
Dietary Lactobacillus reuteri SL001 Improves Growth Performance, Health-Related Parameters, Intestinal Morphology and Microbiota of Broiler Chickens


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Strain
2.2. Study Design and Animal Management
2.3. Growth Performance and Carcass Traits
2.4. Measurements of Health-Related Parameters
2.5. Intestinal Morphology
2.6. Total RNA Isolation and Quantitative Real-Time PCR Analysis
2.7. 16S rDNA Sequencing and Data Analysis
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Carcass Traits
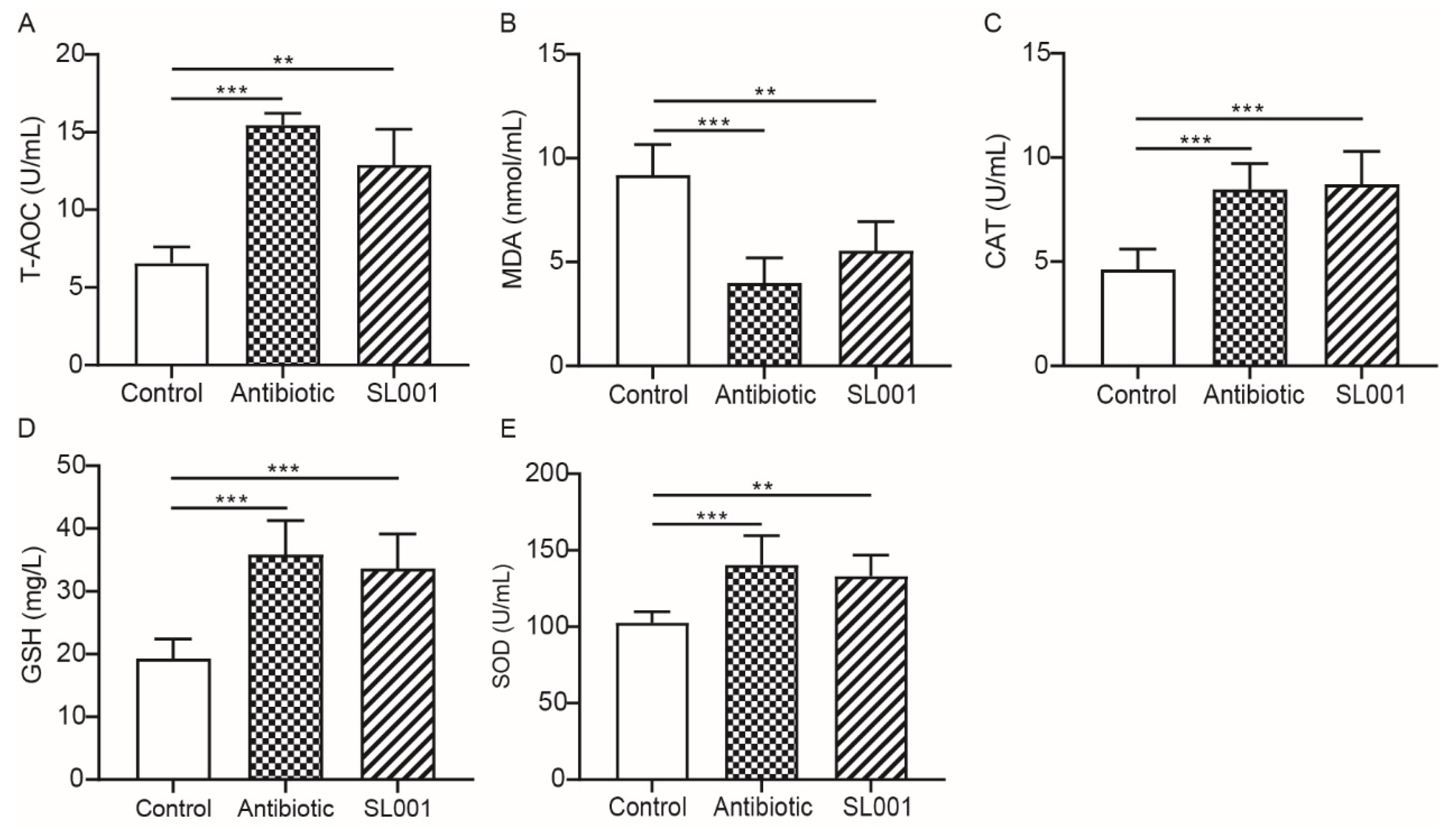
3.3. Health-Related Parameters
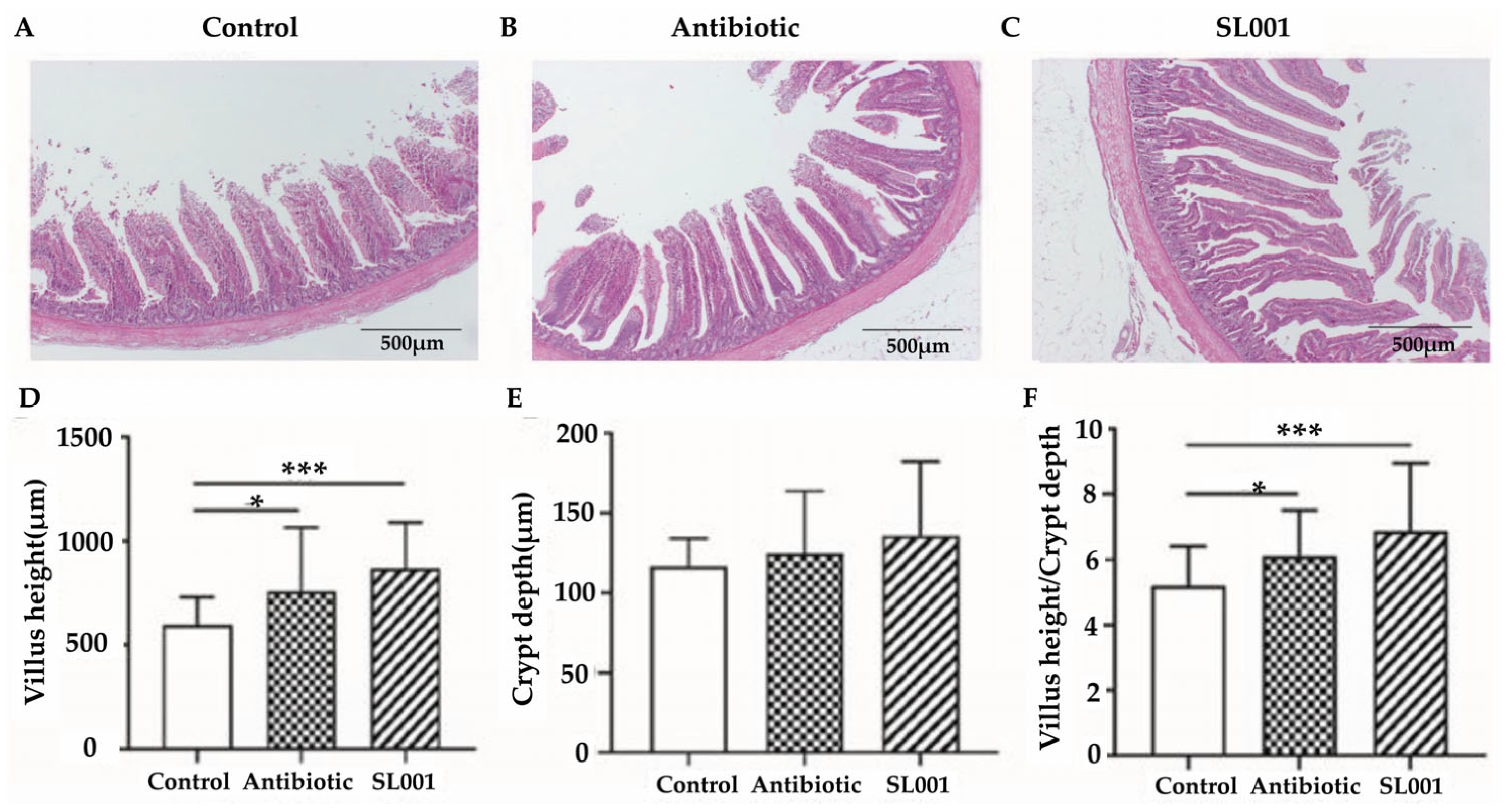
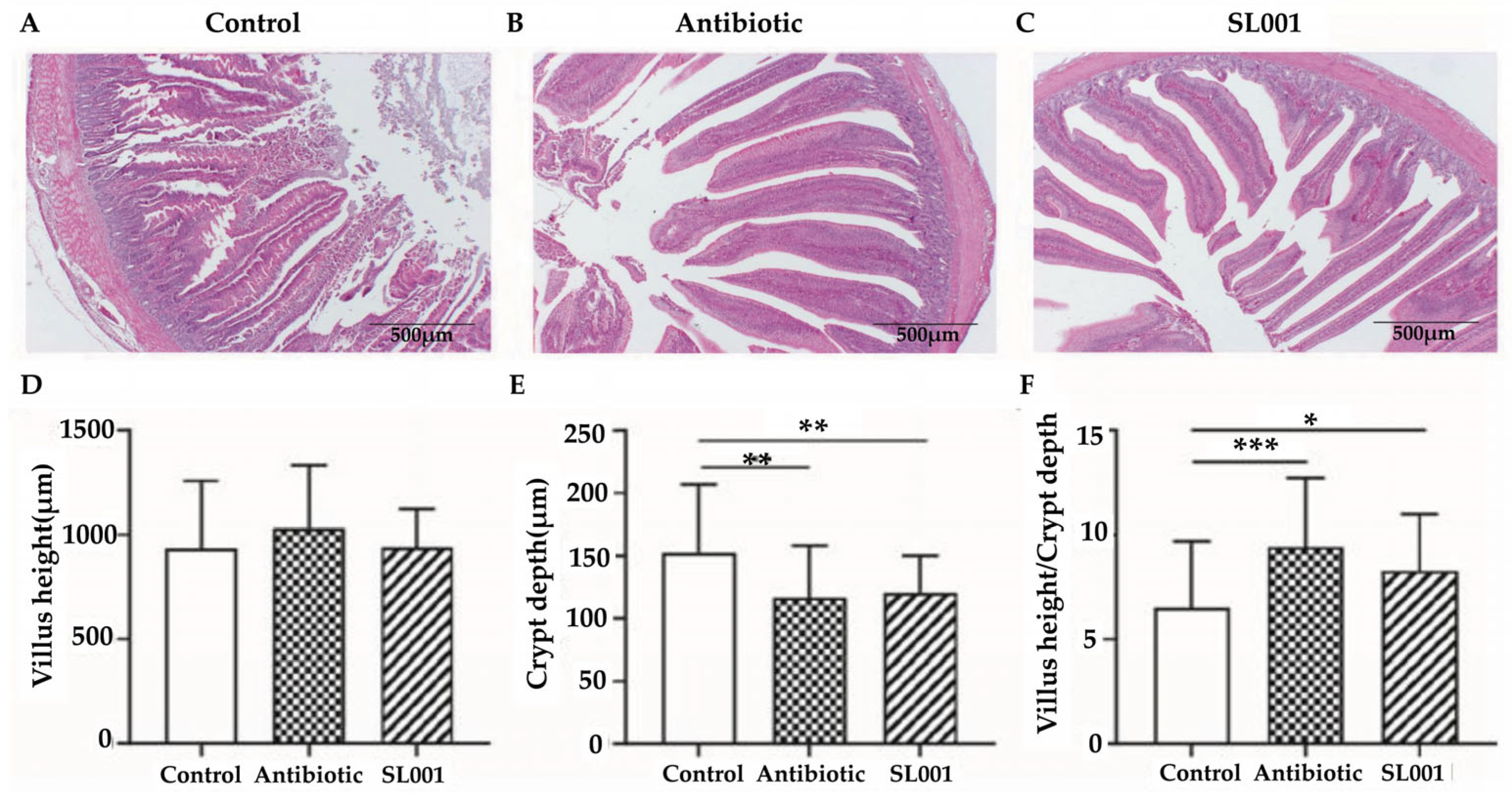
3.4. Intestinal Morphology
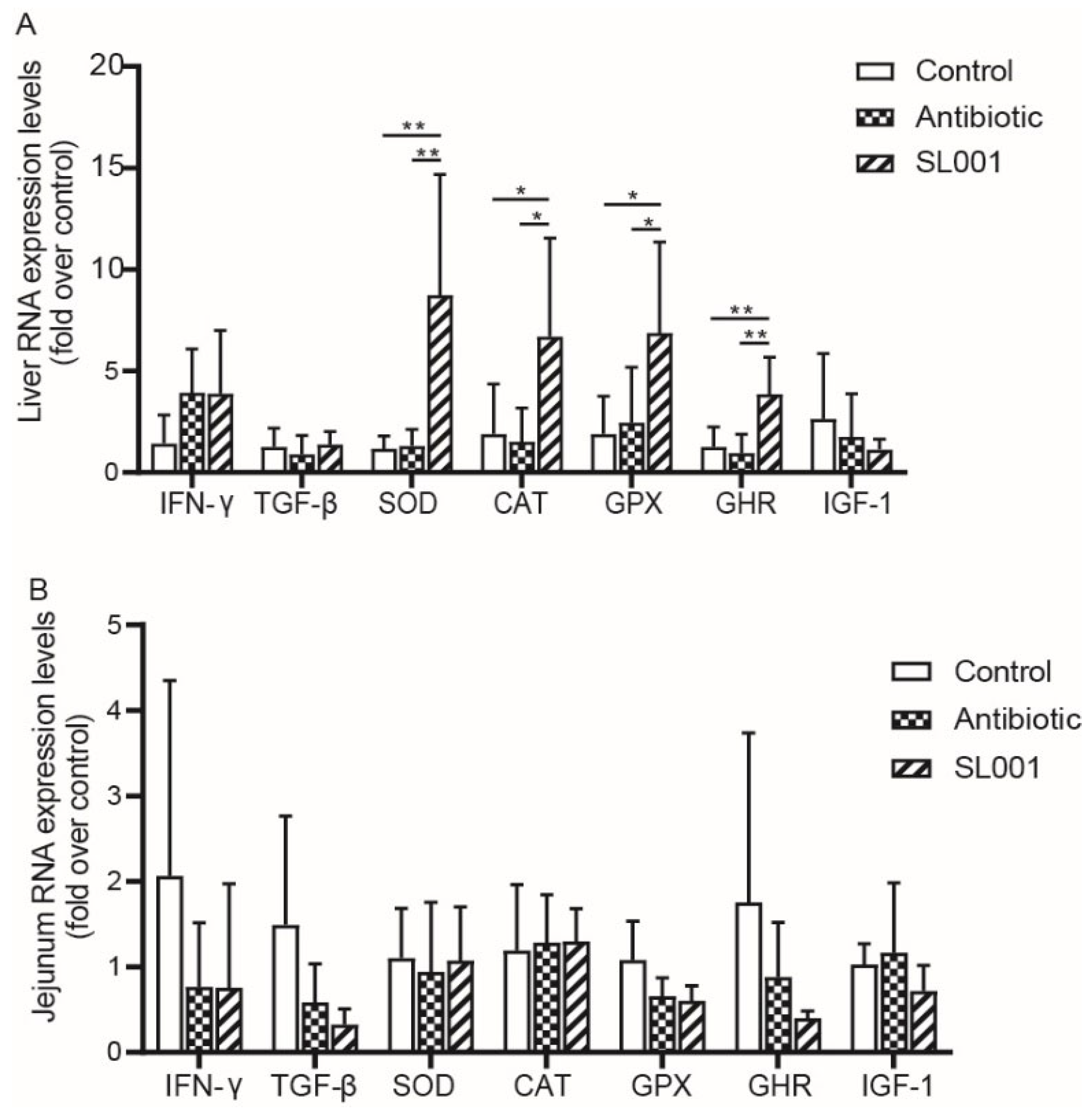
3.5. Gene Expression Levels in Liver and Jejunum
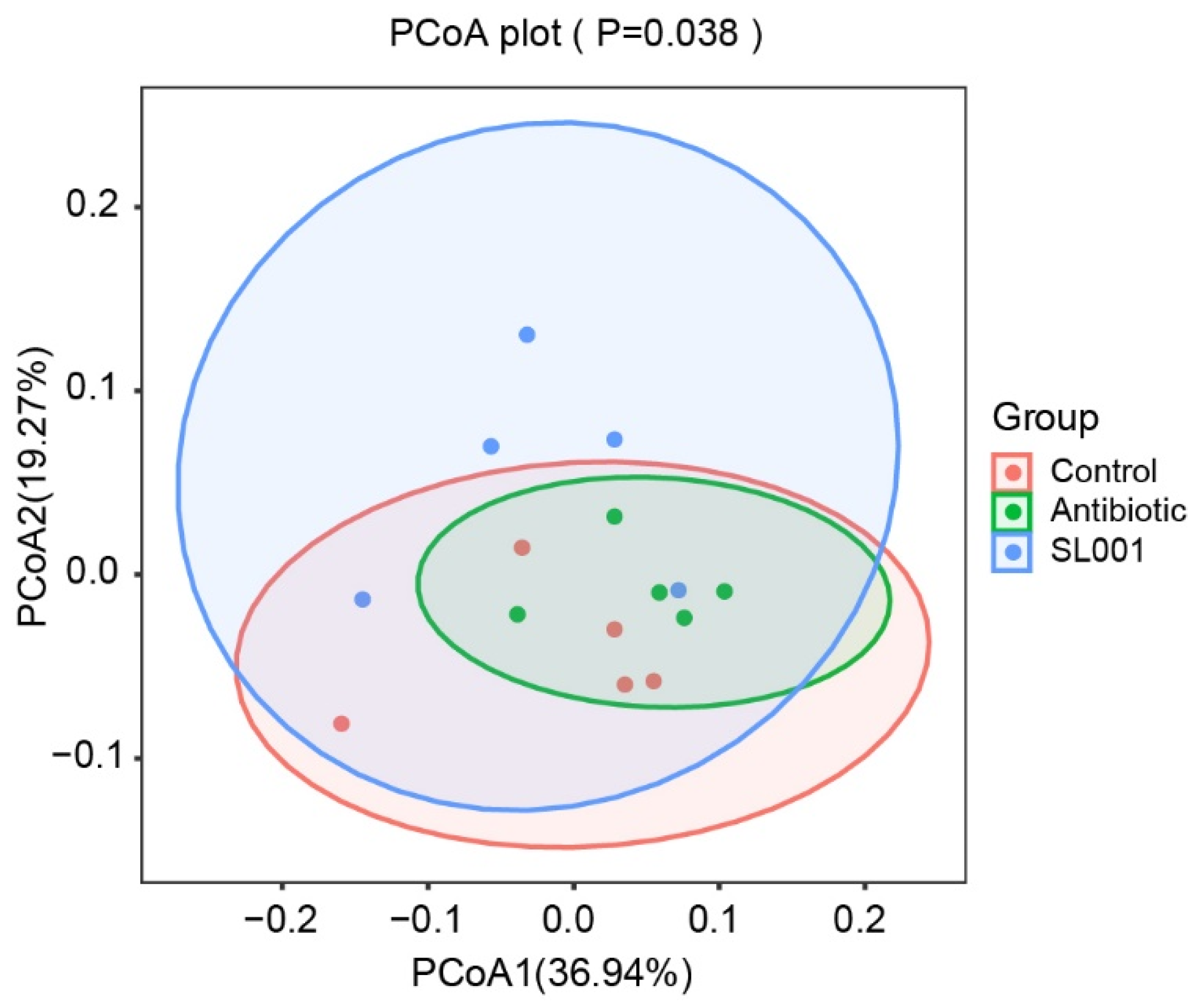
3.6. Microbial Diversity and the Relative Abundance of Intestinal Microbiome
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Roth, N.; Käsbohrer, A.; Mayrhofer, S.; Zitz, U.; Hofacre, C.; Domig, K.J. The application of antibiotics in broiler production and the resulting antibiotic resistance in Escherichia coli: A global overview. Poult. Sci. 2019, 98, 1791–1804. [Google Scholar] [CrossRef]
- Pfaller, M.A. Flavophospholipol use in animals: Positive implications for antimicrobial resistance based on its microbiologic properties. Diagn. Microbiol. Infect. Dis. 2006, 56, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Mehdi, Y.; Létourneau-Montminy, M.-P.; Gaucher, M.-L.; Chorfi, Y.; Suresh, G.; Rouissi, T.; Brar, S.K.; Côté, C.; Ramirez, A.A.; Godbout, S. Use of antibiotics in broiler production: Global impacts and alternatives. Anim. Nutr. 2018, 4, 170–178. [Google Scholar] [CrossRef] [PubMed]
- Berendsen, B.J.A.; Lahr, J.; Nibbeling, C.; Jansen, L.J.M.; Bongers, I.E.A.; Wipfler, E.L.; van de Schans, M.G.M. The persistence of a broad range of antibiotics during calve, pig and broiler manure storage. Chemosphere 2018, 204, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Shang, Y.; Kumar, S.; Oakley, B.; Kim, W.K. Chicken Gut Microbiota: Importance and Detection Technology. Front. Vet. Sci. 2018, 5, 254. [Google Scholar] [CrossRef]
- Kogut, M.H. The effect of microbiome modulation on the intestinal health of poultry. Anim. Feed. Sci. Technol. 2018, 250, 32–40. [Google Scholar] [CrossRef]
- Stanley, D.; Hughes, R.J.; Moore, R.J. Microbiota of the Chicken Gastrointestinal Tract: Influence on Health, Productivity and Disease. Appl. Microbiol. Biotechnol. 2014, 98, 4301–4310. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Jahromi, M.F.; Liang, J.B.; Sieo, C.C.; Kalavathy, R.; Idrus, Z.; Ho, Y.W. In Vitro Assessment of Bioactivities of Lactobacillus Strains as Potential Probiotics for Humans and Chickens. J. Food Sci. 2017, 82, 2734–2745. [Google Scholar] [CrossRef]
- Gonmei, G.; Sapcota, D.; Saikia, G.K.; Deka, P.; Mahanta, J.D.; Kalita, N.; Saikia, B.N.; Talukdar, J.K. Studies on immune response to Newcastle disease virus in broiler chickens fed with Lactobacillus reuteri PIA16 isolated from the gut of indigenous chicken of Assam, India. Vet. World 2019, 12, 1251–1255. [Google Scholar] [CrossRef]
