
The Timing of the Maternal Recognition of Pregnancy Is Specific to Individual Mares

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mares
2.2. Experimental Protocol
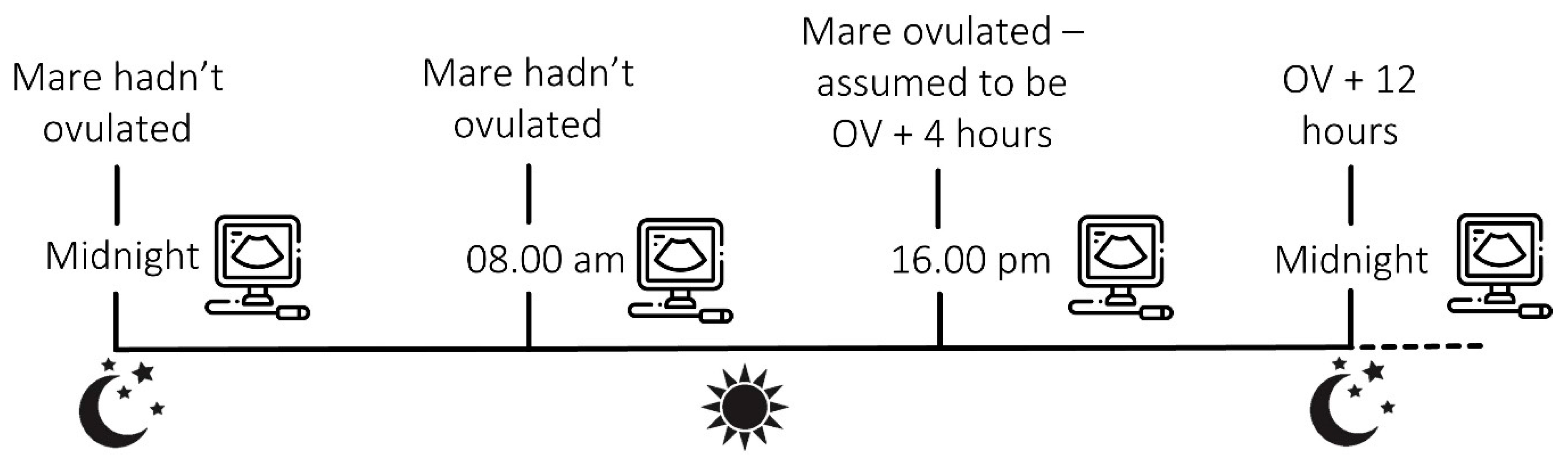
2.2.1. Insemination and Timing of Ovulation
2.2.2. Embryo Reductions
2.2.3. Monitoring Mares for Luteostasis or Luteolysis
2.2.4. Statistical Analysis
3. Results
3.1. The Effect of the Embryo Reduction on Luteostasis
3.2. Embryo Recognition and Vesicle Size
3.3. Twin Embryo Reduction
3.4. Cycle Length and Mare Age
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Short, R.V. Implantation and maternal recognition of pregnancy. In Foetal Autonomy, Ciba Foundation Symposium; Wolstenholme, E.W., O’Connor, M., Eds.; J&A Churchill Ltd.: London, UK, 1969; pp. 2–26. [Google Scholar]
- Bazer, F.W. Pregnancy recognition signalling mechanisms in ruminants and pigs. J. Anim. Sci. Biotech. 2013, 4, 23. [Google Scholar] [CrossRef] [PubMed]
- Douglas, R.H.; Ginther, O.J. Concentration of prostaglandins F in the uterine venous plasma of anaesthetized mares during the oestrous cycle and early pregnancy. Prostaglandins 1976, 11, 251–260. [Google Scholar] [CrossRef] [PubMed]
- Zavy, M.T.; Bazer, F.W.; Sharp, D.C.; Frank, M.; Thatcher, W.W. Uterine luminal prostaglandin F in cycling mares. Prostaglandins 1978, 16, 43–650. [Google Scholar] [CrossRef]
- Vernon, M.W.; Zavy, M.T.; Asquith, R.L.; Sharp, D.C. Prostaglandin F2alpha in the equine endometrium: Steroid modulation and production capacities during the estrous cycle and early pregnancy. Biol. Reprod. 1981, 25, 581–589. [Google Scholar] [CrossRef]
- Berglund, L.A.; Sharp, D.C.; Vernon, M.W.; Thatcher, W.W. Effect of pregnancy and collection technique on prostaglandin F in the uterine lumen of pony mares. J. Reprod. Fert. Suppl. 1982, 32, 335–341. [Google Scholar]
- Stout, T.A.; Allen, W.R. Prostaglandin E(2) and F(2 alpha) production by equine conceptuses and concentrations in conceptus fluids and uterine flushings recovered from early pregnant and dioestrous mares. Reproduction 2002, 123, 261–268. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J.; Domingues, R.R.; Kennedy, V.C.; Dangudubiyyam, S.V. Endogenous and exogenous effects of PGF2α during luteolysis in mares. Theriogenology 2019, 132, 45–52. [Google Scholar] [CrossRef] [PubMed]
- Castro, T.; Jacob, J.C.; Stefani, G.; Domingues, R.R.; Ginther, O.J. Concentrations of progesterone and a PGF2α metabolite during the interovulatory interval compared to the corresponding days of pregnancy in the mare. Theriogenology 2021, 165, 10–17. [Google Scholar] [CrossRef]
- Ababneh, M.M.; Troedsson, M.H. Endometrial phospholipase A2 activity during the oestrous cycle and early pregnancy in mares. Reprod. Domest. Anim. 2013, 48, 46–52. [Google Scholar] [CrossRef]
