


Developmental Changes of Duckling Liver and Isolation of Primary Hepatocytes

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethics Approval
2.2. Animals and Liver Sampling
2.3. Liver Sections and Oil Red O Staining
2.4. Isolation and Culture Procedures
2.5. Cell Proliferation Assays
2.6. Periodic Acid Schiff (PAS) Staining
2.7. Reverse Transcription and PCR Verification
2.8. Immunohistochemistry and Immunofluorescence
2.9. Statistical Analysis
3. Results
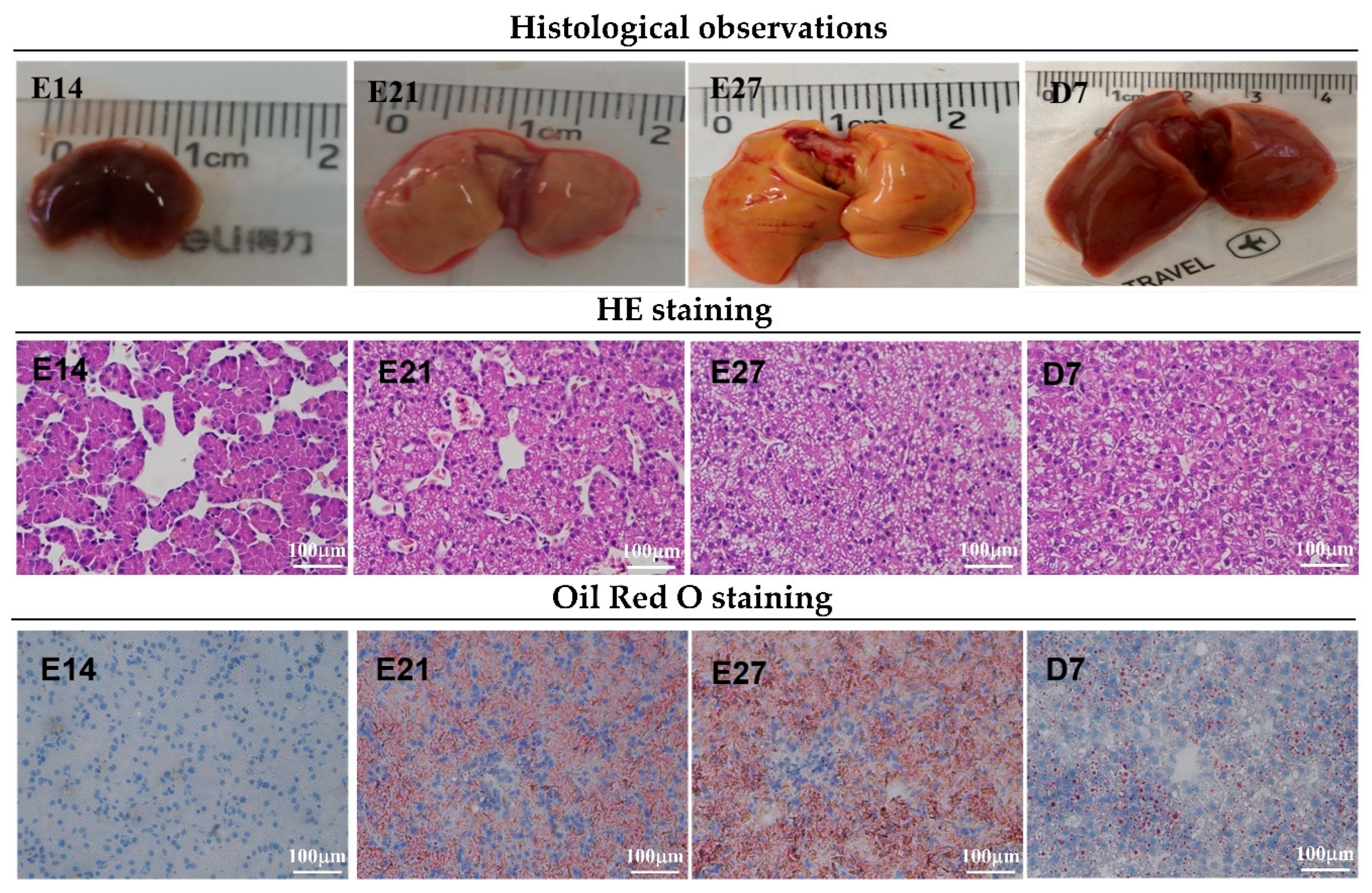
3.1. Morphological Observations of Duck Livers
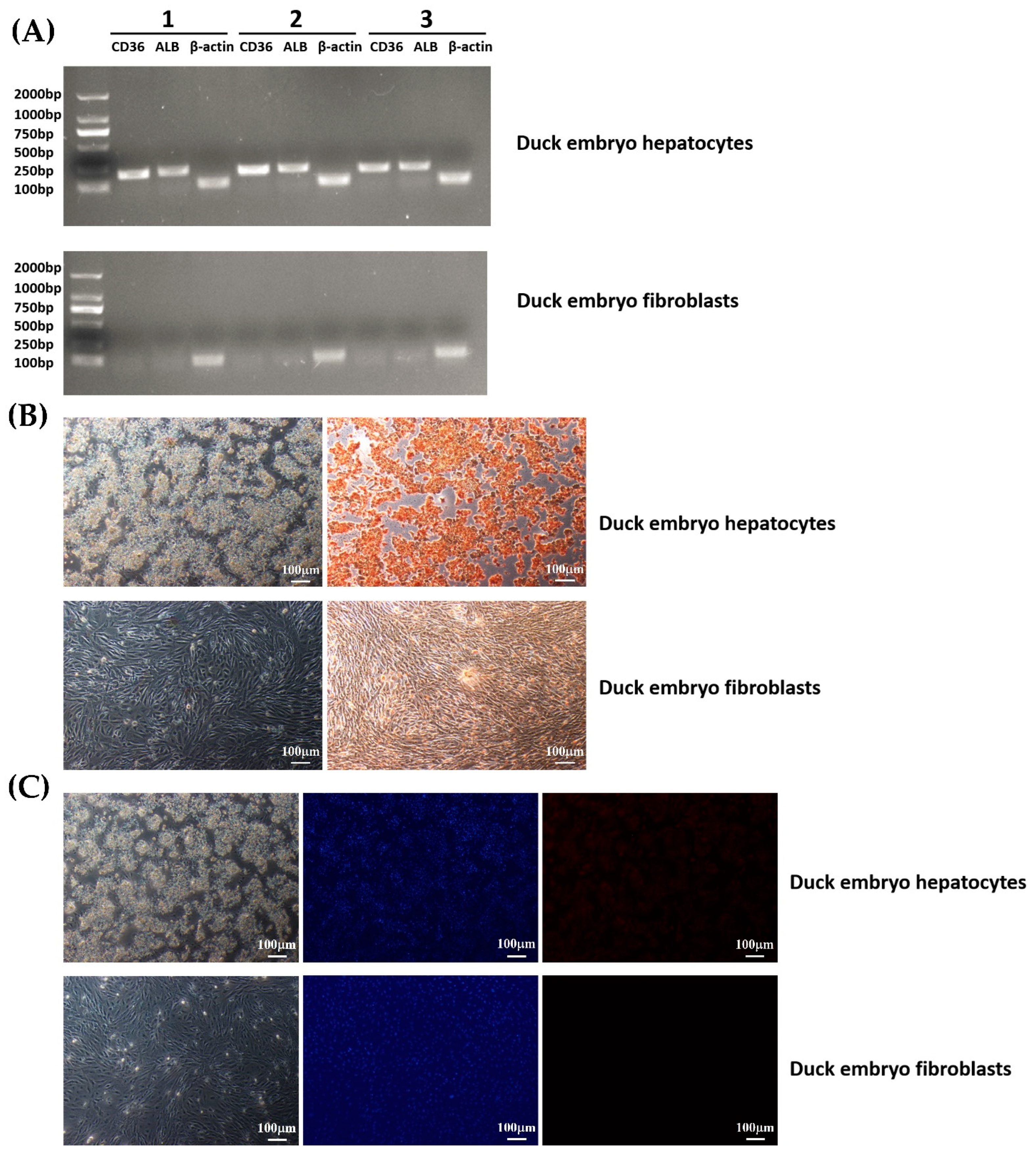
3.2. Morphological Observation and Growth Curve of Cells
3.3. Detection of Glycogen in Duck Embryo Hepatocytes
3.4. Expression Analysis of Function-Specific Genes of Duck Hepatocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Ocak, N.; Sivri, F. Liver colourations as well as performance and digestive tract characteristics of broilers may change as influenced by stage and schedule of feed restriction. J. Anim. Physiol. Anim. Nutr. 2008, 92, 546–553. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Fang, Z.P.; Liu, H.J.; Wang, L.J.; Cheng, Z.; Tang, N.; Li, T.; Liu, T.; Han, H.X.; Cao, G.; et al. HGF/R-spondin1 rescues liver dysfunction through the induction of Lgr5(+) liver stem cells. Nat. Commun. 2017, 8, 1175. [Google Scholar] [CrossRef] [PubMed]
- Hermier, D. Lipoprotein Metabolism and Fattening in Poultry. J. Nutr. 1997, 127, 805S–808S. [Google Scholar] [CrossRef] [PubMed]
- Tareen, S.H.K.; Kutmon, M.; Adriaens, M.E.; Mariman, E.C.M.; de Kok, T.M.; Arts, I.C.W.; Evelo, C.T. Exploring the cellular network of metabolic flexibility in the adipose tissue. Genes Nutr. 2018, 13, 17. [Google Scholar] [CrossRef] [PubMed]
- Kalmar, I.D.; Cools, A.; Verstegen, M.W.; Huyghebaert, G.; Buyse, J.; Roose, P.; Janssens, G.P. Dietary supplementation with dimethylglycine affects broiler performance and plasma metabolites depending on dose and dietary fatty acid profile. J. Anim. Physiol. Anim. Nutr. 2011, 95, 146–153. [Google Scholar] [CrossRef]
