
Effects of PLIN1 Gene Knockout on the Proliferation, Apoptosis, Differentiation and Lipolysis of Chicken Preadipocytes
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Cell Cultures and Differentiation
2.2. Establishment of a Chicken Preadipocyte Line with PLIN1 Knockout
2.3. Cell Proliferation Assay
2.4. Cell Cycle Assay
2.5. Cell Apoptosis Assay
2.6. Oil Red O Staining and Extraction Assay
2.7. RNA Extraction and RT-qPCR
2.8. Western Blotting
2.9. Triglyceride (TG) Assay
2.10. Assay of HSL and ATGL Activity
2.11. Statistical Analysis
3. Results
3.1. Establishment of a Chicken PLIN1 Knockout Preadipocyte Cell Line
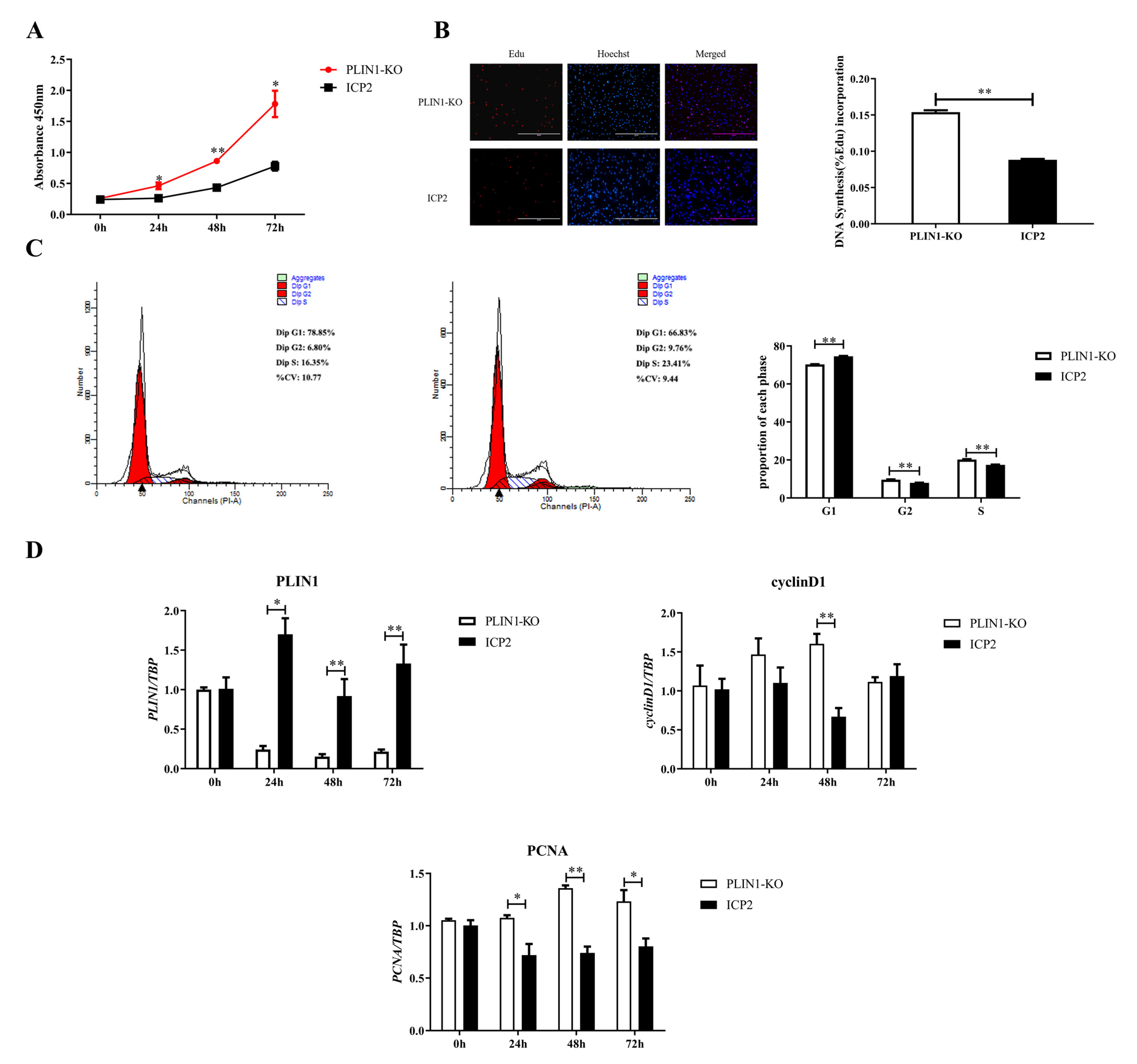
3.2. Knockout of PLIN1 Promotes Proliferation of Chicken Preadipocytes
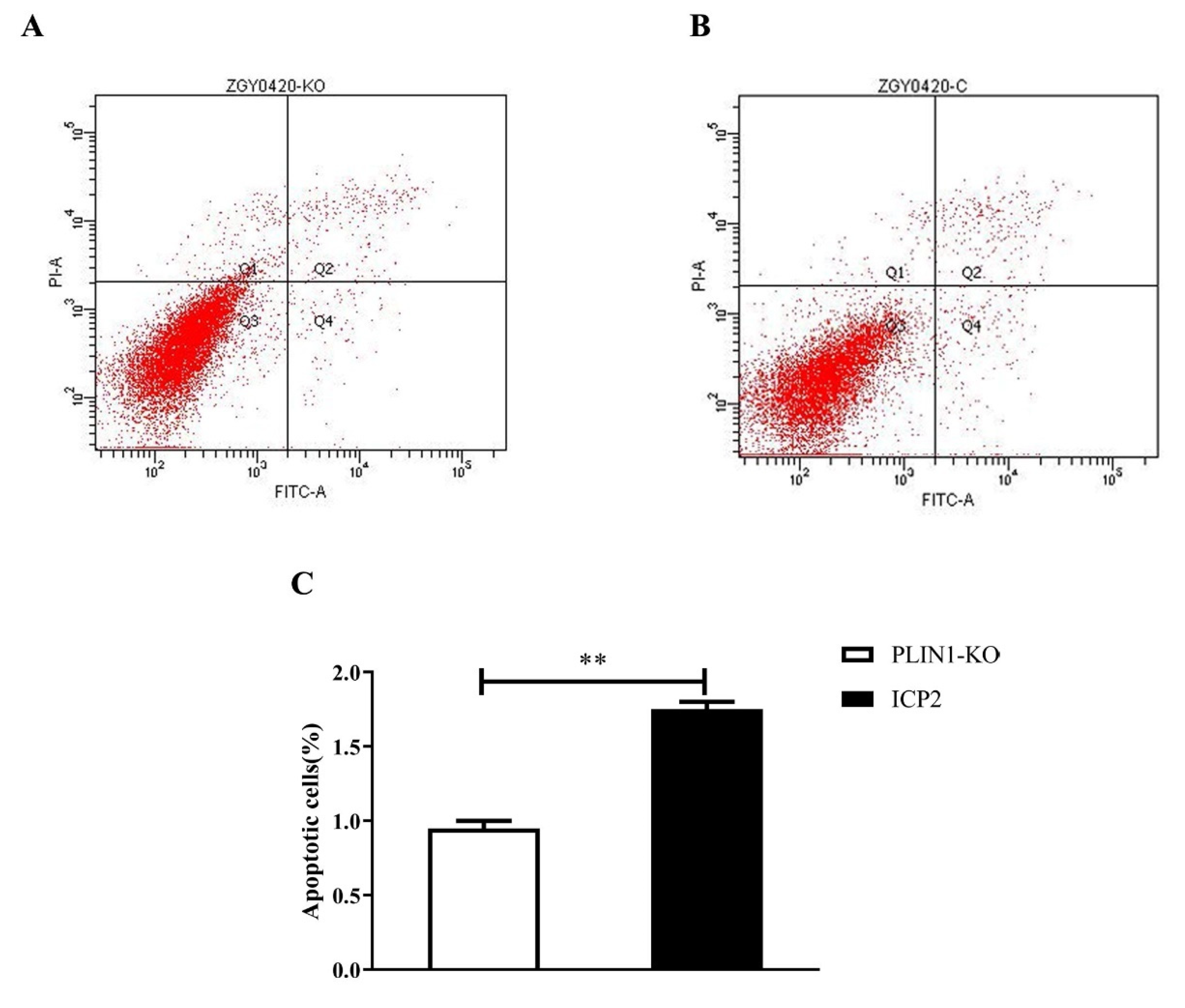
3.3. Knockout of PLIN1 Suppressed Apoptosis of Chicken Preadipocytes
3.4. Knockout of PLIN1 Inhibited the Differentiation of Chicken Preadipocytes
3.5. Effect of PLIN1 Knockout on Lipolysis in Chicken Preadipocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pol, A.; Gross, S.P.; Parton, R.G. Review: Biogenesis of the multifunctional lipid droplet: Lipids, proteins, and sites. J. Cell. Biol. 2014, 204, 635–646. [Google Scholar] [CrossRef] [Green Version]
- Wolins, N.E.; Brasaemle, D.L.; Bickel, P.E. A proposed model of fat packaging by exchangeable lipid droplet proteins. FEBS Lett. 2006, 580, 5484–5491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brasaemle, D.L.; Rubin, B.; Harten, I.A.; Gruia-Gray, J.; Kimmel, A.R.; Londos, C. Perilipin A increases triacylglycerol storage by decreasing the rate of triacylglycerol hydrolysis. J. Biol. Chem. 2000, 275, 38486–38493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinez-Botas, J.; Anderson, J.B.; Tessier, D.; Lapillonne, A.; Chang, B.H.; Quast, M.J.; Gorenstein, D.; Chen, K.H.; Chan, L. Absence of perilipin results in leanness and reverses obesity in Lepr(db/db) mice. Nat. Genet. 2000, 26, 474–479. [Google Scholar] [CrossRef] [PubMed]
- Zhou, W.; Wang, Y.; Li, H. Effects of lipid droplet-coated proteins on lipid accumulation in chicken pre-adipocytes. J. Cell. Mol. Immunol. 2012, 28, 944–947+951. (In Chinese) [Google Scholar]