- Sharp, D.C.; Thatcher, M.J.; Salute, M.E.; Fuchs, A.R. Relationship between endometrial oxytocin receptors and oxytocin-induced prostaglandin F2 alpha release during the oestrous cycle and early pregnancy in pony mares. J. Reprod. Fertil. 1997, 109, 137–144. [Google Scholar] [CrossRef]
- Starbuck, G.R.; Stout, T.A.; Lamming, G.E.; Allen, W.R.; Flint, A.P. Endometrial oxytocin receptor and uterine prostaglandin secretion in mares during the oestrous cycle and early pregnancy. J. Reprod. Fertil. 1998, 113, 173–179. [Google Scholar] [CrossRef] [PubMed]
- Diel de Amorin, M.; Bramer, S.A.; Rajamnickam, G.D.; Klein, C.; Card, C. Endometrial and luteal gene expression of putative gene regulators of equine maternal recognition of pregnancy. Anim. Reprod. Sci. 2002, 245, 107064. [Google Scholar] [CrossRef] [PubMed]
- Klein, C.; Scoggin, K.E.; Ealy, A.D.; Mats, H.T.; Troedsson, M.H.T. Transcriptional profiling of equine endometrium during the time of maternal recognition of pregnancy. Biol. Reprod. 2010, 83, 102–113. [Google Scholar] [CrossRef] [PubMed]
- de Ruijter-Villani, M.; van Tol, H.T.; Stout, T.A. Effect of pregnancy on endometrial expression of luteolytic pathway components in the mare. Reprod. Fertil. Dev. 2015, 27, 834–845. [Google Scholar] [CrossRef] [PubMed]
- Bae, S.E.; Watson, E.D. A light microscopic and ultrastructural study on the presence and location of oxytocin in the equine endometrium. Theriogenology 2003, 60, 909–921. [Google Scholar] [CrossRef]
- Behrendt-Adam, C.Y.; Adams, M.H.; Simpson, K.S.; McDowell, K.J. Oxytocin-neurophysin I mRNA abundance in equine uterine endometrium. Domest. Anim. Endocrinol. 1999, 16, 183–192. [Google Scholar] [CrossRef]
- Diel de Amorin, M.; Dong, L.; Byron, M.; Foster, R.A.; Klein, C.; Saleh, M.; Saleh, T.; Card, C. Characterization of serum and tissue oxytocinase and tissue oxytocin in the pregnant and non-pregnant mare. Sci. Rep. 2023, 13, 4616. [Google Scholar] [CrossRef]
- Stevenson, K.R.; Parkinson, T.J.; Wathes, D.C. Measurement of oxytocin concentrations in plasma and ovarian extracts during the oestrous cycle of mares. J. Reprod. Fertil. 1991, 93, 437–441. [Google Scholar] [CrossRef]
- Stock, A.E.; Emeny, R.T.; Sirois, J.; Fortune, J.E. Oxytocin in mares: Lack of evidence for oxytocin production by or action on preovulatory follicles. Domest. Anim. Endocrinol. 1995, 12, 133–142. [Google Scholar] [CrossRef]
- Flint, A.P.; Sheldrick, E.L. Ovarian oxytocin and the maternal recognition of pregnancy. J. Reprod. Fertil. 1986, 76, 831–839. [Google Scholar] [CrossRef]
- Newcombe, J.R. Embryonic loss and abnormalities of early pregnancy. Equine Vet. Educ. 2000, 12, 88–101. [Google Scholar] [CrossRef]