- Claire D’Andre, H.; Paul, W.; Shen, X.; Jia, X.; Zhang, R.; Sun, L.; Zhang, X. Identification and characterization of genes that control fat deposition in chickens. J. Anim. Sci. Biotechnol. 2013, 4, 43. [Google Scholar] [CrossRef]
- Jiang, R.R.; Zhao, G.P.; Zhao, J.P.; Chen, J.L.; Zheng, M.Q.; Liu, R.R.; Wen, J. Influence of dietary nicotinic acid supplementation on lipid metabolism and related gene expression in two distinct broiler breeds of female chickens. J. Anim. Physiol. Anim. Nutr. 2014, 98, 822–829. [Google Scholar] [CrossRef]
- Liu, W.M.; Lai, S.J.; Lu, L.Z.; Shi, F.X.; Zhang, J.; Liu, Y.; Yu, B.; Tao, Z.R.; Shen, J.D.; Li, G.Q.; et al. Effect of dietary fatty acids on serum parameters, fatty acid compositions, and liver histology in Shaoxing laying ducks. J. Zhejiang Univ. Sci. 2011, 12, 736–743. [Google Scholar] [CrossRef]
- Richards, M.P.; Poch, S.M.; Coon, C.N.; Rosebrough, R.W.; Ashwell, C.M.; McMurtry, J.P. Feed restriction significantly alters lipogenic gene expression in broiler breeder chickens. J. Nutr. 2003, 133, 707–715. [Google Scholar] [CrossRef]
- Park, M.; Yoo, J.H.; Lee, Y.S.; Park, E.J.; Lee, H.J. Ameliorative effects of black ginseng on nonalcoholic fatty liver disease in free fatty acid-induced HepG2 cells and high-fat/high-fructose diet-fed mice. J. Ginseng Res. 2020, 44, 350–361. [Google Scholar] [CrossRef]
- Xu, X.; Chen, X.; Huang, Z.; Chen, D.; He, J.; Zheng, P.; Chen, H.; Luo, J.; Luo, Y.; Yu, B.; et al. Effects of Dietary Apple Polyphenols Supplementation on Hepatic Fat Deposition and Antioxidant Capacity in Finishing Pigs. Animals 2019, 9, 937. [Google Scholar] [CrossRef]
- Osman, R.H.; Liu, L.; Xia, L.; Zhao, X.; Wang, Q.; Sun, X.; Zhang, Y.; Yang, B.; Zheng, Y.; Gong, D.; et al. Fads1 and 2 are promoted to meet instant need for long-chain polyunsaturated fatty acids in goose fatty liver. Mol. Cell. Biochem. 2016, 418, 103–117. [Google Scholar] [CrossRef]
- Qiu, S.; Chen, J.; Bai, Y.; He, J.; Cao, H.; Che, Q.; Guo, J.; Su, Z. GOS Ameliorates Nonalcoholic Fatty Liver Disease Induced by High Fat and High Sugar Diet through Lipid Metabolism and Intestinal Microbes. Nutrients 2022, 14, 2749. [Google Scholar] [CrossRef]
