
One Health Approach: Invasive California Kingsnake (Lampropeltis californiae) as an Important Source of Antimicrobial Drug-Resistant Salmonella Clones on Gran Canaria Island
 , , , ,
, , , ,  , , ,
, , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
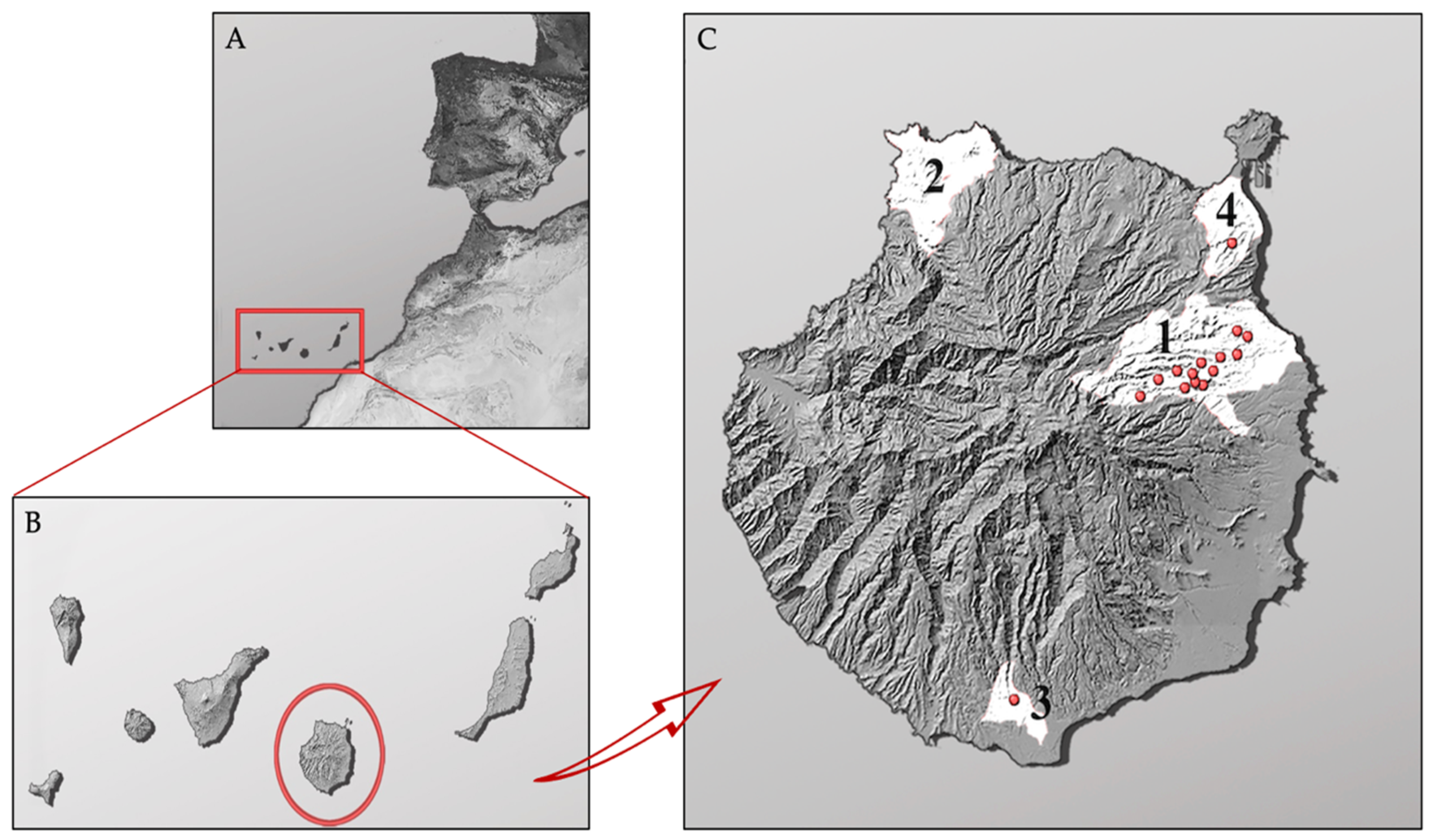
2.1. Sample Collection
2.2. Salmonella Isolation
2.3. Molecular Typing of Salmonella Isolates
2.4. Antimicrobial Susceptibility Testing
2.5. Statistical Analysis
3. Results
3.1. Salmonella spp. Identification and Serotyping
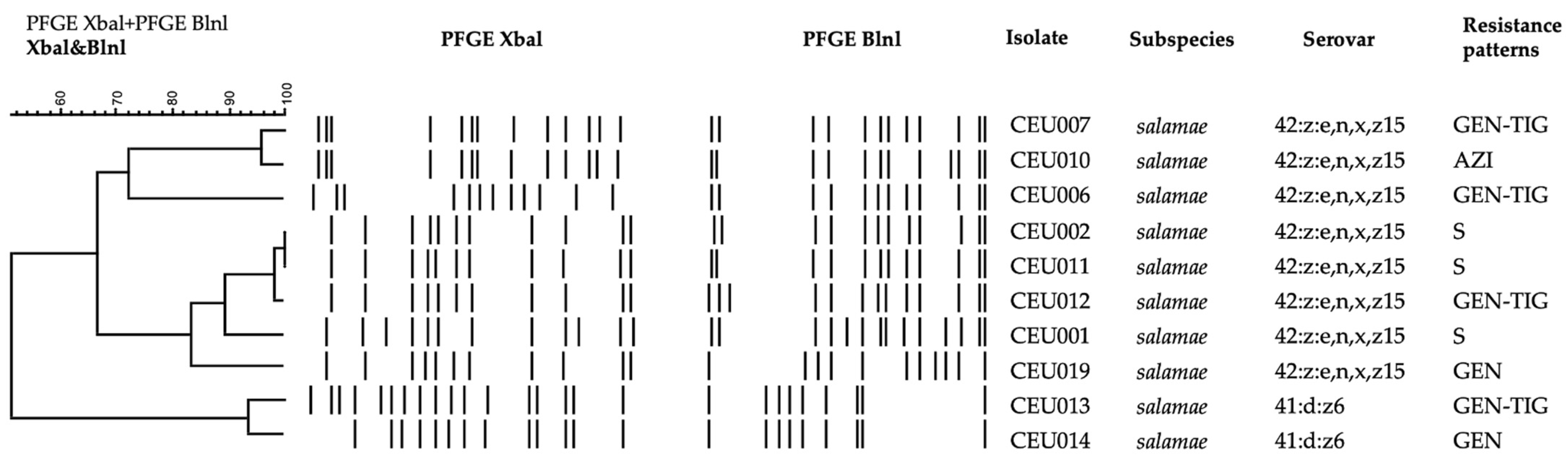
3.2. Salmonella Molecular Typing
3.3. Antimicrobial Susceptibility
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Santana-Hernández, K.M.; Javorská, K.; Rodríguez-Ponce, E.; Fecková, B.; Šlapeta, J.; Modrý, D. Sarcocystis Sp. Infection (Apicomplexa: Sarcocystidae) in Invasive California Kingsnake Lampropeltis californiae (Serpentes: Colubridae) in Gran Canaria. Parasitology 2022, 149, 1419–1424. [Google Scholar] [CrossRef] [PubMed]