- Chen, C.; Li, J.; Zhang, H.; Xie, Y.; Xiong, L.; Liu, H.; Wang, F. Effects of a probiotic on the growth performance, intestinal flora, and immune function of chicks infected with Salmonella pullorum. Poult. Sci. 2020, 99, 5316–5323. [Google Scholar] [CrossRef]
- Kazemi, S.A.; Ahmadi, H.; Torshizi, M.A.K. Evaluating two multistrain probiotics on growth performance, intestinal morphology, lipid oxidation and ileal microflora in chickens. J. Anim. Physiol. Anim. Nutr. 2019, 103, 1399–1407. [Google Scholar] [CrossRef]
- Liu, X.; Yan, H.; Lv, L.; Xu, Q.; Yin, C.; Zhang, K.; Wang, P.; Hu, J. Growth Performance and Meat Quality of Broiler Chickens Supplemented with Bacillus licheniformis in Drinking Water. Asian-Australasian J. Anim. Sci. 2012, 25, 682–689. [Google Scholar] [CrossRef]
- Huyghebaert, G.; Ducatelle, R.; Van Immerseel, F. An update on alternatives to antimicrobial growth promoters for broilers. Vet. J. 2011, 187, 182–188. [Google Scholar] [CrossRef]
- Vieco-Saiz, N.; Belguesmia, Y.; Raspoet, R.; Auclair, E.; Gancel, F.; Kempf, I.; Drider, D. Benefits and Inputs from Lactic Acid Bacteria and Their Bacteriocins as Alternatives to Antibiotic Growth Promoters during Food-Animal Production. Front. Microbiol. 2019, 10, 57. [Google Scholar] [CrossRef]
- Wang, L.; Liu, C.; Chen, M.; Ya, T.; Huang, W.; Gao, P.; Zhang, H. A novel Lactobacillus plantarum strain P-8 activates beneficial immune response of broiler chickens. Int. Immunopharmacol. 2015, 29, 901–907. [Google Scholar] [CrossRef]
- Shokryazdan, P.; Jahromi, M.F.; Liang, J.B.; Ramasamy, K.; Sieo, C.C.; Ho, Y.W. Effects of a Lactobacillus salivarius mixture on performance, intestinal health and serum lipids of broiler chickens. PLoS ONE 2017, 12, e0175959. [Google Scholar] [CrossRef] [PubMed]
- Liu, M.H.R.; Guo, Y.; Sun, W.; Li, J.; Fan, M.; Wang, Y.; Du, H.; Tang, Z.; Chai, C. Influence of Lactobacillus reuteri SL001 on intestinal microbiota in AD model mice and C57BL/6 mice. Chin. J. Biotechnol. 2020, 36, 1887–1900. [Google Scholar]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of General 16S Ribosomal RNA Gene PCR Primers for Classical and Next-Generation Sequencing-Based Diversity Studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [PubMed]
- Khalique, A.; Zeng, D.; Shoaib, M.; Wang, H.; Qing, X.; Rajput, D.S.; Pan, K.; Ni, X. Probiotics mitigating subclinical necrotic enteritis (SNE) as potential alternatives to antibiotics in poultry. AMB Express 2020, 10, 50. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Yang, Y.; Dong, Y.; Ito, K.; Zhang, B. Highly nutritious diet resists Salmonella typhimurium infections by improving intestinal microbiota and morphology in broiler chickens. Poult. Sci. 2020, 99, 7055–7065. [Google Scholar] [CrossRef]
- Jin, L.Z.; Ho, Y.W.; Abdullah, N.; Jalaludin, S. Digestive and Bacterial Enzyme Activities in Broilers Fed Diets Supplemented with Lactobacillus Cultures. Poult. Sci. 2000, 79, 886–891. [Google Scholar] [CrossRef] [PubMed]
- Apata, D. Growth performance, nutrient digestibility and immune response of broiler chicks fed diets supplemented with a culture of Lactobacillus bulgaricus. J. Sci. Food Agric. 2008, 88, 1253–1258. [Google Scholar] [CrossRef]