- Newcombe, J.R. The relationship between the number, diameter, and survival of early embryonic vesicles. Pferdeheilkunde 2004, 20, 214–220. [Google Scholar] [CrossRef]
- Wilsher, S.; Clutton-Brock, A.; Allen, W.R. Successful transfer of day 10 horse embryos: Influence of donor-recipient asynchrony on embryo development. Reproduction 2010, 139, 575–585. [Google Scholar] [CrossRef]
- Boerboom, D.; Brown, K.A.; Vaillancourt, D.; Poitras, P.; Goff, A.K.; Watanabe, K.; Doré, M.; Sirois, J. Expression of key prostaglandin synthases in equine endometrium during late diestrus and early pregnancy. Biol. Reprod. 2004, 70, 391–399. [Google Scholar] [CrossRef] [PubMed]
- Sharp, D.C.; McDowell, K.J.; Weithenauer, J.; Thatcher, W.W. The continuum of events leading to maternal recognition of pregnancy in mares. J. Reprod. Fertil. Suppl. 1989, 37, 101–107. [Google Scholar] [PubMed]
- Wilsher, S.; Newcombe, J.R.; Ismer, A.; Duarte, A.; Kovacsy, S.; Allen, W.R. The effect of embryo reduction and transfer on luteostasis in the mare. Anim. Reprod. Sci. 2022, 242, 107002. [Google Scholar] [CrossRef] [PubMed]
- Wilsher, S.; Ismer, A.; Newcombe, J.; Allen, W.R. Transfer of Day 11 ruptured embryos can prevent luteolysis in recipient mares. J. Equine Vet. Sci. 2022, 113, 103989. [Google Scholar] [CrossRef]
- Newcombe, J.R.; Wilsher, S.; Cuervo-Arango, J. The timing of the maternal recognition of pregnancy signal is specific to individual mares. J. Equine Vet. Sci. 2022, 113, 103977. [Google Scholar] [CrossRef]
- Klein, C. Pregnancy recognition and implantation of the conceptus in the mare. In Regulation of Implantation and Establishment of Pregnancy in Mammals: Tribute to 45 Year Anniversary of Roger V. Short’s “Maternal Recognition of Pregnancy”. Advances in Anatomy, Embryology and Cell Biology; Geisert, R.D., Bazer, F.W., Eds.; Springer International Publishing: Cham, Switzerland, 2015; Volume 216, pp. 165–188. [Google Scholar] [CrossRef]
- Wilsher, S.; Allen, W.R. Uterine influences on embryogenesis and early placentation in the horse revealed by transfer of day-10 embryos to day-3 recipient mares. Reproduction 2009, 137, 583–593. [Google Scholar] [CrossRef]
- Ginther, O.J. Characteristics of the ovulatory season. In Reproductive Biology of the Mare: Basic and Applied Aspects, 1st ed.; University of Wisconsin: Cross Plains, WI, USA, 1979; pp. 133–168. [Google Scholar]
- Hughes, J.P.; Stabenfeldt, G.H.; Evans, J.W. The oestrous cycle of the mare. J. Reprod. Fertil. Suppl. 1975, 23, 161–166. [Google Scholar]
- Hughes, J.P.; Stabenfeldt, G.H.; Evans, J.W. The oestrous cycle of the mare and its uterine control. Aust. Vet. J. 1977, 53, 415–418. [Google Scholar] [CrossRef]