- Piquet, J.C.; Warren, D.L.; Saavedra Bolaños, J.F.; Sánchez Rivero, J.M.; Gallo-Barneto, R.; Cabrera-Pérez, M.Á.; Fisher, R.N.; Fisher, S.R.; Rochester, C.J.; Hinds, B.; et al. Could Climate Change Benefit Invasive Snakes? Modelling the Potential Distribution of the California Kingsnake in the Canary Islands. J. Environ. Manag. 2021, 294, 112917. [Google Scholar] [CrossRef] [PubMed]
- Piquet, J.C.; Maestresalas, B.; López-Darias, M. Coupling Phenotypic Changes to Extinction and Survival in an Endemic Prey Community Threatened by an Invasive Snake. Sci. Rep. 2022, 12, 18249. [Google Scholar] [CrossRef] [PubMed]
- FEDIAF (The European Pet Food Industry). FEDIAF Annual Report 2022; FEDIAF: Brussels, Belgium, 2022. [Google Scholar]
- Fisher, S.R.; Fisher, R.N.; Alcaraz, S.E.; Gallo-Barneto, R.; Patino-Martinez, C.; López-Jurado, L.F.; Rochester, C.J. Life-History Comparisons between the Native Range and an Invasive Island Population of a Colubrid Snake. Available online: https://www.researchgate.net/publication/333262467_Life-history_comparisons_between_the_native_range_and_an_invasive_island_population_of_a_colubrid_snake (accessed on 28 November 2022).