- Salehizadeh, M.; Modarressi, M.H.; Mousavi, S.N.; Ebrahimi, M.T. Effects of probiotic lactic acid bacteria on growth performance, carcass characteristics, hematological indices, humoral immunity, and IGF-I gene expression in broiler chicken. Trop. Anim. Health Prod. 2019, 51, 2279–2286. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Gong, L.; Zhou, Y.; Tang, L.; Zeng, Z.; Wang, Q.; Zou, P.; Yu, D.; Li, W. Probiotic Paenibacillus polymyxa 10 and Lactobacillus plantarum 16 enhance growth performance of broilers by improving the intestinal health. Anim. Nutr. 2021, 7, 829–840. [Google Scholar] [CrossRef]
- Neveling, D.P.; Dicks, L.M. Probiotics: An Antibiotic Replacement Strategy for Healthy Broilers and Productive Rearing. Probiotics Antimicrob. Proteins 2020, 13, 1–11. [Google Scholar] [CrossRef]
- Zhang, L.; Lin, S.; Fengfang, Q.; Ma, J.; Qiu, F.; Jia, R.; Nie, Q.; Zhang, D.; Luo, Q.; Lilong, A.; et al. Chicken GHR natural antisense transcript regulates GHR mRNA in LMH cells. Oncotarget 2016, 7, 73607–73617. [Google Scholar] [CrossRef]
- Olnood, C.G.; Beski, S.S.; Choct, M.; Iji, P.A. Novel probiotics: Their effects on growth performance, gut development, microbial community and activity of broiler chickens. Anim. Nutr. 2015, 1, 184–191. [Google Scholar] [CrossRef]
- Korver, D.R.; Klasing, K.C. Dietary Fish Oil Alters Specific and Inflammatory Immune Responses in Chicks. J. Nutr. 1997, 127, 2039–2046. [Google Scholar] [CrossRef]
- Hardy, H.; Harris, J.; Lyon, E.; Beal, J.; Foey, A.D. Probiotics, Prebiotics and Immunomodulation of Gut Mucosal Defences: Homeostasis and Immunopathology. Nutrients 2013, 5, 1869–1912. [Google Scholar] [CrossRef]
- Ren, Z.; Yan, J.; Whelan, R.; Liao, X.; Bütz, D.E.; Arendt, M.K.; Cook, M.E.; Yang, X.; Crenshaw, T.D. Dietary supplementation of sulfur amino acids improves intestinal immunity to Eimeria in broilers treated with anti-interleukin-10 antibody. Anim. Nutr. 2022, 10, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Wan, G.; Peng, C.; Xu, L.; Yu, Y.; Li, L.; Li, G. Effect of probiotic supplementation on growth performance, intestinal morphology, barrier integrity, and inflammatory response in broilers subjected to cyclic heat stress. Anim. Sci. J. Nihon Chikusan Gakkaiho 2020, 91, e13433. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.F.; Kim, I.H. Effects of multistrain probiotics on growth performance, apparent ileal nutrient digestibility, blood characteristics, cecal microbial shedding, and excreta odor contents in broilers. Poult. Sci. 2014, 93, 364–370. [Google Scholar] [CrossRef]
- Han, J.; Wang, Y.; Song, D.; Lu, Z.; Dong, Z.; Miao, H.; Wang, W.; He, J.; Li, A. Effects of Clostridium butyricum and Lactobacillus plantarum on growth performance, immune function and volatile fatty acid level of caecal digesta in broilers. Food Agric. Immunol. 2018, 29, 797–807. [Google Scholar] [CrossRef]
- Wu, Y.; Zhen, W.; Geng, Y.; Wang, Z.; Guo, Y. Effects of dietary Enterococcus faecium NCIMB 11181 supplementation on growth performance and cellular and humoral immune responses in broiler chickens. Poult. Sci. 2019, 98, 150–163. [Google Scholar] [CrossRef]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef]
