3.1. Description of Ultratenuipalpus parameekeri Castro, Ochoa & Feres sp. nov.
Family Tenuipalpidae Berlese, 1913
Genus Ultratenuipalpus Mitrofanov, 1973
Type species: Ultratenuipalpus meekeri (De Leon), 1957
Diagnosis of the genus (Based on [
2]). “Body shape from elongate-ovate to broadly rounded; broad propodosoma differentiated from narrower opisthosoma (although anterior opisthosoma is broad at junction with propodosoma). Anterior margin of prodorsum usually with median forked projection forming a short notch.” Prodorsum with one pair of lateral body projections anterior to setae
sc2 present or absent. Posterior margin of opisthosoma with a broad rounded projection between setae
h1 usually present. “Prodorsal shield divided by two oblique folds running from vicinity of the eyes angled medially to posterior margin of shield, superficially dividing the shield into three smaller plate-like regions; a small plate is indicated between setae
c3–
d3 on dorsal opisthosomal margin; posteroventral body margin often with band of pustulate cuticle. Dorsal opisthosoma with setae
c1,
c3,
d1,
d3,
e1,
e3,
f3,
h1,
h2 present (except setae
f3 absent in
U. aberrans); setae
f2 present or absent, when present then inserted on lateral margin aligned with lateral setae
e3,
f3,
h1,
h2; setae
c2,
d2,
e2 absent. Setae
h2 not flagellate, similar in form to
h1; setae
sc2,
e3,
f2,
f3,
h1,
h2 flattened, lanceolate, oblanceolate, obovate to ovate, with
sc2 often falcate; form of other dorsal setae variable (e.g.,
sc2 and
f3 flagellate in
U. bunyai). Three pairs of
ps (pseudanal) setae present; female with ps3 positioned anteriorly on anal valves and much shorter than
ps1–
2, which are closely associated with each other and positioned posterolaterad anal valves; setae
ps2 usually much longer than
ps1; male with ps3 modified into accessory genital stylets and inserted on elongate genitoanal valves, with
ps1–
2 positioned as in female. Ventral, genital and anal regions membranous, without defined sclerotized plates; flap of ovipore and anus surrounded by strongly plicate and wrinkled membranous cuticle. Genital setae
g2 inserted slightly anterior to
g1 on reduced genital flap;
g1–
2 often aligned longitudinally with setae
ag. Intercoxal setae (
3a,
4a) not multiplied. Palps four segmented; palp tibiae with 1–2 setae; palp tarsi with 1–3 phaneres, with solenidion always present, sometimes curved, often inserted basally on palp tarsus segment at junction with palp tibia. Dorsal setae on legs inserted in lateral position. Femora of legs I–II with four setae (
d,
l’,
v’,
bv″); genua I–II with three setae (
d,
l’,
l″) (except some species variously described with two setae—
U. acharis (genua I–II with 3–2 setae; possibly
d absent),
U. pterophilus (genua I–II with 2–2)); tibiae I–II usually with five setae (except two species described with four setae,
U. avarua (
v″ absent) and
U. lacorpuzrarosae (possibly
d absent)). Tarsal claws pad-like. Immature stages with setae
c1 inserted distinctly anterior to level of setae
c3. See also diagnosis of [
1].”
Description
Diagnosis. Female: As per genus, in addition to: prodorsal setae v2, sc1 minute to short, and sc2 large, flattened obovate to ovate; dorsal opisthosoma with 10 pairs of setae (f2 present); most of the dorsal opisthosomal setae large, flattened, obovate to ovate, except setae d3 is distinctly short and c3 almost orbicular; pair lateral body projections anterior to setae sc2 and projection between opisthosomal setae h1 both present; palp four segmented, setal formula 0, 0, 2, 2. Male: Opisthosoma narrower than that of the female, with distinct transverse constriction (waist) between setae d1 and e1; many dorsal setae similar in form to those of females, except c1, d1, and e1 short to minute, d3 longer, and setae along posterior margin of opisthosoma (especially e3) narrower and more elongated than those of the female. Tarsi I–II each with two solenidia (ω′ paraxial and ventrolateral; ω” antiaxial); tarsus III with one solenidion ω′ paraxial and ventrolateral. Immatures: with lateral body projection anterior to setae sc2 present (except absent in larvae); posterior projection between setae h1 absent; dorsal setae similar in general form to those of the female, except setae c1, d1, and e1 short to minute. Larvae with central prodorsum and pygidial region of posterior opisthosoma with finely colliculated integument.