- Marin, C.; Ingresa-Capaccioni, S.; González-Bodi, S.; Marco-Jiménez, F.; Vega, S. Free-Living Turtles Are a Reservoir for Salmonella but Not for Campylobacter. PLoS ONE 2013, 8, e72350. [Google Scholar] [CrossRef] [PubMed]
- Kikillus, K.H.; Gartrell, B.D.; Motion, E. Prevalence of Salmonella Spp., and Serovars Isolated from Captive Exotic Reptiles in New Zealand. N. Z. Vet. J. 2011, 59, 174–178. [Google Scholar] [CrossRef] [PubMed]
- Hydeskov, H.B.; Guardabassi, L.; Aalbæk, B.; Olsen, K.E.P.; Nielsen, S.S.; Bertelsen, M.F. Salmonella Prevalence among Reptiles in a Zoo Education Setting. Zoonoses Public Health 2013, 60, 291–295. [Google Scholar] [CrossRef] [PubMed]
- Pedersen, K.; Lassen-Nielsen, A.M.; Nordentoft, S.; Hammer, A.S. Serovars of Salmonella from Captive Reptiles. Zoonoses Public Health 2009, 56, 238–242. [Google Scholar] [CrossRef]
- Bjelland, A.M.; Sandvik, L.M.; Skarstein, M.M.; Svendal, L.; Debenham, J.J. Prevalence of Salmonella Serovars Isolated from Reptiles in Norwegian Zoos. Acta Vet. Scand. 2020, 62, 1–9. [Google Scholar] [CrossRef]
- Marin, C.; Martín-Maldonado, B.; Cerdà-Cuéllar, M.; Sevilla-Navarro, S.; Lorenzo-Rebenaque, L.; Montoro-Dasi, L.; Manzanares, A.; Ayats, T.; Mencía-Gutiérrez, A.; Jordá, J.; et al. Antimicrobial Resistant Salmonella in Chelonians: Assessing Its Potential Risk in Zoological Institutions in Spain. Vet. Sci. 2022, 9, 264. [Google Scholar] [CrossRef]
- Kanagarajah, S.; Waldram, A.; Dolan, G.; Jenkins, C.; Ashton, P.M.; Carrion Martin, A.I.; Davies, R.; Frost, A.; Dallman, T.J.; de Pinna, E.M.; et al. Whole Genome Sequencing Reveals an Outbreak of Salmonella Enteritidis Associated with Reptile Feeder Mice in the United Kingdom, 2012–2015. Food Microbiol. 2018, 71, 32–38. [Google Scholar] [CrossRef]
- Marin, C.; Martelli, F.; Rabie, A.; Davies, R. Commercial Frozen Mice Used by Owners to Feed Reptiles Are Highly Externally Contaminated with Salmonella Enteritidis PT8. Vector Borne Zoonotic Dis. 2018, 18, 453–457. [Google Scholar] [CrossRef] [PubMed]
- Marin, C.; Vega, S.; Marco-Jiménez, F. Tiny Turtles Purchased at Pet Stores Are a Potential High Risk for Salmonella Human Infection in the Valencian Region, Eastern Spain. Vector Borne Zoonotic Dis. 2016, 16, 455–460. [Google Scholar] [CrossRef] [PubMed]
- Zając, M.; Wasyl, D.; Różycki, M.; Bilska-Zając, E.; Fafiński, Z.; Iwaniak, W.; Krajewska, M.; Hoszowski, A.; Konieczna, O.; Fafińska, P.; et al. Free-Living Snakes as a Source and Possible Vector of Salmonella Spp. and Parasites. Eur. J. Wildl. Res. 2016, 62, 161–166. [Google Scholar] [CrossRef]
- Alikhan, N.F.; Zhou, Z.; Sergeant, M.J.; Achtman, M. A Genomic Overview of the Population Structure of Salmonella. PLoS Genet. 2018, 14, e1007261. [Google Scholar] [CrossRef] [PubMed]
- Gwyn, L.B. On Infection with a Para-Colin Bacillus in a Case with All the Clinical Features of Typhoid Fever. Johns Hopkins Hosp. Bull. 1898, 84, 54–56. [Google Scholar]
- Grimont, P.A.D.; Weill, F.-X. Antigenic Formulae of the Salmonella Serovars, 9th ed.; WHO Collaborating Centre for Reference and Research on Salmonella: Paris, France, 2007. [Google Scholar]
- Piquet, J.C.; López-Darias, M. Spatial Ecology to Strengthen Invasive Snake Management on Islands. Sci. Rep. 2022. in print. [Google Scholar] [CrossRef]
- Piquet, J.C.; López-Darias, M. Invasive Snake Causes Massive Reduction of All Endemic Herpetofauna on Gran Canaria. Proc. R. Soc. B Biol. Sci. 2021, 288, 20211939. [Google Scholar] [CrossRef]