- Bai, K.; Huang, Q.; Zhang, J.; He, J.; Zhang, L.; Wang, T. Supplemental effects of probiotic Bacillus subtilis fmbJ on growth performance, antioxidant capacity, and meat quality of broiler chickens. Poult. Sci. 2017, 96, 74–82. [Google Scholar] [CrossRef] [PubMed]
- Wang, A.; Yi, X.; Yu, H.; Dong, B.; Qiao, S. Free radical scavenging activity of Lactobacillus fermentum in vitro and its antioxidative effect on growing-finishing pigs. J. Appl. Microbiol. 2009, 107, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Noureen, S.; Riaz, A.; Arshad, M.; Arshad, N. In vitro selection and in vivo confirmation of the antioxidant ability of Lactobacillus brevis MG 000874. J. Appl. Microbiol. 2018, 126, 1221–1232. [Google Scholar] [CrossRef]
- Zhang, Y.; Du, R.; Wang, L.; Zhang, H. The antioxidative effects of probiotic Lactobacillus casei Zhang on the hyperlipidemic rats. Eur. Food Res. Technol. 2010, 231, 151–158. [Google Scholar] [CrossRef]
- Zhang, L.; Bai, K.; Zhang, J.; Xu, W.; Huang, Q.; Wang, T. Dietary effects of Bacillus subtilis fmbj on the antioxidant capacity of broilers at an early age. Poult. Sci. 2017, 96, 3564–3573. [Google Scholar] [CrossRef] [PubMed]
- He, T.; Long, S.; Mahfuz, S.; Wu, D.; Wang, X.; Wei, X.; Piao, X. Effects of Probiotics as Antibiotics Substitutes on Growth Performance, Serum Biochemical Parameters, Intestinal Morphology, and Barrier Function of Broilers. Animals 2019, 9, 985. [Google Scholar] [CrossRef] [PubMed]
- Gong, L.; Wang, B.; Mei, X.; Xu, H.; Qin, Y.; Li, W.; Zhou, Y. Effects of three probiotic Bacillus on growth performance, digestive enzyme activities, antioxidative capacity, serum immunity, and biochemical parameters in broilers. Anim. Sci. J. Nihon Chikusan Gakkaiho 2018, 89, 1561–1571. [Google Scholar] [CrossRef]
- Dev, K.; Mir, N.A.; Biswas, A.; Kannoujia, J.; Begum, J.; Kant, R.; Mandal, A. Dietary synbiotic supplementation improves the growth performance, body antioxidant pool, serum biochemistry, meat quality, and lipid oxidative stability in broiler chickens. Anim. Nutr. 2020, 6, 325–332. [Google Scholar] [CrossRef] [PubMed]
- Żbikowski, A.; Pawłowski, K.; Śliżewska, K.; Dolka, B.; Nerc, J.; Szeleszczuk, P. Comparative Effects of Using New Multi-Strain Synbiotics on Chicken Growth Performance, Hematology, Serum Biochemistry and Immunity. Animals 2020, 10, 1555. [Google Scholar] [CrossRef]
- Harr, K.E. Clinical Chemistry of Companion Avian Species: A Review. Vet. Clin. Pathol. 2002, 31, 140–151. [Google Scholar] [CrossRef]
- Hussein, E.; Selim, S. Efficacy of yeast and multi-strain probiotic alone or in combination on growth performance, carcass traits, blood biochemical constituents, and meat quality of broiler chickens. Livest. Sci. 2018, 216, 153–159. [Google Scholar] [CrossRef]
- Li, T.; Xing, G.; Shao, Y.; Zhang, L.; Li, S.; Lu, L.; Liu, Z.; Liao, X.; Luo, X. Dietary calcium or phosphorus deficiency impairs the bone development by regulating related calcium or phosphorus metabolic utilization parameters of broilers. Poult. Sci. 2020, 99, 3207–3214. [Google Scholar] [CrossRef]
