used. Animal 2019, 13, 1658–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarazaga, L.A.; Gatica, M.C.; Hernández, H.; Chemineau, P.; Delgadillo, J.A.; Guzmán, J.L. Photoperiod-treated bucks are equal to melatonin-treated bucks for inducing reproductive behaviour and physiological functions via the “male effect” in Mediterranean goats. Anim. Reprod. Sci. 2019, 202, 58–64. [Google Scholar] [CrossRef] [PubMed]
- Okamura, H.; Murata, K.; Sakamoto, K.; Wakabayashi, Y.; Ohkura, S.; Takeuchi, Y.; Mori, Y. Male effect pheromone tickles the gonadotrophin-releasing hormone pulse generator. J. Neuroendocr. 2010, 22, 825–832. [Google Scholar] [CrossRef] [PubMed]
- Delgadillo, J.A.; Leboeuf, B.; Chemineau, P. Abolition of seasonal variations in semen quality and maintenance of sperm fertilizing ability by photoperiodic cycles in goat bucks. Small Rumin. Res. 1992, 9, 47–59. [Google Scholar] [CrossRef]
- Delgadillo, J.A.; Cortez, M.E.; Duarte, G.; Chemineau, P.; Malpaux, B. Evidence that the photoperiod controls the annual changes in testosterone secretion, testicular and body weight in subtropical male goats. Reprod. Nutr. Dev. 2004, 44, 183–193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zarazaga, L.A.; Gatica, M.C.; Delgado-Pertiñez, M.; Hernández, H.; Guzmán, J.L.; Delgadillo, J.A. Photoperiod-treatment in Mediterranean bucks can improve the reproductive performance of the male effect depending on the extent of their seasonality. Animals 2021, 11, 400. [Google Scholar] [CrossRef]
- Sanford, L.M.; Palmer, W.M.; Howland, B.E. Influence of sexual activity on serum levels of LH and testosterone in the ram. Can. J. Anim. Sci. 1974, 54, 579–585. [Google Scholar] [CrossRef]
- Yarney, T.A.; Sanford, L.M. The reproductive endocrine response of adult rams to sexual encounters with estrual ewes is season dependent. Horm. Behav. 1983, 17, 169–182. [Google Scholar] [CrossRef]
- Howland, B.E.; Sanford, L.M.; Palmer, W.M. Changes in the serum levels of LH, FSH, prolactin, testosterone and cortisol associated with season and mating in male pygmy goats. J. Androl. 1985, 6, 89–96. [Google Scholar] [CrossRef]
- Walkden-Brown, S.W.; Restall, B.J.; Norton, B.W.; Scaramuzzi, R. The ‘female effect’ in Australian cashmere goats: Effect of season and quality of diet on the LH and testosterone response of bucks to oestrous does. J. Reprod. Fertil. 1994, 100, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Tilbrook, A.J.; Pearce, D.T.; Akkari, L.; Lindsay, D.R. Stimulation of LH secretion in rams by association with ewes. Proc. Aust. Soc. Reprod. Biol. 1983, 15, 50. [Google Scholar]
- Gonzalez, R.; Orgeur, P.; Poindron, P.; Signoret, J.P. Female effect in sheep. I The effects of sexual receptivity of females and the sexual experience of rams. Reprod. Nutr. Dev. 1991, 31, 97–102. [Google Scholar] [CrossRef] [PubMed]
- Rosa, H.J.D.; Juniper, D.T.; Bryant, M.J. The effect of exposure to oestrous ewes on rams’ sexual behaviour, plasma testosterone concentration and ability to stimulate ovulation in seasonally anoestrous ewes. Appl. Anim. Behav. Sci. 2000, 67, 293–305. [Google Scholar] [CrossRef]