- McWhorter, A.; Owens, J.; Valcanis, M.; Olds, L.; Myers, C.; Smith, I.; Trott, D.; McLelland, D. In Vitro Invasiveness and Antimicrobial Resistance of Salmonella enterica Subspecies Isolated from Wild and Captive Reptiles. Zoonoses Public Health 2021, 68, 402–412. [Google Scholar] [CrossRef] [PubMed]
- Marco-Fuertes, A.; Marin, C.; Lorenzo-Rebenaque, L.; Vega, S.; Montoro-Dasi, L. Antimicrobial Resistance in Companion Animals: A New Challenge for the One Health Approach in the European Union. Vet. Sci. 2022, 9, 208. [Google Scholar] [CrossRef] [PubMed]
- Vega, S.; Marín, C. ¿Qué Es One Health? In One Health: Cambio Climático, Contaminación Ambiental y el Impacto Sobre la Salud Humana y Animal; Ábrego Bonaforte, J., Díaz Sáez, G.J., Eds.; Amazing Books: Zaragoza, Spain, 2019; pp. 31–60. ISBN 978-84-17403-51-5. [Google Scholar]
- Edrington, T.S.; Farrow, R.L.; Hume, M.E.; Anderson, P.N.; Hagevoort, G.R.; Caldwell, D.J.; Callaway, T.R.; Anderson, R.C.; Nisbet, D.J. Evaluation of the Potential Antimicrobial Resistance Transfer from a Multi-Drug Resistant Escherichia coli to Salmonella in Dairy Calves. Curr. Microbiol. 2013, 66, 132–137. [Google Scholar] [CrossRef]
- Kwon, J.; Kim, S.G.; Kim, H.J.; Giri, S.S.; Kim, S.W.; Lee, S.B.; Park, S.C. Bacteriophage as an Alternative to Prevent Reptile-Associated Salmonella Transmission. Zoonoses Public Health 2021, 68, 131–143. [Google Scholar] [CrossRef] [PubMed]
- Kiebler, C.A.; Bottichio, L.; Simmons, L.; Basler, C.; Klos, R.; Gurfield, N.; Roberts, E.; Kimura, A.; Lewis, L.S.; Bird, K.; et al. Outbreak of Human Infections with Uncommon Salmonella Serotypes Linked to Pet Bearded Dragons, 2012–2014. Zoonoses Public Health 2020, 67, 425–434. [Google Scholar] [CrossRef] [PubMed]
- EUR-Lex—32013D0652—ES—EUR-Lex. Available online: https://eur-lex.europa.eu/eli/dec_impl/2013/652/oj?locale=es (accessed on 29 November 2022).
- EUCAST: New S, I and R Definitions. Available online: https://www.eucast.org/newsiandr (accessed on 29 November 2022).
- Marin, C.; Lorenzo-Rebenaque, L.; Laso, O.; Villora-Gonzalez, J.; Vega, S. Pet Reptiles: A Potential Source of Transmission of Multidrug-Resistant Salmonella. Front. Vet. Sci. 2021, 7, 1157. [Google Scholar] [CrossRef] [PubMed]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-Resistant, Extensively Drug-Resistant and Pandrug-Resistant Bacteria: An International Expert Proposal for Interim Standard Definitions for Acquired Resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed]
- Merkevičienė, L.; Butrimaitė-Ambrozevičienė, Č.; Paškevičius, G.; Pikūnienė, A.; Virgailis, M.; Dailidavičienė, J.; Daukšienė, A.; Šiugždinienė, R.; Ruzauskas, M. Serological Variety and Antimicrobial Resistance in Salmonella Isolated from Reptiles. Biology 2022, 11, 836. [Google Scholar] [CrossRef] [PubMed]
- Dec, M.; Zając, M.; Puchalski, A.; Szczepaniak, K.; Urban-Chmiel, R. Pet Reptiles in Poland as a Potential Source of Transmission of Salmonella. Pathogens 2022, 11, 1125. [Google Scholar] [CrossRef] [PubMed]
- Whitten, T.; Bender, J.B.; Smith, K.; Leano, F.; Scheftel, J. Reptile-Associated Salmonellosis in Minnesota, 1996-2011. Zoonoses Public Health 2015, 62, 199–208. [Google Scholar] [CrossRef]
- View of Prevalence, Risk Factors and Antimicrobial Resistance of Salmonella Infection in Turtles in Khon Kaen Province. Available online: https://he01.tci-thaijo.org/index.php/tjvm/article/view/247794/167908 (accessed on 18 January 2023).