- Wang, W.; Zhang, T.; Wu, C.; Wang, S.; Wang, Y.; Li, H.; Wang, N. Immortalization of chicken preadipocytes by retroviral transduction of chicken TERT and TR. PLoS ONE 2017, 12, e0177348. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Sun, N.; Zeng, J.; Zeng, Y.; Fan, Y.; Feng, W.; Li, J. Differential expression of miR-672-5p and miR-146a-5p in osteoblasts in rats after steroid intervention. Gene 2016, 591, 69–73. [Google Scholar] [CrossRef]
- Wang, Z.; Zhou, H.; Zheng, H.; Zhou, X.; Shen, G.; Teng, X.; Liu, X.; Zhang, J.; Wei, X.; Hu, Z.; et al. Autophagy-based unconventional secretion of HMGB1 by keratinocytes plays a pivotal role in psoriatic skin inflammation. Autophagy 2021, 17, 529–552. [Google Scholar] [CrossRef]
- Greenberg, A.S.; Egan, J.J.; Wek, S.A.; Garty, N.B.; Blanchette-Mackie, E.J.; Londos, C. Perilipin, a major hormonally regulated adipocyte-specific phosphoprotein associated with the periphery of lipid storage droplets. J. Biol. Chem. 1991, 266, 11341–11346. [Google Scholar] [CrossRef]
- Liu, B.; Wang, Y.; Shi, H.; Wang, Q.; Li, H. Characteristics of C/EBPα antiserum preparation and tissue expression in chickens. J. Agric. Biotechnol. 2009, 17, 47–51. (In Chinese) [Google Scholar]
- Wang, H. Screening and Analysis of Important Genes Affecting Chicken Fat Metabolism. Ph.D. Dissertation, Northeast Agricultural University, Harbin, China, 2008. (In Chinese). [Google Scholar]
- Wroblewska, A.; Dhainaut, M.; Ben-Zvi, B.; Rose, S.A.; Park, E.S.; Amir, E.D.; Bektesevic, A.; Baccarini, A.; Merad, M.; Rahman, A.H.; et al. Protein Barcodes Enable High-Dimensional Single-Cell CRISPR Screens. Cell 2018, 175, 1141–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Concordet, J.P.; Haeussler, M. CRISPOR: Intuitive guide selection for CRISPR/Cas9 genome editing experiments and screens. Nucleic. Acids. Res. 2018, 46, W242–W245. [Google Scholar] [CrossRef] [PubMed]
- Hebert, T.L.; Wu, X.; Yu, G.; Goh, B.C.; Halvorsen, Y.D.; Wang, Z.; Moro, C.; Gimble, J.M. Culture effects of epidermal growth factor (EGF) and basic fibroblast growth factor (bFGF) on cryopreserved human adipose-derived stromal/stem cell proliferation and adipogenesis. J. Tissue Eng. Regen. Med. 2009, 3, 553–561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Janke, J.; Engeli, S.; Gorzelniak, K.; Luft, F.C.; Sharma, A.M. Mature