- Liu, L.; Zhao, X.; Wang, Q.; Sun, X.; Xia, L.; Wang, Q.; Yang, B.; Zhang, Y.; Montgomery, S.; Meng, H.; et al. Prosteatotic and Protective Components in a Unique Model of Fatty Liver: Gut Microbiota and Suppressed Complement System. Sci. Rep. 2016, 6, 31763. [Google Scholar] [CrossRef]
- Zaefarian, F.; Abdollahi, M.R.; Cowieson, A.; Ravindran, V. Avian Liver: The Forgotten Organ. Animals 2019, 9, 63. [Google Scholar] [CrossRef]
- Tavernier, A.; Davail, S.; Ricaud, K.; Bernadet, M.D.; Gontier, K. Genes involved in the establishment of hepatic steatosis in Muscovy, Pekin and mule ducks. Mol. Cell. Biochem. 2017, 424, 147–161. [Google Scholar] [CrossRef]
- Wei, R.; Han, C.; Deng, D.; Ye, F.; Gan, X.; Liu, H.; Li, L.; Xu, H.; Wei, S. Research progress into the physiological changes in metabolic pathways in waterfowl with hepatic steatosis. Br. Poult. Sci. 2021, 62, 118–124. [Google Scholar] [CrossRef]
- Lu, L.; Chen, Y.; Wang, Z.; Li, X.; Chen, W.; Tao, Z.; Shen, J.; Tian, Y.; Wang, D.; Li, G.; et al. The goose genome sequence leads to insights into the evolution of waterfowl and susceptibility to fatty liver. Genome Biol. 2015, 16, 89. [Google Scholar] [CrossRef]
- Begum, S.; Joshi, M.; Ek, M.; Holgersson, J.; Kleman, M.I.; Sumitran-Holgersson, S. Characterization and engraftment of long-term serum-free human fetal liver cell cultures. Cytotherapy 2010, 12, 201–211. [Google Scholar] [CrossRef]
- Garcia-Martinez, R.; Caraceni, P.; Bernardi, M.; Gines, P.; Arroyo, V.; Jalan, R. Albumin: Pathophysiologic basis of its role in the treatment of cirrhosis and its complications. Hepatology 2013, 58, 1836–1846. [Google Scholar] [CrossRef]
- Ward, E.S.; Ober, R.J. Hepatic function of FcRn revealed: Implications for overcoming drug-mediated hepatotoxicity. Hepatology 2017, 66, 2083–2085. [Google Scholar] [CrossRef] [PubMed]
- Pimentel, C.F.; Jiang, Z.G.; Otsubo, T.; Feldbrügge, L.; Challies, T.L.; Nasser, I.; Robson, S.; Afdhal, N.; Lai, M. Poor Inter-test Reliability Between CK18 Kits as a Biomarker of NASH. Dig. Dis. Sci. 2016, 61, 905–912. [Google Scholar] [CrossRef] [PubMed]