- Colon, V.A.; Lugsomya, K.; Lam, H.K.; Wahl, L.C.; Parkes, R.S.V.; Cormack, C.A.; Horlbog, J.A.; Stevens, M.; Stephan, R.; Magouras, I. Serotype Diversity and Antimicrobial Resistance Profile of Salmonella enterica Isolates from Freshwater Turtles Sold for Human Consumption in Wet Markets in Hong Kong. Front. Vet. Sci. 2022, 9, 912693. [Google Scholar] [CrossRef] [PubMed]
- Casalino, G.; Bellati, A.; Pugliese, N.; Camarda, A.; Faleo, S.; Lombardi, R.; Occhiochiuso, G.; D’onghia, F.; Circella, E. Salmonella Infection in Turtles: A Risk for Staff Involved in Wildlife Management? Animals 2021, 11, 1529. [Google Scholar] [CrossRef]
- Kuroki, T.; Ishihara, T.; Furukawa, I.; Okatani, A.T.; Kato, Y. Prevalence of Salmonella in Wild Snakes in Japan. Jpn. J. Infect. Dis. 2013, 66, 295–298. [Google Scholar] [CrossRef]
- Piasecki, T.; Chrząstek, K.; Wieliczko, A. Salmonella Serovar Spectrum Associated with Reptiles in Poland. Acta Vet. Brno 2014, 83, 287–294. [Google Scholar] [CrossRef]
- Sadiq, M.S.; Othman, R.M. Phylogenetic Tree Constructed of Salmonella enterica Subspecies Enterica Isolated from Animals and Humans in Basrah and Baghdad Governorates, Iraq. Iraqi J. Vet. Sci. 2022, 36, 895–903. [Google Scholar] [CrossRef]
- Gambi, L.; Ravaioli, V.; Rossini, R.; Tranquillo, V.; Boscarino, A.; Mattei, S.; D’incau, M.; Tosi, G.; Fiorentini, L.; di Donato, A. Prevalence of Different Salmonella enterica Subspecies and Serotypes in Wild Carnivores in Emilia-Romagna Region, Italy. Animals 2022, 12, 3368. [Google Scholar] [CrossRef]
- Gil-Molino, M.; Gonçalves, P.; Risco, D.; Martín-Cano, F.E.; García, A.; Rey, J.; Fernández-Llario, P.; Quesada, A. Dissemination of Antimicrobial-Resistant Isolates of Salmonella Spp. in Wild Boars and Its Relationship with Management Practices. Transbound. Emerg. Dis. 2022, 69, e1488–e1502. [Google Scholar] [CrossRef] [PubMed]
- Shalaby, A.; Ismail, M.M.; El-Sharkawy, H. Isolation, Identification, and Genetic Characterization of Antibiotic Resistance of Salmonella Species Isolated from Chicken Farms. J. Trop. Med. 2022, 2022, 6065831. [Google Scholar] [CrossRef] [PubMed]
- López-Quintana, B.; Rivas-González, P.; Toro-Rueda, C.; Enríquez-Crego, A. Infección Por Salmonella enterica Subespecie Salamae En Un Paciente Ecuatoguineano Consumidor de Carne de Tortuga. Enferm. Infecc. Microbiol. Clínica 2015, 33, 430–431. [Google Scholar] [CrossRef] [PubMed]
- Yan, M.; Zhou, Y.; Cao, Y.; Li, Z.; Lu, X.; Pang, B.; Wang, S.; Kan, B. Salmonella enterica Subsp. II Serovar 4,5,12:A:- May Cause Gastroenteritis Infections in Humans. Gut Microbes 2022, 14, 2089007. [Google Scholar] [CrossRef]
- Hounmanou, Y.M.G.; Baniga, Z.; García, V.; Dalsgaard, A. Salmonella salamae and S. waycross Isolated from Nile Perch in Lake Victoria Show Limited Human Pathogenic Potential. Sci. Rep. 2022, 12, 4229. [Google Scholar] [CrossRef] [PubMed]
- Pulford, C.V.; Wenner, N.; Redway, M.L.; Rodwell, E.V.; Webster, H.J.; Escudero, R.; Kröger, C.; Canals, R.; Rowe, W.; Lopez, J.; et al. The Diversity, Evolution and Ecology of Salmonella in Venomous Snakes. PLoS Negl. Trop. Dis. 2019, 13, e0007169. [Google Scholar] [CrossRef] [PubMed]