adipocytes inhibit in vitro differentiation of human preadipocytes via angiotensin type 1 receptors. Diabetes 2002, 51, 1699–1707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Levine, J.A.; Jensen, M.D.; Eberhardt, N.L.; O′Brien, T. Adipocyte macrophage colony-stimulating factor is a mediator of adipose tissue growth. J. Clin. Investig. 1998, 101, 1557–1564. [Google Scholar] [CrossRef] [Green Version]
- Saint-Marc, P.; Kozak, L.P.; Ailhaud, G.; Darimont, C.; Negrel, R. Angiotensin II as a trophic factor of white adipose tissue: Stimulation of adipose cell formation. Endocrinology 2001, 142, 487–492. [Google Scholar] [CrossRef]
- Sethi, J.K. Activatin′ human adipose progenitors in obesity. Diabetes 2010, 59, 2354–2357. [Google Scholar] [CrossRef] [Green Version]
- Stewart, A.; Guan, H.; Yang, K. BMP-3 promotes mesenchymal stem cell proliferation through the TGF-beta/activin signaling pathway. J. Cell. Physiol. 2010, 223, 658–666. [Google Scholar]
- Wang, X.; Huang, M.; Wang, Y. The effect of insulin, TNFalpha and DHA on the proliferation, differentiation and lipolysis of preadipocytes isolated from large yellow croaker (Pseudosciaena Crocea R.). PLoS ONE 2012, 7, e48069. [Google Scholar]
- Xing, H.; Northrop, J.P.; Grove, J.R.; Kilpatrick, K.E.; Su, J.L.; Ringold, G.M. TNF alpha-mediated inhibition and reversal of adipocyte differentiation is accompanied by suppressed expression of PPARgamma without effects on Pref-1 expression. Endocrinology 1997, 138, 2776–2783. [Google Scholar] [CrossRef]
- Yamashiro, H.; Inamoto, T.; Yagi, M.; Ueno, M.; Kato, H.; Takeuchi, M.; Miyatake, S.; Tabata, Y.; Yamaoka, Y. Efficient proliferation and adipose differentiation of human adipose tissue-derived vascular stromal cells transfected with basic fibroblast growth factor gene. Tissue Eng. 2003, 9, 881–892. [Google Scholar] [CrossRef] [PubMed]
- Yan, A.; Avraham, T.; Zampell, J.C.; Haviv, Y.S.; Weitman, E.; Mehrara, B.J. Adipose-derived stem cells promote lymphangiogenesis in response to VEGF-C stimulation or TGF-beta1 inhibition. Future Oncol. 2011, 7, 1457–1473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, S.; Raza, S.H.A.; Zhao, C.; Cheng, G.; Zan, L. Overexpression of PLIN1 Promotes Lipid Metabolism in Bovine Adipocytes. Animals 2020, 10, 1944. [Google Scholar] [CrossRef] [PubMed]