- Fitzpatrick, E.; Mitry, R.R.; Quaglia, A.; Hussain, M.J.; Debruyne, R.; Dhawan, A. Serum levels of CK18 M30 and leptin are useful predictors of steatohepatitis and fibrosis in paediatric NAFLD. J. Pediatr. Gastroenterol. Nutr. 2010, 51, 500–506. [Google Scholar] [CrossRef] [PubMed]
- Lebeau, P.F.; Byun, J.H.; Platko, K.; Al-Hashimi, A.A.; Lhoták, Š.; MacDonald, M.E.; Mejia-Benitez, A.; Prat, A.; Igdoura, S.A.; Trigatti, B.; et al. Pcsk9 knockout exacerbates diet-induced non-alcoholic steatohepatitis, fibrosis and liver injury in mice. JHEP Rep. Innov. Hepatol. 2019, 1, 418–429. [Google Scholar] [CrossRef]
- Holleboom, A.G.; Vergeer, M.; Hovingh, G.K.; Kastelein, J.J.; Kuivenhoven, J.A. The value of HDL genetics. Curr. Opin. Lipidol. 2008, 19, 385–394. [Google Scholar] [CrossRef]
- Unno, Y.; Sakai, M.; Sakamoto, Y.; Kuniyasu, A.; Nagai, R.; Nakayama, H.; Horiuchi, S. Glycolaldehyde-modified bovine serum albumin downregulates leptin expression in mouse adipocytes via a CD36-mediated pathway. Ann. N. Y. Acad. Sci. 2005, 1043, 696–701. [Google Scholar] [CrossRef]
- Alpini, G.; Phillips, J.O.; Vroman, B.; Larusso, N.F. Recent advances in the isolation of liver cells. Hepatology 2010, 20, 494–514. [Google Scholar] [CrossRef]
- Howard, R.B.; Christensen, A.K.; Gibbs, F.A.; Pesch, L.A. The enzymatic preparation of isolated intact parenchymal cells from rat liver. J. Cell Biol. 1967, 35, 675–684. [Google Scholar] [CrossRef]
- Seglen, P.O. Preparation of isolated rat liver cells. Methods Cell Biol. 1976, 13, 29–83. [Google Scholar] [CrossRef]
- Zhai, X.; Wang, W.; Dou, D.; Ma, Y.; Gang, D.; Jiang, Z.; Shi, B.; Jin, B. A novel technique to prepare a single cell suspension of isolated quiescent human hepatic stellate cells. Sci. Rep. 2019, 9, 12757. [Google Scholar] [CrossRef]
- Wang, X.; Yan, P.; Liu, L.; Luo, Y.; Zhao, L.; Liu, H.; Tang, Q.; Long, K.; Jin, L.; Ma, J.; et al. MicroRNA expression profiling reveals potential roles for microRNA in the liver during pigeon (Columba livia) development. Poult. Sci. 2020, 99, 6378–6389. [Google Scholar] [CrossRef]
- Dere, E.; Boverhof, D.R.; Burgoon, L.D.; Zacharewski, T.R. In vivo-in vitro toxicogenomic comparison of TCDD-elicited gene expression in Hepa1c1c7 mouse hepatoma cells and C57BL/6 hepatic tissue. BMC Genom. 2006, 7, 80. [Google Scholar] [CrossRef]