- Pławińska-Czarnak, J.; Wódz, K.; Piechowicz, L.; Tokarska-Pietrzak, E.; Bełkot, Z.; Bogdan, J.; Wiśniewski, J.; Kwieciński, P.; Kwieciński, A.; Anusz, K. Wild Duck (Anas Platyrhynchos) as a Source of Antibiotic-Resistant Salmonella enterica Subsp. Diarizonae O58—The First Report in Poland. Antibiotics 2022, 11, 530. [Google Scholar] [CrossRef] [PubMed]
- Andruzzi, M.N.; Krath, M.L.; Lawhon, S.D.; Boudreau, B. Salmonella enterica Subspecies Houtenae as an Opportunistic Pathogen in a Case of Meningoencephalomyelitis and Bacteriuria in a Dog. BMC Vet. Res. 2020, 16, 437. [Google Scholar] [CrossRef] [PubMed]
- Uelze, L.; Borowiak, M.; Flieger, A.; Simon, S.; Tausch, S.H.; Malorny, B. Complete Genome Sequence of Salmonella enterica Subsp. Diarizonae Serovar 61:K:1,5,(7) Strain 14-SA00836-0, Isolated from Human Urine. Microbiol. Resour. Announc. 2020, 9, e00683-20. [Google Scholar] [CrossRef] [PubMed]
- Horvath, L.; Kraft, M.; Fostiropoulos, K.; Falkowski, A.; Tarr, P.E. Salmonella enterica Subspecies Diarizonae Maxillary Sinusitis in a Snake Handler: First Report. Open Forum Infect. Dis. 2016, 3, ofw066. [Google Scholar] [CrossRef] [PubMed]
- Karle, E.; Schuh, S.; Ziadeh, A.; Patil, S.M. A Rare Cause of Community-Acquired Bacterial Pneumonia in a Patient with Poorly Controlled Diabetes. Chest 2022, 162, A514. [Google Scholar] [CrossRef]
- Zajaç, M.; Skarżyńska, M.; Lalak, A.; Kwit, R.; Śmiałowska-Węglińska, A.; Pasim, P.; Szulowski, K.; Wasyl, D. Salmonella in Captive Reptiles and Their Environment—Can We Tame the Dragon? Microorganisms 2021, 9, 1012. [Google Scholar] [CrossRef] [PubMed]
- Xu, H.; Cao, X.; Zhang, W.; Li, Q. Co-Existence of Two Ciprofloxacin-Resistant Salmonella enterica Serovar Kentucky Strains in the Urine of a Bladder Cancer Patient. J. Glob. Antimicrob. Resist. 2022, 30, 354–356. [Google Scholar] [CrossRef] [PubMed]
- Neelambike, S.M.; Chitharagi, V.; Rao, M.R.; Badveti, S.; Rao, R. Ceftriaxone Resistant Salmonella enterica Serovar Kentucky as a Cause of Bacteremia and Gastroenteritis: A Case Report. J. Lab. Physicians 2023, 15, 175–178. [Google Scholar] [CrossRef] [PubMed]
- Zajac, M.; Wasyl, D.; Hoszowski, A.; le Hello, S.; Szulowski, K. Genetic Lineages of Salmonella enterica Serovar Kentucky Spreading in Pet Reptiles. Vet. Microbiol. 2013, 166, 686–689. [Google Scholar] [CrossRef]
- Slowey, R.; Kim, S.W.; Prendergast, D.; Madigan, G.; van Kessel, J.A.S.; Haley, B.J. Genomic Diversity and Resistome Profiles of Salmonella enterica Subsp. Enterica Serovar Kentucky Isolated from Food and Animal Sources in Ireland. Zoonoses Public Health 2022, 69, 1–12. [Google Scholar] [CrossRef]
- Salaheen, S.; Kim, S.W.; Haley, B.J.; van Kessel, J.A.S. Differences between the Global Transcriptomes of Salmonella enterica Serovars Dublin and Cerro Infecting Bovine Epithelial Cells. BMC Genom. 2022, 23, 498. [Google Scholar] [CrossRef] [PubMed]
- Cohn, A.R.; Orsi, R.H.; Carroll, L.M.; Liao, J.; Wiedmann, M.; Cheng, R.A. Salmonella enterica Serovar Cerro Displays a Phylogenetic Structure and Genomic Features Consistent with Virulence Attenuation and Adaptation to Cattle. Front. Microbiol. 2022, 13, 4725. [Google Scholar] [CrossRef] [PubMed]