- Lyu, Y.; Su, X.; Deng, J.; Liu, S.; Zou, L.; Zhao, X.; Wei, S.; Geng, B.; Xu, G. Defective differentiation of adipose precursor cells from lipodystrophic mice lacking perilipin 1. PLoS ONE 2015, 10, e0117536. [Google Scholar] [CrossRef]
- Dalton, S. Linking the Cell Cycle to Cell Fate Decisions. Trends Cell Biol. 2015, 25, 592–600. [Google Scholar] [CrossRef] [Green Version]
- Schafer, K.A. The cell cycle: A review. Vet. Pathol. 1998, 35, 461–478. [Google Scholar] [CrossRef]
- Tuncer, E.; Turk, N.S.; Arici, S.; Duzcan, S.E.; Demirkan, N.C. Expression of p16 protein and cyclin D1 in periampullary carcinomas. Turk Patoloji Derg. 2011, 27, 17–22. [Google Scholar] [CrossRef]
- Zheng, X.; Wang, Y.; Liu, B.; Liu, C.; Liu, D.; Zhu, J.; Yang, C.; Yan, J.; Liao, X.; Meng, X.; et al. Bmi-1-shRNA inhibits the proliferation of lung adenocarcinoma cells by blocking the G1/S phase through decreasing cyclin D1 and increasing p21/p27 levels. Nucleic Acid Ther. 2014, 24, 210–216. [Google Scholar] [CrossRef] [Green Version]
- Moldovan, G.L.; Pfander, B.; Jentsch, S. PCNA, the maestro of the replication fork. Cell 2007, 129, 665–679. [Google Scholar] [CrossRef] [Green Version]
- Krahmer, N.; Guo, Y.; Wilfling, F.; Hilger, M.; Lingrell, S.; Heger, K.; Newman, H.W.; Schmidt-Supprian, M.; Vance, D.E.; Mann, M.; et al. Phosphatidylcholine synthesis for lipid droplet expansion is mediated by localized activation of CTP: Phosphocholine cytidylyltransferase. Cell Metab. 2011, 14, 504–515. [Google Scholar] [CrossRef] [Green Version]
- Wilfling, F.; Wang, H.; Haas, J.T.; Krahmer, N.; Gould, T.J.; Uchida, A.; Cheng, J.X.; Graham, M.; Christiano, R.; Frohlich, F.; et al. Triacylglycerol synthesis enzymes mediate lipid droplet growth by relocalizing from the ER to lipid droplets. Dev. Cell 2013, 24, 384–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, Z.; Gong, J.; Wu, H.; Xu, W.; Wu, L.; Xu, D.; Gao, J.; Wu, J.W.; Yang, H.; Yang, M.; et al. Perilipin1 promotes unilocular lipid droplet formation through the activation of Fsp27 in adipocytes. Nat. Commun. 2013, 4, 1594. [Google Scholar] [CrossRef] [Green Version]