- Zhu, Y.; Wen, J.; Jiang, X.; Wang, W.; Yang, L. High calcium to phosphorus ratio impairs growth and bone mineralization in Pekin ducklings. Poult. Sci. 2018, 97, 1163–1169. [Google Scholar] [CrossRef]
- Khare, A.; Gaur, S. Cholesterol-Lowering Effects of Lactobacillus Species. Curr. Microbiol. 2020, 77, 638–644. [Google Scholar] [CrossRef] [PubMed]
- Alaqil, A.A.; Abbas, A.O.; El-Beltagi, H.S.; El-Atty, H.K.A.; Mehaisen, G.M.K.; Moustafa, E.S. Dietary Supplementation of Probiotic Lactobacillus acidophilus Modulates Cholesterol Levels, Immune Response, and Productive Performance of Laying Hens. Animals 2020, 10, 1588. [Google Scholar] [CrossRef] [PubMed]
- Gheorghe, A.; Lefter, N.A.; Idriceanu, L.; Ropotă, M.; Hăbeanu, M. Effects of dietary extruded linseed and Lactobacillus acidophilus on growth performance, carcass traits, plasma lipoprotein response, and caecal bacterial populations in broiler chicks. Ital. J. Anim. Sci. 2020, 19, 822–832. [Google Scholar] [CrossRef]
- Jiang, J.; Liu, S.; Jamal, T.; Ding, T.; Qi, L.; Lv, Z.; Yu, D.; Shi, F. Effects of dietary sweeteners supplementation on growth performance, serum biochemicals, and jejunal physiological functions of broiler chickens. Poult. Sci. 2020, 99, 3948–3958. [Google Scholar] [CrossRef]
- Xu, L.; Sun, X.; Wan, X.; Li, K.; Jian, F.; Li, W.; Jiang, R.; Han, R.; Li, H.; Kang, X.; et al. Dietary supplementation with Clostridium butyricum improves growth performance of broilers by regulating intestinal microbiota and mucosal epithelial cells. Anim. Nutr. 2021, 7, 1105–1114. [Google Scholar] [CrossRef] [PubMed]
- Abramowicz, K.; Krauze, M.; Ognik, K. The Effect of a Probiotic Preparation Containing Bacillus subtilis PB6 in the Diet of Chickens on Redox and Biochemical Parameters in Their Blood. Ann. Anim. Sci. 2019, 19, 433–451. [Google Scholar] [CrossRef]
- Wang, Y.; Gu, Q. Effect of probiotic on growth performance and digestive enzyme activity of Arbor Acres broilers. Res. Vet. Sci. 2010, 89, 163–167. [Google Scholar] [CrossRef]
- Ducatelle, R.; Goossens, E.; De Meyer, F.; Eeckhaut, V.; Antonissen, G.; Haesebrouck, F.; Van Immerseel, F. Biomarkers for monitoring intestinal health in poultry: Present status and future perspectives. Vet. Res. 2018, 49, 43. [Google Scholar] [CrossRef]
- Rodrigues, D.R.; Briggs, W.; Duff, A.; Chasser, K.; Murugesan, R.; Pender, C.; Ramirez, S.; Valenzuela, L.; Bielke, L.R. Comparative effectiveness of probiotic-based formulations on cecal microbiota modulation in broilers. PLoS ONE 2020, 15, e0225871. [Google Scholar] [CrossRef]
- Ma, Y.; Wang, W.; Zhang, H.; Wang, J.; Zhang, W.; Gao, J.; Wu, S.; Qi, G. Supplemental Bacillus subtilis DSM 32315 manipulates intestinal structure and microbial composition in broiler chickens. Sci. Rep. 2018, 8, 15358. [Google Scholar] [CrossRef]
- Wang, Y.; Sun, J.; Zhong, H.; Li, N.; Xu, H.; Zhu, Q.; Liu, Y. Effect of probiotics on the meat flavour and gut microbiota of chicken. Sci. Rep. 2017, 7, 6400. [Google Scholar] [CrossRef] [PubMed]
- Barka, E.A.; Vatsa, P.; Sanchez, L.; Gaveau-Vaillant, N.; Jacquard, C.; Klenk, H.-P.; Clément, C.; Ouhdouch, Y.; van Wezel, G.P. Taxonomy, Physiology, and Natural Products of Actinobacteria. Microbiol. Mol. Biol. Rev. 2016, 80, 1–43. [Google Scholar] [CrossRef] [PubMed]