- Heidler, B.; Aurich, J.E.; Pohl, W.; Aurich, C. Bodyweight of mares and foals, estrous cycles and plasma glucose concentration in lactating and non-lactating Lipizzaner mares. Theriogenology 2004, 61, 883–893. [Google Scholar] [CrossRef]
- Cuervo-Arango, J.; Mateu-Sánchez, S.; Aguilar, J.J.; Nielsen, J.M.; Etcharren, V.; Vettorazzi, M.L.; Newcombe, J.R. The effect of the interval from PGF treatment to ovulation on embryo recovery and pregnancy rate in the mare. Theriogenology 2015, 83, 1272–1278. [Google Scholar] [CrossRef]
- Mateu-Sánchez, S.; Newcombe, J.R.; Garcés-Narro, C.; Cuervo-Arango, J. The period of the follicular phase during which the uterus of mares shows estrus-like echotexture influences the subsequent pregnancy rate. Theriogenology 2016, 86, 1506–1515. [Google Scholar] [CrossRef]
- Cuervo-Arango, J.; Claes, A.N.; Ruijter-Villani, M.M.; Stout, T.A. Likelihood of pregnancy after embryo transfer is reduced in recipient mares with a short preceding oestrus. Equine Vet. J. 2018, 50, 386–390. [Google Scholar] [CrossRef]
- Grabowska, A.; Kozdrowski, R. Relationship between estrus endometrial edema and progesterone production in pregnant mares two weeks after ovulation. BMC Vet. Res. 2022, 18, 414. [Google Scholar] [CrossRef]
- Silva, E.S.M.; Cuervo-Arango, J.; de Ruijter-Villani, M.; Klose, K.; Oquendo, P.S.; Stout, T.A.E. Effect of the duration of estradiol priming prior to progesterone administration on endometrial gene expression in anestrous mares. Theriogenology 2019, 131, 96–105. [Google Scholar] [CrossRef]
- Satoh, S.; Hochi, S. A study of reproduction in the mare. II. The oestrous duration and oestrous cycle. J. Jpn. Soc. Vet. Sci. 1933, 12, 200–220. [Google Scholar] [CrossRef]
- Ginther, O.J.; Whitmore, H.L.; Squires, E.L. Characteristics of estrus, diestrus and ovulation in mares and effects of season and nursing. Am. J. Vet. Res. 1972, 33, 1935. [Google Scholar]
- McCracken, J.A.; Custer, E.E.; Lamsa, J.C. Luteolysis: A neuroendocrine mediated event. Physiol. Rev. 1999, 79, 263–323. [Google Scholar] [CrossRef]
- Spencer, T.E.; Bazer, F.W. Conceptus signals for establishment and maintenance of pregnancy. Reprod. Biol. Endocrin. 2004, 2, 49. [Google Scholar] [CrossRef] [PubMed]
- Tomanelli, R.N.; Sertich, P.L.; Watson, E.D. Soluble oestrogen and progesterone receptors in the endometrium of the mare. J. Reprod. Fertil. Suppl. 1991, 44, 267–273. [Google Scholar] [PubMed]
- Watson, E.D.; Skolnik, S.B.; Zanecosky, H.G. Progesterone and estrogen receptor distribution in the endometrium of mares. Theriogenology 1992, 32, 197–204. [Google Scholar] [CrossRef] [PubMed]