- Altiok, S.; Xu, M.; Spiegelman, B.M. PPARgamma induces cell cycle withdrawal: Inhibition of E2F/DP DNA-binding activity via down-regulation of PP2A. Genes Dev. 1997, 11, 1987–1998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cui, H.X.; Zheng, M.Q.; Liu, R.R.; Zhao, G.P.; Chen, J.L.; Wen, J. Liver dominant expression of fatty acid synthase (FAS) gene in two chicken breeds during intramuscular-fat development. Mol. Biol. Rep. 2012, 39, 3479–3484. [Google Scholar] [CrossRef] [PubMed]
- Otto, S.; Przetakiewicz, Z. Use and evaluation of the usefulness of synthetic materials—Dallop, Marlex and Gore-tex in alloplasty of ventral hernia. Wiad. Lek. 1990, 43, 858–865. [Google Scholar]
- Conte, M.; Franceschi, C.; Sandri, M.; Salvioli, S. Perilipin 2 and Age-Related Metabolic Diseases: A New Perspective. Trends Endocrinol. Metab. 2016, 27, 893–903. [Google Scholar] [CrossRef] [PubMed]
- Xu, G.; Sztalryd, C.; Lu, X.; Tansey, J.T.; Gan, J.; Dorward, H.; Kimmel, A.R.; Londos, C. Post-translational regulation of adipose differentiation-related protein by the ubiquitin/proteasome pathway. J. Biol. Chem. 2005, 280, 42841–42847. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Chen, X.; Zhang, P.; Liu, G. Perilipin2 is an Earlier Marker Than Perilipin1 for Identifying Adipocyte Regeneration in Fat Grafts. Aesthet. Surg. J. 2021, 41, NP646–NP652. [Google Scholar] [CrossRef]
- Brasaemle, D.L.; Subramanian, V.; Garcia, A.; Marcinkiewicz, A.; Rothenberg, A. Perilipin A and the control of triacylglycerol metabolism. Mol. Cell. Biochem. 2009, 326, 15–21. [Google Scholar] [CrossRef]
- Wang, H.; Hu, L.; Dalen, K.; Dorward, H.; Marcinkiewicz, A.; Russell, D.; Gong, D.; Londos, C.; Yamaguchi, T.; Holm, C.; et al. Activation of hormone-sensitive lipase requires two steps, protein phosphorylation and binding to the PAT-1 domain of lipid droplet coat proteins. J. Biol. Chem. 2009, 284, 32116–32125. [Google Scholar] [CrossRef] [Green Version]
- Li, S. Transcriptional Regulation Mechanism of Bovine PLIN1 Gene and Its Effect on Proliferation, Differentiation and Lipid Metabolism of Precursor Adipocytes. Ph.D. Dissertation, Northwest A&F University, Xianyang, China, 2020. (In Chinese). [Google Scholar]