- Schanbacher, B.D.; Orgeur, P.; Pelletier, J.; Signoret, J.P. Behavioural and hormonal responses of sexually experienced Ile-de-France rams to oestrous females. Anim. Reprod. Sci. 1987, 14, 293–300. [Google Scholar] [CrossRef]
- Gonzalez, R.; Poindron, P.; Signoret, J.P. Temporal variation in LH and testosterone responses of rams after the introduction of oestrous females during the breeding season. J. Reprod. Fertil. 1988, 83, 201–208. [Google Scholar] [CrossRef] [Green Version]
- Hileman, S.M.; Jackson, G.L. Regulation of gonadotrophin-releasing hormone secretion by testosterone in male sheep. J. Reprod. Fertil. 1999, 54 (Suppl. 54), 231–242. [Google Scholar] [CrossRef]
- Adams, T.E. Using gonadotropin-releasing hormone (GnRH) and GnRH analogs to modulate testis function and enhance the productivity of domestic animals. Anim. Reprod. Sci. 2005, 88, 127–139. [Google Scholar] [CrossRef]
- Padula, A.M. GnRH analogues-agonists and antagonists. Anim. Reprod. Sci. 2005, 88, 115–126. [Google Scholar] [CrossRef]
- Trigg, T.E.; Doyle, A.G.; Walsh, J.D.; Swangchan-Uthai, T. A review of advances in the use of the GnRH agonist deslorelin in control of reproduction. Theriogenology 2006, 66, 1507–1512. [Google Scholar] [CrossRef]
- Janett, F.; Gerig, T.; Tschuor, A.C.; Amatayakul-Chantler, S.; Walker, J.; Howard, R.; Bollwein, H.; Thun, R. Vaccination against gonadotropin-releasing factor (GnRF) with Bopriva significantly decreases testicular development, serum testosterone levels and physical activity in pubertal bulls. Theriogenology 2012, 78, 182–188. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Junaidi, A.; Williamson, P.E.; Cummins, J.M.; Martin, G.B.; Blackberry, M.A.; Trigg, T.E. Use of a new drug delivery formulation of the gonadotrophin-releasing hormone analogue deslorelin for reversible long-term contraception in male dogs. Reprod. Fertil. Dev. 2003, 15, 317–322. [Google Scholar] [CrossRef] [PubMed]
- Junaidi, A.; Williamson, P.E.; Martin, G.B.; Blackberry, M.A.; Cummins, J.M. Dose-response studies for pituitary and testicular function in male dogs treated with the GnRH superagonist, deslorelin. Reprod. Dom. Anim. 2009, 44, 725–734. [Google Scholar] [CrossRef] [PubMed]
- Kopera, I.; Tuz, R.; Kotula-Balak, M.; Schwarz, T.; Koczanowski, J.; Bilinska, B. Morphofunctional alterations in testicular cells of deslorelin-treated boars: An immunohistochemical study. J. Exp. Zool. Part. Ecol. Genet. Physiol. 2008, 309A, 117–126. [Google Scholar] [CrossRef]
- Kopera, I.; Tuz, R.; Hejmej, A.; Schwarz, T.; Koczanowski, J.; Bilińska, B. Immunolocalization of androgen receptor in the boar epididymis: The effect of GnRH agonist deslorelin. Reprod. Domest. Anim. 2009, 44, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Fraser, H.M.; Lincoln, G.A. Effects of chronic treatment with an LHRH agonist on the secretion of LH, FSH and testosterone in the ram. Biol. Reprod. 1980, 22, 269–276. [Google Scholar] [CrossRef] [Green Version]
- Lincoln, G.A.; Fraser, H.M.; Abbott, M.P. Blockade of pulsatile LH, FSH and testosterone secretion in rams by constant infusion of an LHRH agonist. J. Reprod. Fertil. 1986, 77, 587–597. [Google Scholar] [CrossRef]