- Tabashsum, Z.; Peng, M.; Alvarado-Martinez, Z.; Aditya, A.; Bhatti, J.; Romo, P.B.; Young, A.; Biswas, D. Competitive reduction of poultry-borne enteric bacterial pathogens in chicken gut with bioactive Lactobacillus casei. Sci. Rep. 2020, 10, 16259. [Google Scholar] [CrossRef] [PubMed]
- Dubin, K.; Callahan, M.K.; Ren, B.; Khanin, R.; Viale, A.; Ling, L.; No, D.; Gobourne, A.; Littmann, E.; Huttenhower, C.; et al. Intestinal microbiome analyses identify melanoma patients at risk for checkpoint-blockade-induced colitis. Nat. Commun. 2016, 7, 10391. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | 1 to 21 d | 22 to 42 d |
|---|---|---|
| Ingredient (%) | ||
| Corn | 58.00 | 63.00 |
| Soybean (44%) | 28.20 | 24.90 |
| Gluten (60%) | 7.00 | 6.00 |
| Dicalcium phosphate | 1.80 | 1.20 |
| Limestone | 1.30 | 1.30 |
| Oil soybean | 2.00 | 2.00 |
| Salts | 0.30 | 0.30 |
| L-Lys-HCL | 0.20 | 0.20 |
| DL-Met | 0.20 | 0.10 |
| Premix 1 | 1.00 | 1.00 |
| Total | 100.00 | 100.00 |
| Nutritional ingredient 2 | ||
| Energy (MJ/kg) | 12.60 | 12.84 |
| Crude protein % | 21.99 | 20.28 |
| Calcium % | 0.98 | 0.84 |
| Total phosphor % | 0.72 | 0.59 |
| Methionine % | 0.59 | 0.47 |
| Methionine + Cysteine % | 0.95 | 0.80 |
| Lysine % | 1.14 | 1.06 |
| Gene | Primer Sequence (5′-3′) | Accession Number |
|---|---|---|
| IFN-γ | Forward: AACGCCAAAGCCTCCTCAAC Reverse: TGAGGTGAAGGTTGCGAGGC | NM_205427.1 |
| TGF-β | Forward: CGGGACGGATGAGAAGAAC Reverse: CGGCCCACGTAGTAAATGAT | NM_205454.1 |
| SOD | Forward: GCACGGTGGACCAAAAGA Reverse: AACGAGGTCCAGCATTTCC | NM_205064.1 |
| CAT | Forward: GGTTCGGTGGGGTTGTCTTT Reverse: CACCAGTGGTCAAGGCATCT | NM_001031215.2 |
| GPX | Forward: CAGGAGAACGCCACCAACG Reverse: TCTCAGGAAGGCGAACAGC | NM_001277853.1 |
| GHR | Forward: AACACAGATACCCAACAGCC Reverse: AGAAGTCAGTGTTTGTCAGGG | NM_001001293.1 |
| IGF-1 | Forward: CATTCATTTCTTCTACCTTGGC Reverse: AGCAGCATTCATCCACTATTCC | NM_001004384.2 |
| β-actin | Forward: GAGAAATTGTGCGTGACATCA Reverse: CCTGAACCTCTCATTGCCA | NM_205518.1 |
| Items | Control | Antibiotic | SL001 | p-Value |
|---|---|---|---|---|
| BW (g) | ||||
| Initial | 47.34 ± 0.39 | 47.36 ± 0.57 | 47.74 ± 0.26 | 0.799 |
| 3 weeks | 650.26 ± 8.25 ab | 635.82 ± 7.56 b | 676.42 ± 14.89 a | 0.051 |
| 6 weeks | 1644.41 ± 40.85 b | 1607.17 ± 30.39 b | 1780.68 ± 34.69 a | 0.009 |
| ADG (g/day) | ||||
| 0–3 weeks | 28.71 ± 0.38 ab | 28.02 ± 0.36 b | 29.94 ± 0.72 a | 0.055 |
| 4–6 weeks | 46.98 ± 1.55 b | 46.37 ± 1.45 b | 52.78 ± 1.46 a | 0.015 |
| 0–6 weeks | 37.85 ± 0.87 b | 37.20 ± 10.732 b | 41.36 ± 2.16 a | 0.006 |
| ADFI (g/day) | ||||
| 0–3 weeks | 44.14 ± 1.15 | 43.02 ± 0.97 | 43.94 ± 1.77 | 0.823 |
| 4–6 weeks | 92.06 ± 2.34 c | 107.12 ± 1.87 a | 100.00 ± 2.36 b | <0.001 |
| 0–6 weeks | 68.10 ± 1.53 b | 75.07 ± 1.08 a | 71.97 ± 1.93 ab | 0.021 |
| F/G (g/g) | ||||
| 0–3 weeks | 1.54 ± 0.25 | 1.54 ± 0.53 | 1.47 ± 0.07 | 0.624 |
| 4–6 weeks | 1.96 ± 0.07 b | 2.32 ± 0.06 a | 1.90 ± 0.08 b | 0.002 |
| 0–6 weeks | 1.80 ± 0.04 b | 2.02 ± 0.04 a | 1.75 ± 0.06 b | 0.004 |