- Rachdaoui, N.; Sarkar, D.K. Effects of alcohol on the endocrine system. Endocrinol. Metab. Clin. North Am. 2013, 42, 593–615. [Google Scholar] [CrossRef]
- Li, M.; Xu, Z.; Lu, W.; Wang, L.; Zhang, Y. Potential Pharmacokinetic Effect of Chicken Xenobiotic Receptor Activator on Sulfadiazine: Involvement of P-glycoprotein Induction. Antibiotics 2022, 11, 1005. [Google Scholar] [CrossRef]
- Mason, W.S.; Litwin, S.; Jilbert, A.R. Immune selection during chronic hepadnavirus infection. Hepatol. Int. 2008, 2, 3–16. [Google Scholar] [CrossRef]
- Shen, L.; Hillebrand, A.; Wang, D.Q.; Liu, M. Isolation and primary culture of rat hepatic cells. J. Vis. Exp. JoVE 2012, 29, 3917. [Google Scholar] [CrossRef]
- Inzaugarat, M.E.; De Matteo, E.; Baz, P.; Lucero, D.; García, C.C.; Gonzalez Ballerga, E.; Daruich, J.; Sorda, J.A.; Wald, M.R.; Cherñavsky, A.C. New evidence for the therapeutic potential of curcumin to treat nonalcoholic fatty liver disease in humans. PLoS ONE 2017, 12, e0172900. [Google Scholar] [CrossRef]
- Lee, Y.S.; Eun, H.S.; Kim, S.Y.; Jeong, J.M.; Seo, W.; Byun, J.S.; Jeong, W.I.; Yi, H.S. Hepatic immunophenotyping for streptozotocin-induced hyperglycemia in mice. Sci. Rep. 2016, 6, 30656. [Google Scholar] [CrossRef]
- Rizvi, F.; Shukla, S.; Kakkar, P. Essential role of PH domain and leucine-rich repeat protein phosphatase 2 in Nrf2 suppression via modulation of Akt/GSK3β/Fyn kinase axis during oxidative hepatocellular toxicity. Cell Death Dis. 2014, 5, e1153. [Google Scholar] [CrossRef]
- Chen, X.; Cheng, L.; Lu, Y.; Yang, Z.; Lu, L. Paeoniflorin regulates macrophage activation in dimethylnitrosamine-induced liver fibrosis in rats. Bmc Complement. Altern. Med. 2012, 12, 254. [Google Scholar] [CrossRef]
- Okuyama, S.; Kawamura, F.; Kubiura, M.; Tsuji, S.; Osaki, M.; Kugoh, H.; Oshimura, M.; Kazuki, Y.; Tada, M. Real-time fluorometric evaluation of hepatoblast proliferation in vivo and in vitro using the expression of CYP3A7 coding for human fetus-specific P450. Pharmacol. Res. Perspect. 2020, 8, e00642. [Google Scholar] [CrossRef]