- Vickery, B.H.; McRae, G.I.; Briones, W.V.; Roberts, B.B.; Worden, A.C.; Schanbacher, B.; Falvo, R.E. Dose-response studies on male reproductive parameters in dogs with nafarelin acetate, a potent LHRH agonist. J. Androl. 1985, 6, 53–60. [Google Scholar] [CrossRef]
- Gautier, C.; Schmidt, K.; Aurich, J.; Aurichm, C. Effects of implants containing the GnRH agonist deslorelin on testosterone release and semen characteristics in Shetland stallions. Anim. Reprod. Sci. 2018, 195, 230–241. [Google Scholar] [CrossRef]
- Amundson, B.C.; Wheaton, J.E. Effects of chronic LHRH treatment on brain LHRH content, pituitary and plasma LH and ovarian follicular activity in the anestrous ewe. Biol. Reprod. 1979, 20, 633–638. [Google Scholar] [CrossRef]
- Kauffold, J.; Rohrmann, H.; Boehm, J.; Wehrend, A. Effects of long-term treatment with the GnRH agonist deslorelin (Suprelorin®) on sexual function in boars. Theriogenology 2010, 74, 733–740. [Google Scholar] [CrossRef] [PubMed]
- Giriboni, J.; Lacuesta, L.; Santiago-Moreno, J.; Ungerfeld, R. Chronic use of a GnRH-agonist (deslorelin) or immunization against GnRH: Effects on testicular function and sperm quality of bucks. Domest. Anim. Endocrinol. 2002, 71, 106395. [Google Scholar] [CrossRef] [PubMed]
- Walkden-Brown, S.W.; Restall, B.J.; Norton, B.W.; Scaramuzzi, R.; Martin, G. Effect of nutrition on seasonal patterns of LH, FSH and testosterone concentration, testicular mass, sebaceous gland volume and odour in Australian cashmere goats. J. Reprod. Fertil. 1994, 102, 351–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morand-Fehr, P.; Sauvant, D. Goat nutrition. In Alimentation des Bovins, Ovins et Caprins; Jarrige, R., Ed.; Institut National de la Recherche Agronomique: Paris, France, 1988; pp. 281–304. [Google Scholar]
- Walkden-Brown, S.W.; Restall, B.J.; Henniawati. The male effect in the Australian cashmere goat. 2. Enhancement with buck nutrition and use of oestrous female. Anim. Reprod. Sci. 1993, 32, 69–84. [Google Scholar] [CrossRef]
- Chemineau, P.; Malpaux, B.; Delgadillo, J.A.; Guerin, Y.; Ravault, J.P.; Thimonier, J.; Pelletier, J. Control of sheep and goat reproduction: Use of light and melatonin. Anim. Reprod. Sci. 1992, 30, 157–184. [Google Scholar] [CrossRef]
- Andueza, D.; Alabart, J.L.; Lahoz, B.; Muñoz, F.; Folch, J. Early pregnancy diagnosis in sheep using near-infrared spectroscopy on blood plasma. Theriogenology 2014, 81, 509–513. [Google Scholar] [CrossRef]
- Schrick, F.N.; Surface, R.A.; Pritchard, J.Y.; Dailey, R.A.; Townsend, E.C.; Inskeep, E.K. Ovarian structures during the estrous cycle and early pregnancy in ewes. Biol. Reprod. 1993, 49, 1133–1140. [Google Scholar] [CrossRef] [Green Version]
- Ungerfeld, R.; Ramos, M.A.; González-Pensado, S.P. Ram effect: Adult rams induce a greater reproductive response in anestrous ewes than yearling rams. Anim. Reprod. Sci. 2008, 103, 271–277. [Google Scholar] [CrossRef]
- Zarazaga, L.A.; Guzmán, J.L.; Domínguez, C.; Pérez, M.C.; Prieto, R. Effects of season and feeding level on reproductive activity and semen quality in Payoya buck goats. Theriogenology 2009, 71, 1316–1325. [Google Scholar] [CrossRef]