- McDowell, K.J.; Adams, M.H.; Adam, C.Y.; Simpson, K.S. Changes in equine endometrial oestrogen receptor α and progesterone receptor mRNAs during the oestrous cycle, early pregnancy and after treatment with exogenous steroids. J. Reprod. Fertil. 1999, 117, 135–142. [Google Scholar] [CrossRef] [PubMed]
- Aupperle, H.; Ozgen, S.; Schoon, H.A.; Schoon, D.; Hoppen, H.O.; Sieme, H.; Tannapfel, A. Cyclic endometrial steroid hormone receptor expression and proliferation intensity in the mare. Equine Vet. J. 2000, 32, 228–232. [Google Scholar] [CrossRef]
- Hartt, L.S.; Carling, S.J.; Joyce, M.M.; Johnson, G.A.; Vanderwall, D.K.; Ott, T.L. Temporal and spatial associations of oestrogen receptor alpha and progesterone receptor in the endometrium of cyclic and early pregnant mares. Reproduction 2005, 130, 241–250. [Google Scholar] [CrossRef]
- Maciel, L.F.S.; Silva, E.S.M.; Oliveira-Filho, J.P.; Fritsch, S.C.; Rossi, R.S.; Lourenção, J.A.C.; Meira, C. Endometrial expression of estrogen and progesterone receptors in non-cyclic mares treated only with long-acting progesterone. Theriogenology 2018, 108, 185–191. [Google Scholar] [CrossRef]
- Woody, C.O.; First, N.L.; Pope, A.L. Effect of exogenous progesterone on cycle length. J. Anim. Sci. 1967, 26, 139–141. [Google Scholar] [CrossRef]
- Garrett, J.E.; Geisert, R.D.; Zavy, M.T.; Gries, L.K.; Wettemann, R.P.; Buchanan, D.S. Effect of exogenous progesterone on prostaglandin F2α release and the interestrous interval in the bovine. Prostaglandins 1988, 36, 85–96. [Google Scholar] [CrossRef]
- Pope, W.F.; Cardenas, H.; Wiley, T.M.; McClure, K.E. Dose-response relationships of exogenous progesterone shortly after ovulation on estrous cycle length, blastocyst development and fertility in sheep. Anim. Reprod. Sci. 1995, 38, 109–117. [Google Scholar] [CrossRef]
- Klein, C.; Troedsson, M.H. Maternal recognition of pregnancy in the horse: A mystery still to be solved. Reprod. Fertil. Dev. 2011, 23, 952–963. [Google Scholar] [CrossRef] [PubMed]
- Klein, C. Early pregnancy in the mare: Old concepts revisited. Domest. Anim. Endocrinol. 2016, 56, S212–S217. [Google Scholar] [CrossRef]
- Stout, T.A. Embryo-maternal communication during the first 4 weeks of equine pregnancy. Theriogenology 2016, 86, 349–354. [Google Scholar] [CrossRef]
- Swegen, A. Maternal recognition of pregnancy in the mare: Does it exist and why do we care? Reproduction 2021, 161, 139–155. [Google Scholar] [CrossRef]
- Merkl, M.; Ulbrich, S.E.; COtzdorff Herbahc, N.; Wanke, R.; Wolf, E.; Handler, J. Microarray analysis of equine endometrium at Days 8 and 12 of pregnancy. Biol. Reprod. 2010, 83, 874–886. [Google Scholar] [CrossRef]
- Beyer, T.; Rink, E.; Scarlet, D.; Walter, I.; Kunert, S.; Aurich, C. Early luteal phase progestin concentration influences endometrial function in pregnant mares. Theriogenology 2019, 125, 236–241. [Google Scholar] [CrossRef]