- Weerakhun, S.; Kongsanan, P.; Kongkham, W. Distribution of Salmonella Spp. and Serovars Isolated from Captive Reptiles in Four Zoos of the Zoological Park Organization, Thailand, between 2017 and 2018. Thai J. Vet. Med. 2022, 52, 349–358. [Google Scholar]
- Cambre, R.C.; Earl Green, D.; Smith, E.E.; Montali, R.J. Salmoneilosis and Arizonosis in the Reptile Collection at the National Zoological Park. J. Am. Vet. Med. Assoc. 1980, 177, 803. [Google Scholar]
- Scheelings, T.F.; Lightfoot, D.; Holz, P. Prevalence of Salmonella in Australian Reptiles. J. Wildl. Dis. 2011, 47, 1–11. [Google Scholar] [CrossRef]
- Nakadai, A.; Kuroki, T.; Kato, Y.; Suzuki, R.; Yamai, S.; Yaginuma, C.; Shiotani, R.; Yamanouchi, A.; Hayashidani, H. Prevalence of Salmonella Spp. in Pet Reptiles in Japan. J. Vet. Med. Sci. 2005, 67, 97–101. [Google Scholar] [CrossRef] [PubMed]
- Moran, A.B.; Mcwhorter, A.C. A Hitherto Undescribed Salmonella Serotype: S. hennepin. Int. J. Syst. Evol. Microbiol. 1966, 16, 143–144. [Google Scholar] [CrossRef]
- Brown, E.W.; Bell, R.; Zhang, G.; Timme, R.; Zheng, J.; Hammack, T.S.; Allard, M.W. Salmonella Genomics in Public Health and Food Safety. EcoSal Plus 2021, 9, eESP-0008. [Google Scholar] [CrossRef]
- Rush, E.M.; Amadi, V.A.; Johnson, R.; Lonce, N.; Hariharan, H. Salmonella Serovars Associated with Grenadian Tree Boa (Corallus grenadensis) and Their Antimicrobial Susceptibility. Vet. Med. Sci. 2020, 6, 565–569. [Google Scholar] [CrossRef]
- Ramos, C.P.; Santana, J.A.; Morcatti Coura, F.; Xavier, R.G.C.; Leal, C.A.G.; Oliveira Junior, C.A.; Heinemann, M.B.; Lage, A.P.; Lobato, F.C.F.; Silva, R.O.S. Identification and Characterization of Escherichia coli, Salmonella Spp., Clostridium perfringens, and C. difficile Isolates from Reptiles in Brazil. Biomed. Res. Int. 2019, 2019, 9530732. [Google Scholar] [CrossRef]
- Thomas, M.; Fenske, G.J.; Antony, L.; Ghimire, S.; Welsh, R.; Ramachandran, A.; Scaria, J. Whole Genome Sequencing-Based Detection of Antimicrobial Resistance and Virulence in Non-Typhoidal Salmonella enterica Isolated from Wildlife. Gut Pathog. 2017, 9, 66. [Google Scholar] [CrossRef] [PubMed]
- Bertelloni, F.; Chemaly, M.; Cerri, D.; le Gall, F.; Ebani, V.V. Salmonella Infection in Healthy Pet Reptiles: Bacteriological Isolation and Study of Some Pathogenic Characters. Acta Microbiol. Immunol. Hung. 2016, 63, 203–216. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira Iovine, R.; Dejuste, C.; Miranda, F.; Filoni, C.; Bueno, M.G.; de Carvalho, V.M. Isolation of Escherichia coli and Salmonella Spp. from Free-Ranging Wild Animals. Braz. J. Microbiol. 2015, 46, 1257–1263. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.Y.; Chen, W.C.; Chin, S.C.; Lai, Y.H.; Tung, K.C.; Chiou, C.S.; Hsu, Y.M.; Chang, C.C. Prevalence and Antimicrobial Susceptibility of Salmonellae Isolates from Reptiles in Taiwan. J. Vet. Diagn. Investig. 2010, 22, 44–50. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| n | % Salmonella spp. | p-Value | ||
|---|---|---|---|---|
| Location | 1 | 57 | 86.6 | |
| (nuclei) | 2 | 4 | 0 | |