- Yeoh, G.C.; Bennett, F.A.; Oliver, I.T. Hepatocyte differentiation in culture. Appearance of tyrosine aminotransferase. Biochem. J. 1979, 180, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Kitani, H.; Yoshioka, M.; Takenouchi, T.; Sato, M.; Yamanaka, N. Isolation and characterization of macrophages from a mixed primary culture of bovine liver cells. Vet. Immunol. Immunopathol. 2011, 140, 341–345. [Google Scholar] [CrossRef]
- Zhang, M.; Tu, W.J.; Zhang, Q.; Wu, X.L.; Zou, X.Y.; Jiang, S. Osteocalcin reduces fat accumulation and inflammatory reaction by inhibiting ROS-JNK signal pathway in chicken embryonic hepatocytes. Poult. Sci. 2022, 101, 102026. [Google Scholar] [CrossRef]
- Li, H.; Li, Y.; Yang, L.; Zhang, D.; Liu, Z.; Wang, Y.; Han, R.; Li, G.; Li, Z.; Tian, Y.; et al. Identification of a Novel Lipid Metabolism-Associated Hepatic Gene Family Induced by Estrogen via ERα in Chicken (Gallus gallus). Front. Genet. 2020, 11, 271. [Google Scholar] [CrossRef] [PubMed]
- Nagaya, M.; Kubota, S.; Suzuki, N.; Akashi, K.; Mitaka, T. Thermoreversible gelation polymer induces the emergence of hepatic stem cells in the partially injured rat liver. Hepatology 2006, 43, 1053–1062. [Google Scholar] [CrossRef]
- Allende, D.S.; Gawrieh, S.; Cummings, O.W.; Belt, P.; Wilson, L.; Van Natta, M.; Behling, C.A.; Carpenter, D.; Gill, R.M.; Kleiner, D.E.; et al. Glycogenosis is common in nonalcoholic fatty liver disease and is independently associated with ballooning, but lower steatosis and lower fibrosis. Liver Int. Off. J. Int. Assoc. Study Liver 2021, 41, 996–1011. [Google Scholar] [CrossRef] [PubMed]
- Luan, Z.; Fan, X.; Song, H.; Li, R.; Zhang, W.; Zhang, J. Testosterone promotes GPX5 expression of goat epididymal epithelial cells cultured in vitro. Vitr. Cell. Dev. Biol. Anim. 2019, 55, 677–685. [Google Scholar] [CrossRef]
- Jia, H.; Liu, C.; Yang, Y.; Zhu, H.; Chen, F.; Liu, J.; Zhou, L. Inhibition of duck hepatitis B virus replication by mimic peptides in vitro. Exp. Ther. Med. 2015, 10, 1697–1703. [Google Scholar] [CrossRef]