- Granneman, J.G.; Moore, H.P.; Krishnamoorthy, R.; Rathod, M. Perilipin controls lipolysis by regulating the interactions of AB-hydrolase containing 5 (Abhd5) and adipose triglyceride lipase (Atgl). J. Biol. Chem. 2009, 284, 34538–34544. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haemmerle, G.; Lass, A.; Zimmermann, R.; Gorkiewicz, G.; Meyer, C.; Rozman, J.; Heldmaier, G.; Maier, R.; Theussl, C.; Eder, S.; et al. Defective lipolysis and altered energy metabolism in mice lacking adipose triglyceride lipase. Science 2006, 312, 734–737. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Heckmann, B.L.; Zhang, X.; Smas, C.M.; Liu, J. Distinct mechanisms regulate ATGL-mediated adipocyte lipolysis by lipid droplet coat proteins. Mol. Endocrinol. 2013, 27, 116–126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lampidonis, A.D.; Rogdakis, E.; Voutsinas, G.E.; Stravopodis, D.J. The resurgence of Hormone-Sensitive Lipase (HSL) in mammalian lipolysis. Gene 2011, 477, 1–11. [Google Scholar] [CrossRef]
- Lan, Y.L.; Lou, J.C.; Lyu, W.; Zhang, B. Update on the synergistic effect of HSL and insulin in the treatment of metabolic disorders. Ther. Adv. Endocrinol. Metab. 2019, 10, 2042018819877300. [Google Scholar] [CrossRef]
- Feng, C.; Xu, X.; Dong, W.; Chen, C.; Yan, J. CRISPR/Cas9 knockout of PLIN1 gene enhances lipolytic effect in 3T3-L1 adipo-cytes. Chin. J. Biotechnol. 2020, 36, 1386–1394. (In Chinese) [Google Scholar]
- Zhang, S.; Liu, G.; Xu, C.; Liu, L.; Zhang, Q.; Xu, Q.; Jia, H.; Li, X.; Li, X. Perilipin 1 Mediates Lipid Metabolism Homeostasis and Inhibits Inflammatory Cytokine Synthesis in Bovine Adipocytes. Front. Immunol. 2018, 9, 467. [Google Scholar] [CrossRef] [Green Version]
- Zou, L.; Wang, W.; Liu, S.; Zhao, X.; Lyv, Y.; Du, C.; Su, X.; Geng, B.; Xu, G. Spontaneous hypertension occurs with adipose tissue dysfunction in perilipin-1 null mice. Biochim. Biophys. Acta. 2016, 1862, 182–191. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Name | Oligonucleotide Sequence (5’-3’) |
|---|---|
| PLIN1-2e-g1 | GACCTACACCAGCACCAAAGAGG |
| PLIN1-2e-g2 | CACAAATGGGTGGACCTCTTTGG |
| PLIN1-2e-g3 | GTCTATGAGCAGGGAGTGAAGGG |
| PLIN1-4e-g4 | CCATCCCGGCAGCTGCCATCTGG |
| PLIN1-4e-g5 | ATCCTGGGGCTGGCAGTCGGGGG |
| PLIN1-5e-g6 | AGGTCTCCCAGCAGCAATCCAGG |