| Items (g/g) | Control | Antibiotic | SL001 | p-Value |
|---|---|---|---|---|
| Carcass | 79.08 ± 0.76 ab | 76.36 ± 2.08 b | 81.87 ± 0.40 a | 0.019 |
| Breast | 18.15 ± 0.50 | 17.49 ± 0.41 | 18.73 ± 0.51 | 0.199 |
| Abdominal fat | 1.68 ± 0.10 | 1.62 ± 0.10 | 1.80 ± 0.15 | 0.560 |
| Liver | 2.42 ± 0.10 | 2.39 ± 0.11 | 2.23 ± 0.17 | 0.580 |
| Gizzard | 2.18 ± 0.22 a | 2.04 ± 0.10 a | 1.44 ± 0.1 b | 0.004 |
| Bursa | 0.20 ± 0.02 | 0.21 ± 0.02 | 0.22 ± 0.02 | 0.613 |
| Spleen | 0.14 ± 0.03 | 0.10 ± 0.01 | 0.13 ± 0.01 | 0.308 |
| Heart | 0.45 ± 0.02 b | 0.46 ± 0.02 b | 0.52 ± 0.03 a | 0.050 |
| Stomach | 0.41 ± 0.02 | 0.42 ± 0.02 | 0.43 ± 0.02 | 0.915 |
| Pancreas | 0.19 ± 0.01 | 0.19 ± 0.01 | 0.20 ± 0.01 | 0.906 |
| Items | Control | Antibiotic | SL001 | p-Value |
|---|---|---|---|---|
| IL-6 (pg/mL) | 29.34 ± 1.20 a | 16.57 ± 1.44 b | 18.58 ± 0.49 b | <0.001 |
| IL-4 (pg/mL) | 159.88 ± 2.87 a | 103.42 ± 5.00 b | 102.49 ± 5.60 b | <0.001 |
| IL-10 (pg/mL) | 36.13 ± 2.41 b | 63.52 ± 1.52 a | 57.72 ± 2.85 a | <0.001 |
| TNF-α (pg/mL) | 74.67 ± 3.14 a | 50.96 ± 2.94 b | 54.85 ± 1.21 b | <0.001 |
| TGF-β (pg/mL) | 134.32 ± 4.71 b | 195.92 ± 3.65 a | 184.12 ± 9.04 a | <0.001 |
| IgM (μg/mL) | 895.42 ± 25.63 b | 1223.36 ± 29.51 a | 1143.87 ± 26.70 a | <0.001 |
| IgA (μg/mL) | 514.04 ± 12.70 b | 709.08 ± 23.94 a | 644.02 ± 5.81 a | <0.001 |
| IgG (μg/mL) | 1776.63 ± 78.83 c | 2787.04 ± 65.58 a | 2339.49 ± 68.89 b | <0.001 |
| IgE (μg/mL) | 15.94 ± 0.84 c | 22.56 ± 0.67 a | 20.18 ± 0.33 b | <0.001 |
| sIgA (ng/mL) | 1492.51 ± 63.28 b | 2166.70 ± 142.63 a | 2172.14 ± 99.97 a | <0.001 |
| C3 (μg/mL) | 594.37 ± 12.46 b | 769.84 ± 38.57 a | 709.93 ± 41.41 ab | 0.007 |
| C4 (μg/mL) | 136.39 ± 7.08 b | 192.21 ± 12.77 a | 208.21 ± 8.71 a | <0.001 |
| Items | Control | Antibiotic | SL001 | p-Value |
|---|---|---|---|---|
| ALB (g/L) | 24.19 ± 1.03 b | 37.37 ± 2.24 a | 35.31 ± 1.72 a | <0.001 |
| TP (g/L) | 49.72 ± 1.25 b | 66.89 ± 1.78 a | 63.53 ± 1.95 a | <0.001 |
| Cr (μmol/L) | 265.71 ± 16.19 a | 186.56 ± 6.24 b | 218.13 ± 2.52 a | <0.001 |
| UA (μmol/L) | 255.97 ± 5.83 a | 191.54 ± 8.00 b | 208.16 ± 6.49 b | <0.001 |
| AKP (U/L) | 60.77 ± 1.49 b | 89.14 ± 2.52 a | 89.17 ± 3.31 a | <0.001 |
| GPT (U/L) | 45.23 ± 6.17 b | 57.40 ± 2.15 a | 56.81 ± 2.20 ab | 0.082 |
| GOT (U/L) | 70.17 ± 5.10 b | 82.24 ± 2.54 ab | 83.49 ± 4.62 a | 0.080 |
| AMS (U/dL) | 151.00 ± 8.97 b | 278.45 ± 19.50 a | 254.95 ± 19.53 a | <0.001 |
| LPS (U/L) | 593.40 ± 43.50 b | 954.49 ± 26.40 a | 864.28 ± 36.35 a | <0.001 |
| PEP (U/mL) | 46.05 ± 1.25 b | 69.02 ± 4.38 a | 59.41 ± 5.48 ab | 0.005 |
| TCHO (mmol/L) | 4.68 ± 0.19 a | 2.87 ± 0.18 c | 3.44 ± 0.10 b | <0.001 |
| TG (mmol/L) | 1.88 ± 0.05 a | 1.28 ± 0.06 b | 1.37 ± 0.07 b | <0.001 |
| VLDL (mmol/L) | 14.01 ± 0.53 a | 7.90 ± 0.48 b | 7.41 ± 0.50 b | <0.001 |
| LDL (mmol/L) | 9.99 ± 0.26 a | 6.08 ± 0.39 c | 7.71 ± 0.17 b | <0.001 |
| HDL (mg/dL) | 110.30 ± 5.79 c | 164.16 ± 4.89 a | 143.23 ± 4.14 b | <0.001 |
| Items | Control | Antibiotic | SL001 | p-Value |
|---|---|---|---|---|
| Observed_OTUs | 567.20 ± 31.33 | 581.40 ± 15.92 | 592.40 ± 20.68 | 0.755 |
| Chao1 | 569.23 ± 31.73 | 583.76 ± 16.39 | 592.92 ± 20.57 | 0.781 |