- Lee, J.; Foster, D.N.; Bottje, W.G.; Jang, H.M.; Chandra, Y.G.; Gentles, L.E.; Kong, B.W. Establishment of an immortal chicken embryo liver-derived cell line. Poult. Sci. 2013, 92, 1604–1612. [Google Scholar] [CrossRef]
- Jacquinot, P.M.; Léger, D.; Wieruszeski, J.M.; Coddeville, B.; Montreuil, J.; Spik, G. Change in glycosylation of chicken transferrin glycans biosynthesized during embryogenesis and primary culture of embryo hepatocytes. Glycobiology 1994, 4, 617–624. [Google Scholar] [CrossRef]
- Schacke, M.; Glück, B.; Wutzler, P.; Sauerbrei, A. In vitro cultivation and cryopreservation of duck embryonic hepatocytes. J. Virol. Methods 2009, 157, 25–31. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Primers | Sequence (5′–3′) | Accession Number | Product Length (bp) |
|---|---|---|---|
| CD36-F | AGCCCAAATGAGAAGGAACA | XM_038183702.1 | 194 |
| CD36-R | GCATCCCCAATAACAGCAGA | ||
| ALB-F | ACTGCCCTCCATTTTCCTG | NM_001310394.1 | 202 |
| ALB-R | TCTGTTGTCCTCGTATTCCTTG | ||
| β-actin-F | ATGTCGCCCTGGATTTCG | EF667345.1 | 165 |
| β-actin-R | CACAGGACTCCATACCCAAGAA |
| Age | E14 | E21 | E27 | D7 | p Value |
|---|---|---|---|---|---|
| Length (cm) | 1.23 ± 0.07 a | 2.06 ± 0.07 b | 2.23 ± 0.07 c | 4.18 ± 0.11 d | 0.000 |
| Weight (g) | 0.22 ± 0.03 a | 0.64 ± 0.02 b | 1.07 ± 0.06 c | 5.19 ± 0.23 d | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, Q.; Wang, L.; Hu, X.; Yuan, C.; Zhang, Y.; Chang, G.; Chen, G. Developmental Changes of Duckling Liver and Isolation of Primary Hepatocytes. Animals 2023, 13, 1820. https://doi.org/10.3390/ani13111820
Bao Q, Wang L, Hu X, Yuan C, Zhang Y, Chang G, Chen G. Developmental Changes of Duckling Liver and Isolation of Primary Hepatocytes. Animals. 2023; 13(11):1820. https://doi.org/10.3390/ani13111820
Chicago/Turabian StyleBao, Qiang, Laidi Wang, Xiaodan Hu, Chunyou Yuan, Yang Zhang, Guobin Chang, and Guohong Chen. 2023. "Developmental Changes of Duckling Liver and Isolation of Primary Hepatocytes" Animals 13, no. 11: 1820. https://doi.org/10.3390/ani13111820
APA StyleBao, Q., Wang, L., Hu, X., Yuan, C., Zhang, Y., Chang, G., & Chen, G. (2023). Developmental Changes of Duckling Liver and Isolation of Primary Hepatocytes. Animals, 13(11), 1820. https://doi.org/10.3390/ani13111820