- Hoffmann, C.; Ellenberger, C.; Mattos, R.C.; Aupperle, H.; Dhein, S.; Stief, B.; Schoon, H.-A. The equine endometrosis: New insights into the pathogenesis. Anim. Reprod. Sci. 2009, 111, 261–278. [Google Scholar] [CrossRef] [PubMed]
- Ginther, O.J. Mobility of the early equine conceptus. Theriogenology 1983, 19, 603–611. [Google Scholar] [CrossRef]
- Ginther, O.J. Intrauterine movement of the early conceptus in barren and post-partum mares. Theriogenology 1984, 22, 83–95. [Google Scholar] [CrossRef]
- Leith, G.S.; Ginther, O.J. Characterisation of intrauterine mobility of the early equine conceptus. Theriogenology 1984, 22, 401–408. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Mare ID | Hours Post Ovulation When Embryo Reduction Was Undertaken | No. of Singleton Reductions | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| 244 | 248 | 252 | 256 | 260 | 264 | 268 | 272 | 276 | 280 | 284 | 288 | 292 | 296 | 300 | 304 | 308 | 312 | 316 | 320 | 324 | 328 | 332 | 336 | 340 | 344 | 348 | 352 | 356 | ||
| Day 10 | Day 11 | Day 12 | Day 13 | Day 14 | ||||||||||||||||||||||||||
| ZA | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 | 0/1 | 0/2 | 0/1 | 0/1 | 0/2 | 1/2 | 1/1 | 1/2 | 2/2 | 1/1 | 1/1 | 3/3 | 24 | ||||||||||||
| AA | 0/1 | 0/1 | 0/2 | 0/1 | 0/2 | 0/1 | 1/2 | 2/3 | 2/5 | 1/2 | 2/2 | 1/1 | 23 | |||||||||||||||||
| GY | 0/1 | 0/1 | 0/1 | 0/2 | 0/1 | 1/3 | 0/2 | 1/1 | 2/2 | 0/2 | 2/4 | 1/1 | 1/1 | 22 | ||||||||||||||||
| CO | 0/1 | 0/1 | 0/1 | 0/1 | 2/2 | 1/1 | 1/1 | 1/1 | 1/1 | 10 | ||||||||||||||||||||
| DE | 0/1 | 0/1 | 1/1 | 1/1 | 1/1 | 5 | ||||||||||||||||||||||||
| IR | 0/1 | 0/1 | 1/1 | 1/1 | 1/1 | 1/1 | 6 | |||||||||||||||||||||||
| PS | 0/1 | 0/1 | 1/1 | 1/1 | 1/1 | 1/1 | 1/1 | 7 | ||||||||||||||||||||||
| PY | 0/1 | 0/1 | 3/6 | 2/2 | 4/5 | 3/3 | 2/2 | 1/1 | 1/1 | 1/1 | 23 | |||||||||||||||||||
| RS | 0/1 | 0/1 | 1/2 | 2/2 | 2/3 | 0/1 | 1/1 | 1/1 | 12 | |||||||||||||||||||||
| CSO | 0/1 | 0/1 | 1/2 | 0/1 | 1/2 | 2/3 | 1/1 | 1/1 | 2/2 | 2/2 | 1/1 | 1/1 | 18 | |||||||||||||||||
| Day of Embryo Reduction | Median (25–75% Range; n) Embryo Diameter (mm) of Vesicles at the Time of Reduction in Mares That Became: | |
|---|---|---|
| Luteolytic | Luteostatic | |
| Day 10 | 4.5 (4.2–5.4; n = 5) | 6.6 (6.2–7.0; n = 2) |
| Day 11 | 7.1 (5.5–7.7; n = 21) | 6.8 (5.1–8.3; n = 29) |
| Day 12 | 8.3 (7.4–10.3; n = 12) | 9.8 (7.7–11.2; n = 18) |
| Day 13 | 12.7 (7.7–14.7; n = 4) | 10.8 (7.3–13.2; n = 6) |
| Day 14 | 12.9 (8.6–17.1; n = 2) | 15.1 (13.9–20.7; n = 7) |
| Day and Hour of Embryo Reduction | Median (25–75% Range; n) Embryo Diameter (mm) of Vesicles at the Time of Reduction in Mares That Became: | ||
|---|---|---|---|
| Luteolytic | Luteostatic | ||