| 3 | 4 | 6.7 | ||
| 4 | 8 | 6.7 | ||
| Sex | Female | 37 | 73.3 a | p = 0.042 |
| Male | 36 | 26.7 b | ||
| Body Length | ≤900 cm | 33 | 40.0 | p = 0.647 |
| >900 cm | 40 | 60.0 | ||
| Weight | ≤250 g | 39 | 46.7 | p = 0.732 |
| 251–500 g | 28 | 40.0 | ||
| >500 g | 6 | 13.3 |
| Subspecies | Serovar | n | Nuclei | Prevalence (%) |
|---|---|---|---|---|
| salamae | 42:z:e,n.x.z15 | 8 | 1 | 53.3 a |
| 41:d:z6 | 2 | 1 | 13.2 b | |
| enterica | Cerro | 1 | 1 | 6.7 b |
| Kentucky | 1 | 3 | 6.7 b | |
| Midway | 1 | 1 | 6.7 b | |
| diarizonae | 60:-:- | 1 | 4 | 6.7 b |
| houtenae | 43:z4,z23:- | 1 | 1 | 6.7 b |
| Species | Serovar | n | CIP | AMP | NAL | GEN | CHL | TRS | TRI | COL | CTA | AZI | CTZ | TIG |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| S. enterica subsp. salamae | 42:z:e,n.x.z15 | 8 | 0 | 0 | 0 | 4 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 3 |
| 41:d:z6 | 2 | 0 | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | |
| S. enterica subsp. enterica | Cerro | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 1 |
| Kentucky | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Midway | 1 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | |
| S. enterica subsp. diarizonae | 60:-:- | 1 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 |
| S. enterica subsp. houtenae | 43:z4,z23:- | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Santana-Hernández, K.M.; Rodríguez-Ponce, E.; Medina, I.R.; Acosta-Hernández, B.; Priestnall, S.L.; Vega, S.; Marin, C.; Cerdà-Cuéllar, M.; Marco-Fuertes, A.; Ayats, T.; et al. One Health Approach: Invasive California Kingsnake (Lampropeltis californiae) as an Important Source of Antimicrobial Drug-Resistant Salmonella Clones on Gran Canaria Island. Animals 2023, 13, 1790. https://doi.org/10.3390/ani13111790
Santana-Hernández KM, Rodríguez-Ponce E, Medina IR, Acosta-Hernández B, Priestnall SL, Vega S, Marin C, Cerdà-Cuéllar M, Marco-Fuertes A, Ayats T, et al. One Health Approach: Invasive California Kingsnake (Lampropeltis californiae) as an Important Source of Antimicrobial Drug-Resistant Salmonella Clones on Gran Canaria Island. Animals. 2023; 13(11):1790. https://doi.org/10.3390/ani13111790
Chicago/Turabian StyleSantana-Hernández, Kevin M., Eligia Rodríguez-Ponce, Inmaculada Rosario Medina, Begoña Acosta-Hernández, Simon L. Priestnall, Santiago Vega, Clara Marin, Marta Cerdà-Cuéllar, Ana Marco-Fuertes, Teresa Ayats, and et al. 2023. "One Health Approach: Invasive California Kingsnake (Lampropeltis californiae) as an Important Source of Antimicrobial Drug-Resistant Salmonella Clones on Gran Canaria Island" Animals 13, no. 11: 1790. https://doi.org/10.3390/ani13111790
APA StyleSantana-Hernández, K. M., Rodríguez-Ponce, E., Medina, I. R., Acosta-Hernández, B., Priestnall, S. L., Vega, S., Marin, C., Cerdà-Cuéllar, M., Marco-Fuertes, A., Ayats, T., García Beltrán, T., & Lupiola-Gómez, P. A. (2023). One Health Approach: Invasive California Kingsnake (Lampropeltis californiae) as an Important Source of Antimicrobial Drug-Resistant Salmonella Clones on Gran Canaria Island. Animals, 13(11), 1790. https://doi.org/10.3390/ani13111790