| Shannon | 7.00 ± 0.09 | 7.06 ± 0.10 | 7.12 ± 0.12 | 0.739 |
| Simpson | 0.978 ± 0.004 | 0.976 ± 0.002 | 0.980 ± 0.002 | 0.679 |
| Items | Control | Antibiotic | SL001 | p-Value |
|---|---|---|---|---|
| Firmicutes | 71.71 ± 4.18 | 78.98 ± 2.20 | 72.53 ± 3.77 | 0.308 |
| Bacteroidetes | 22.16 ± 3.45 | 16.23 ± 1.94 | 20.50 ± 3.13 | 0.364 |
| Proteobacteria | 4.35 ± 0.96 | 3.35 ± 0.48 | 3.00 ± 0.41 | 0.357 |
| Tenericutes | 0.94 ± 0.22 | 0.88 ± 0.23 | 1.57 ± 0.25 | 0.112 |
| Verrucomicrobia | 0.51 ± 0.22 | 0.30 ± 0.14 | 1.70 ± 0.72 | 0.092 |
| Lentisphaerae | 0.19 ± 0.12 | 0.11 ± 0.05 | 0.07 ± 0.02 | 0.535 |
| Actinobacteria | 0.042 ± 0.008 b | 0.078 ± 0.017 b | 0.21 ± 0.042 a | 0.002 |
| Chloroflexi | 0.002 ± 0.002 b | 0.000 ± 0.000 b | 0.208 ± 0.077 a | 0.008 |
| Acidobacteria | 0.002 ± 0.002 b | 0.000 ± 0.000 b | 0.116 ± 0.046 a | 0.014 |
| Cyanobacteria | 0.076 ± 0.034 | 0.018 ± 0.010 | 0.018 ± 0.006 | 0.109 |
| Genus | Control | Antibiotic | SL001 | p-Value |
|---|---|---|---|---|
| Faecalibacterium | 13.02 ± 1.91 a | 13.12 ± 1.19 a | 7.57 ± 1.72 b | 0.053 |
| Ruminococcaceae_UCG-014 | 6.40 ± 0.67 b | 7.53 ± 0.92 ab | 10.27 ± 1.40 a | 0.058 |
| Alistipes | 6.18 ± 2.08 | 8.48 ± 1.63 | 9.09 ± 1.60 | 0.497 |
| Ruminococcaceae_UCG-005 | 5.08 ± 0.73 | 6.23 ± 1.67 | 6.02 ± 1.72 | 0.839 |
| Parabacteroides | 9.85 ± 1.79 a | 3.30 ± 1.53 b | 3.98 ± 2.01 b | 0.046 |
| Bacteroides | 4.85 ± 1.46 | 3.28 ± 0.72 | 6.22 ± 1.81 | 0.367 |
| Lachnospiraceae_unclassified | 4.21 ± 0.64 | 4.68 ± 0.44 | 3.75 ± 0.51 | 0.493 |
| Clostridiales_vadinBB60_group_unclassified | 3.07 ± 0.42 b | 3.58 ± 0.40 ab | 5.62 ± 1.26 a | 0.100 |
| Intestinimonas | 4.92 ± 0.63 | 3.74 ± 0.88 | 3.61 ± 0.71 | 0.424 |
| Lactobacillus | 2.76 ± 0.63 b | 4.96 ± 0.74 a | 2.76 ± 0.28 b | 0.031 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chai, C.; Guo, Y.; Mohamed, T.; Bumbie, G.Z.; Wang, Y.; Zeng, X.; Zhao, J.; Du, H.; Tang, Z.; Xu, Y.; et al. Dietary Lactobacillus reuteri SL001 Improves Growth Performance, Health-Related Parameters, Intestinal Morphology and Microbiota of Broiler Chickens. Animals 2023, 13, 1690. https://doi.org/10.3390/ani13101690
Chai C, Guo Y, Mohamed T, Bumbie GZ, Wang Y, Zeng X, Zhao J, Du H, Tang Z, Xu Y, et al. Dietary Lactobacillus reuteri SL001 Improves Growth Performance, Health-Related Parameters, Intestinal Morphology and Microbiota of Broiler Chickens. Animals. 2023; 13(10):1690. https://doi.org/10.3390/ani13101690
Chicago/Turabian StyleChai, Chunli, Yaowen Guo, Taha Mohamed, Gifty Z. Bumbie, Yan Wang, Xiaojing Zeng, Jinghua Zhao, Huamao Du, Zhiru Tang, Yetong Xu, and et al. 2023. "Dietary Lactobacillus reuteri SL001 Improves Growth Performance, Health-Related Parameters, Intestinal Morphology and Microbiota of Broiler Chickens" Animals 13, no. 10: 1690. https://doi.org/10.3390/ani13101690
APA StyleChai, C., Guo, Y., Mohamed, T., Bumbie, G. Z., Wang, Y., Zeng, X., Zhao, J., Du, H., Tang, Z., Xu, Y., & Sun, W. (2023). Dietary Lactobacillus reuteri SL001 Improves Growth Performance, Health-Related Parameters, Intestinal Morphology and Microbiota of Broiler Chickens. Animals, 13(10), 1690. https://doi.org/10.3390/ani13101690