| Day 11 | 0–4 h | 5.7 (5.0–6.9; n = 9) | 6.0 (4.4–7.3; n = 6) |
| 8–12 h | 7.2 (6.1–7.7; n = 8) | 5.8 (5.0–7.9; n = 8) | |
| 16–20 h | 7.7 (6.2–8.1; n = 4) | 7.7 (5.9–9.1; n = 15) | |
| Overall | 7.1 (5.5–7.7; n = 21) | 6.8 (5.1–8.3; n = 29) | |
| Day 12 | 0–4 h | 8.6 (7.3–10.3; n = 3) | 9.5 (5.5–11.1; n = 8) |
| 8–12 h | 7.7 (6.7–8.5; n = 7) | 9.9 (6.9–11.4; n = 5) | |
| 16–20 h | 10.9 (10.2–11.6; n = 2) | 9.7 (7.6–11.9; n = 5) | |
| Overall | 8.3 (7.4–10.3; n = 12) | 9.8 (7.7–11.2; n = 18) | |
| Mare ID | Day and Hours Post-Ovulation of Embryo Reduction | Median (25–75% Range; n) Embryonic Vesicle Diameter (mm) Prior to Reduction and Timing of Reduction (h Post-Ovulation) in Mares That Became: | |
|---|---|---|---|
| Luteolytic | Luteostatic | ||
| AA | Day 12 | 7.2 (5.2–8.4; n = 4) | 13.5 (8.7–11.9; n = 6) |
| Hours | 302 (294–304; n = 4) | 300 (295–305; n = 6) | |
| GY | Day 12 | 8.3 (7.5–9.8; n = 4) | 9.6 (8.7–10.0; n = 6) |
| Hours | 304 (295–307; n = 4) | 304 (299–309; n = 6) | |
| PY | Day 11 | 4.8 (4.3–5.2; n = 5) | 6.2 (4.7–7.1; n = 12) |
| Hours | 268 (268–272; n = 4) a | 276 (272–284; n = 6) b | |
| Mare ID | Cycle Length (Mean Days ± SEM) | ||
|---|---|---|---|
| Pregnant, Embryo Reduction with No Luteostasis | Not Pregnant | All Cycles | |
| ZA | 20.7 ± 0.7 (n = 12) | 20.8 ± 1.1 (n = 5) | 20.8 ± 0.7 (n = 17) a |
| CSO | 21.3 ± 1.3 (n = 4) | 20.8 ± 1.1 (n = 6) | 21.0 ± 0.8 (n = 10) a |
| PY | 21.1 ± 1.3 (n = 4) | 21.7 ± 1.8 (n = 2) | 21.4 ± 1.1 (n = 6) |
| GY | 21.4 ± 1.0 (n = 7) | 21.9 ± 1.0 (n = 7) | 21.6 ± 0.7 (n = 14) a |
| RS | 22.0 ± 1.5 (n = 3) | 22.3 ± 1.3 (n = 4) | 22.1 ± 1.0 (n = 7) |
| CO | 24.0 ± 1.5 (n = 3) | 21.3 ± 1.5 (n = 3) | 22.7 ± 1.1 (n = 6) |
| AA | 24.9 ± 0.9 (n = 8) | 24.4 ± 0.8 (n = 10) | 24.6 ± 0.6 (n = 18) b |
| All mares | 22.2 ± 0.5 (n = 41) | 21.9 ± 0.5 (n = 37) | 22.1 ± 0.3 (n = 78) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Newcombe, J.R.; Cuervo-Arango, J.; Wilsher, S. The Timing of the Maternal Recognition of Pregnancy Is Specific to Individual Mares. Animals 2023, 13, 1718. https://doi.org/10.3390/ani13101718
Newcombe JR, Cuervo-Arango J, Wilsher S. The Timing of the Maternal Recognition of Pregnancy Is Specific to Individual Mares. Animals. 2023; 13(10):1718. https://doi.org/10.3390/ani13101718
Chicago/Turabian StyleNewcombe, John R., Juan Cuervo-Arango, and Sandra Wilsher. 2023. "The Timing of the Maternal Recognition of Pregnancy Is Specific to Individual Mares" Animals 13, no. 10: 1718. https://doi.org/10.3390/ani13101718
APA StyleNewcombe, J. R., Cuervo-Arango, J., & Wilsher, S. (2023). The Timing of the Maternal Recognition of Pregnancy Is Specific to Individual Mares. Animals, 13(10), 1718. https://doi.org/10.3390/ani13101718