- Walkden-Brown, S.W.; Restall, B.J.; Scaramuzzi, R.; Martin, G.B.; Blackberry, M.A. Seasonality in male Australian cashmere goats: Long term effects of castration and testosterone or oestradiol treatment on changes in LH, FSH and prolactin concentrations, and body growth. Small Rumin. Res. 1997, 26, 239–252. [Google Scholar] [CrossRef]
- Jenkinson, D.M.; Blackbum, P.S.; Proudfoot, R. Seasonal changes in the skin glands of the goat. Br. Vet. J. 1967, 123, 541–549. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | Natural (n = 27) | Photo (n = 27) | Significance 1 |
|---|---|---|---|
| Females ovulating (%) | 89 | 100 | NS |
| Interval between the introduction of males and normal ovulation | 10.21 ± 0.75 | 11.04 ± 0.58 | NS |
| Females in estrous and ovulating (%) | 74 | 96 | * |
| Interval between the introduction of males and estrous | 6.50 ± 0.58 | 6.36 ± 0.48 | NS |
| Ovulation rate | 1.57 ± 0.13 | 1.78 ± 0.14 | NS |
| Fecundity (%) | 85 | 61 | NS |
| Fertility (%) | 56 | 52 | NS |
| Prolificacy | 1.53 ± 0.13 | 1.50 ± 0.14 | NS |
| Productivity | 0.85 ± 0.17 | 0.78 ± 0.16 | NS |
| Variable | Photo (n = 14) | Photo-Ago Long (n = 13) | Photo-Ago Short (n = 15) | Natural-Ago Short (n = 13) | Natural (n = 15) |
|---|---|---|---|---|---|
| Females ovulating (%) | 100 | 92 | 100 | 92 | 100 |
| Interval between the introduction of males and normal ovulation (days) | 6.36 ± 0.46 | 5.83 ± 0.50 | 6.07 ± 0.34 | 6.58 ± 0.66 | 5.20 ± 0.36 |
| Females in estrous and ovulating (%) | 93 | 85 | 93 | 85 | 100 |
| Interval between the introduction of male and estrous (days) | 4.62 ± 0.72 | 3.45 ± 0.80 | 3.07 ± 0.54 | 3.27 ± 0.66 | 2.67 ± 0.40 |
| Fecundity (%) | 92 | 64 | 93 | 73 | 93 |
| Fertility (%) | 86 | 54 | 87 | 62 | 80 |
| Prolificacy | 1.75 ± 0.18 | 1.86 ± 0.14 | 1.31 ± 0.13 | 1.63 ± 0.18 | 1.50 ± 0.15 |
| Productivity | 1.50 ± 0.23 | 1.00 ± 0.28 | 1.13 ± 0.17 | 1.00 ± 0.25 | 1.20 ± 0.20 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zarazaga, L.Á.; Gatica, M.-C.; De La Rosa, I.; Delgado-Pertíñez, M.; Guzmán, J.L. The High Testosterone Concentrations of the Bucks Used in the “Male Effect” Is Not a Prerequisite for Obtaining High Ovarian Activity in Goats from Mediterranean Latitudes. Animals 2022, 12, 954. https://doi.org/10.3390/ani12080954
Zarazaga LÁ, Gatica M-C, De La Rosa I, Delgado-Pertíñez M, Guzmán JL. The High Testosterone Concentrations of the Bucks Used in the “Male Effect” Is Not a Prerequisite for Obtaining High Ovarian Activity in Goats from Mediterranean Latitudes. Animals. 2022; 12(8):954. https://doi.org/10.3390/ani12080954
Chicago/Turabian StyleZarazaga, Luis Ángel, María-Carolina Gatica, Ignacio De La Rosa, Manuel Delgado-Pertíñez, and José Luis Guzmán. 2022. "The High Testosterone Concentrations of the Bucks Used in the “Male Effect” Is Not a Prerequisite for Obtaining High Ovarian Activity in Goats from Mediterranean Latitudes" Animals 12, no. 8: 954. https://doi.org/10.3390/ani12080954
APA StyleZarazaga, L. Á., Gatica, M. -C., De La Rosa, I., Delgado-Pertíñez, M., & Guzmán, J. L. (2022). The High Testosterone Concentrations of the Bucks Used in the “Male Effect” Is Not a Prerequisite for Obtaining High Ovarian Activity in Goats from Mediterranean Latitudes. Animals, 12(8), 954. https://doi.org/10.3390/ani12080954