| PLIN1-5e-g7 | AGCAGTGCTCCCAGCACCCTGGG |
| PLIN1-6e-g8 | CTGGCGGCAGAGCAGGATGCCGG |
| Primer Name | Primer Sequence (5’-3’) |
|---|---|
| PLIN1-2e-F | TCCCCATGGTGGACCAAAAA |
| PLIN1-2e-R | GCAGGTACTGTCCCACTGTT |
| PLIN1-4e-F | TTCTGTATGGCTGGCTACCTC |
| PLIN1-4e-R | CTTCCTCCACCTCAGCAGAAC |
| PLIN1-5/6e-F | GAGAAAGATCCTGGTGCAGGG |
| PLIN1-5/6e-R | GAGAACATGGTGGGACTGAGA |
| Gene 1 | Accession Number | Primer Sequence (5ʹ to 3ʹ) |
|---|---|---|
| PPARγ | NM_001001460 | F: GTGCAATCAAAATGGAGCC R: CTTACAACCTTCACATGCAT |
| A-FABP | NM_204290 | F: ATGTGCGACCAGTTTGT R: TCACCATTGATGCTGATAG |
| PLIN1 | NM_001127439 | F: GGGGTGACTGGCGGTTGTA R: GCCGTAGAGGTTGGCGTAG |
| FAS | NM_205155 | F: AAGGCGGAAGTCAACGG R: TTGATGGTGAGGAGTCG |
| PLIN2 | NM_001031420.1 | F:TCTTGGGAAGTCGTGTGGTG |
| R: CACGTGCACGGAACTTTGAA | ||
| TBP | NM_205103 | F:GCGTTTTGCTGCTGTTATTATGAG |
| R:TCCTTGCTGCCAGTCTGGAC | ||
| CyclinD1 | NM_205381 | F:CTCGGAGCTACCTGCATGTTT |
| R:GTTTACGGATGATCTGTTTGGTG | ||
| PCNA | NM_204170 | F:GTGCTGGGACCTGGGTT |
| R:CGTATCCGCATTGTCTTCTG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhai, G.; Pang, Y.; Zou, Y.; Wang, X.; Liu, J.; Zhang, Q.; Cao, Z.; Wang, N.; Li, H.; Wang, Y. Effects of PLIN1 Gene Knockout on the Proliferation, Apoptosis, Differentiation and Lipolysis of Chicken Preadipocytes. Animals 2023, 13, 92. https://doi.org/10.3390/ani13010092
Zhai G, Pang Y, Zou Y, Wang X, Liu J, Zhang Q, Cao Z, Wang N, Li H, Wang Y. Effects of PLIN1 Gene Knockout on the Proliferation, Apoptosis, Differentiation and Lipolysis of Chicken Preadipocytes. Animals. 2023; 13(1):92. https://doi.org/10.3390/ani13010092
Chicago/Turabian StyleZhai, Guiying, Yongjia Pang, Yichong Zou, Xinyu Wang, Jie Liu, Qi Zhang, Zhiping Cao, Ning Wang, Hui Li, and Yuxiang Wang. 2023. "Effects of PLIN1 Gene Knockout on the Proliferation, Apoptosis, Differentiation and Lipolysis of Chicken Preadipocytes" Animals 13, no. 1: 92. https://doi.org/10.3390/ani13010092
APA StyleZhai, G., Pang, Y., Zou, Y., Wang, X., Liu, J., Zhang, Q., Cao, Z., Wang, N., Li, H., & Wang, Y. (2023). Effects of PLIN1 Gene Knockout on the Proliferation, Apoptosis, Differentiation and Lipolysis of Chicken Preadipocytes. Animals, 13(1), 92. https://doi.org/10.3390/ani13010092