Body measurements: distance between setae v2–h1 350 (340–375), sc2–sc2 220 (215–230); other measurements: v2–v2 45 (37–45), sc1–sc1 93 (90–110), c1–c1 60 (55–63), c3–c3 260 (245–265), d1–d1 40 (37–45), d3–d3 240 (230–240), e1–e1 32 (27–33), e3–e3 220 (215–225), f2–f2 205 (205–215), f3–f3 170 (170–185), h1–h1 58 (58–68), h2–h2 120 (115–135).
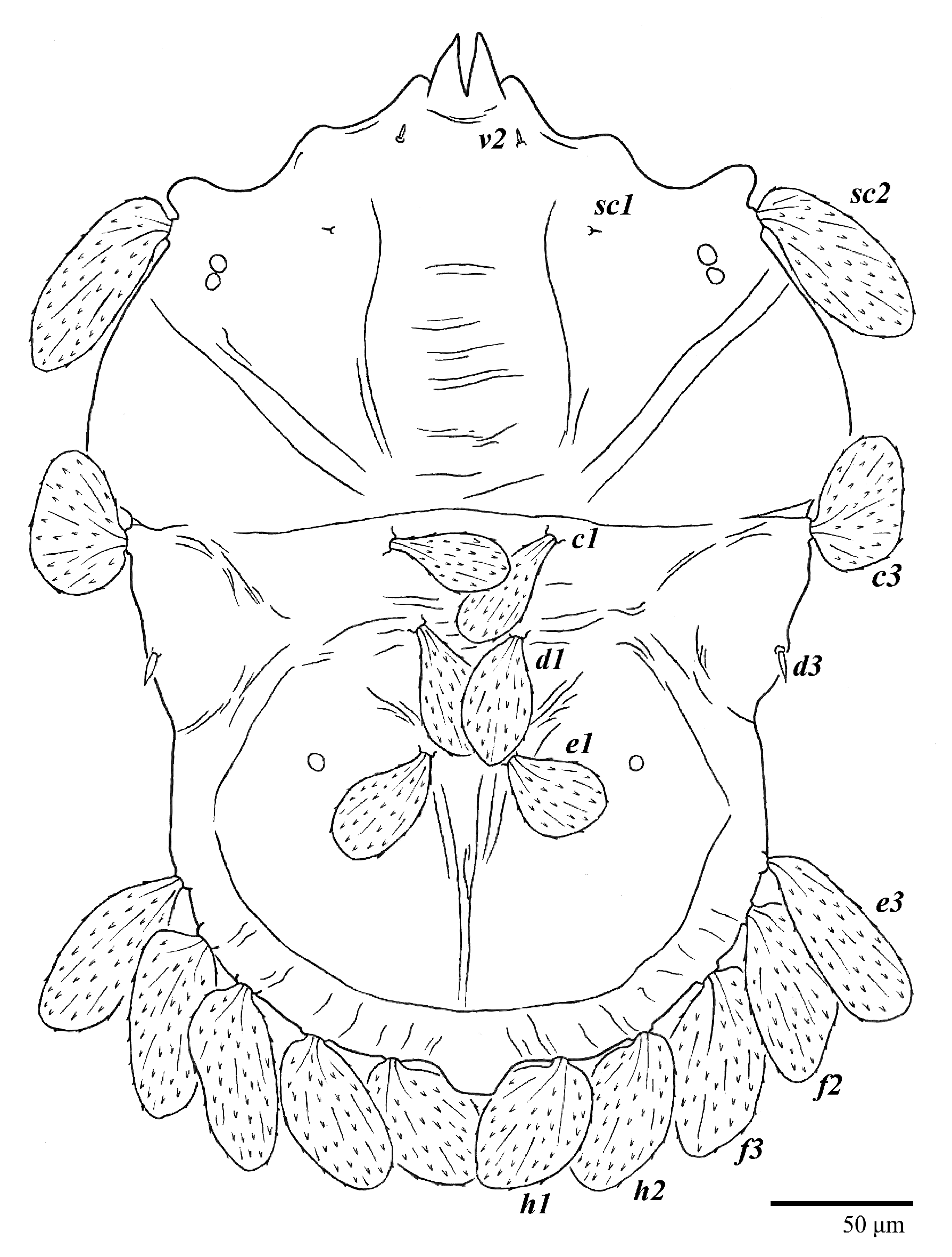
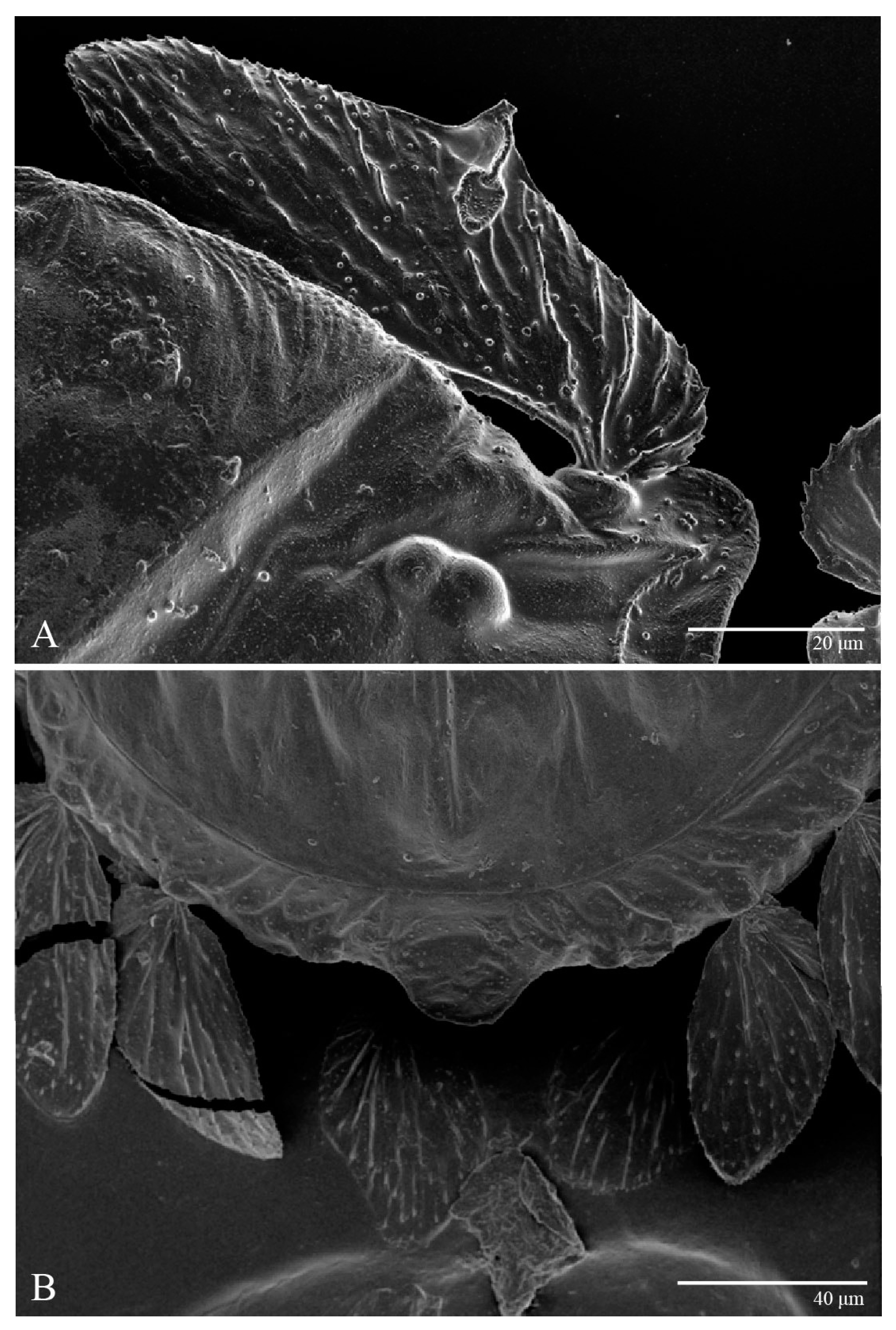
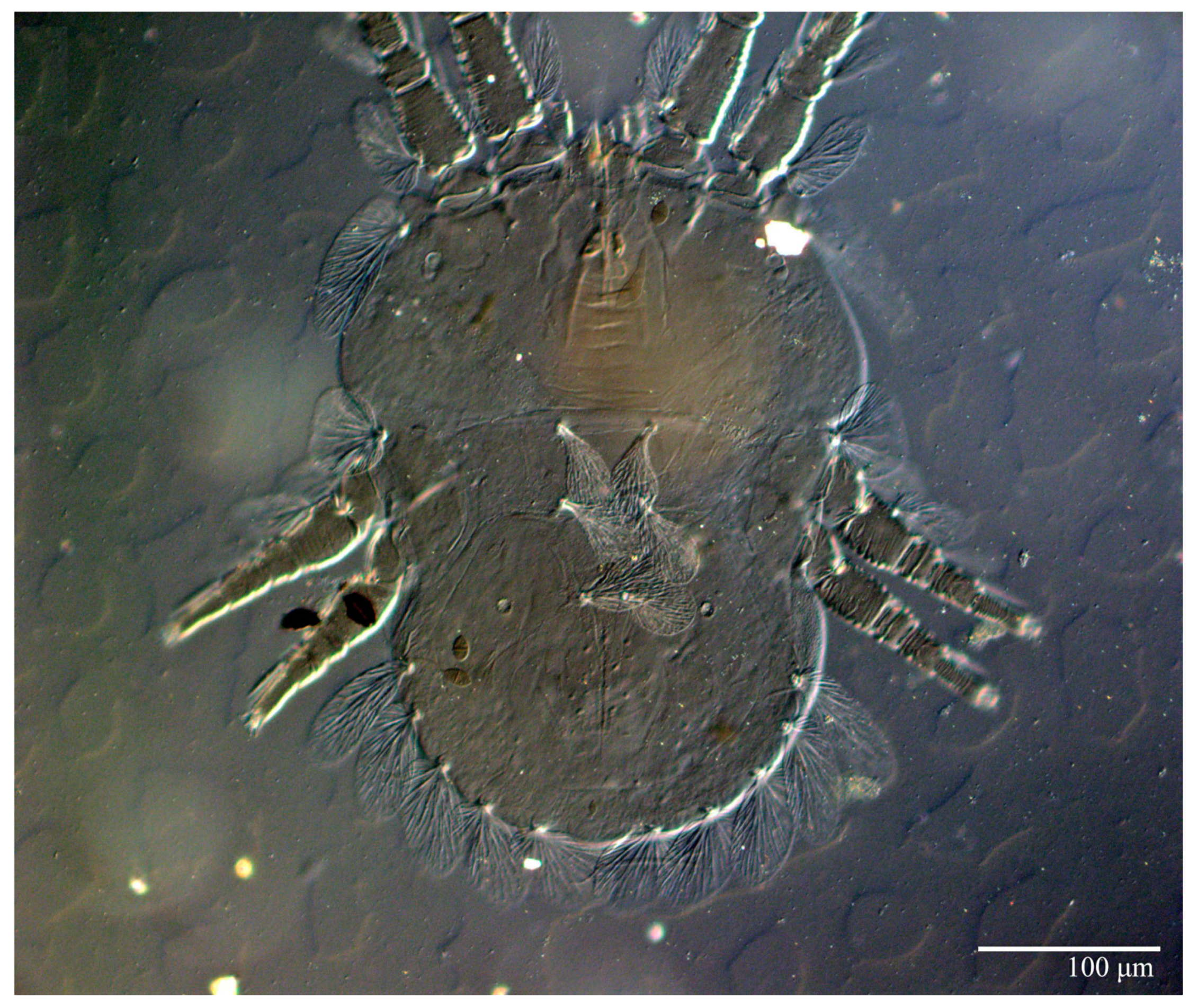
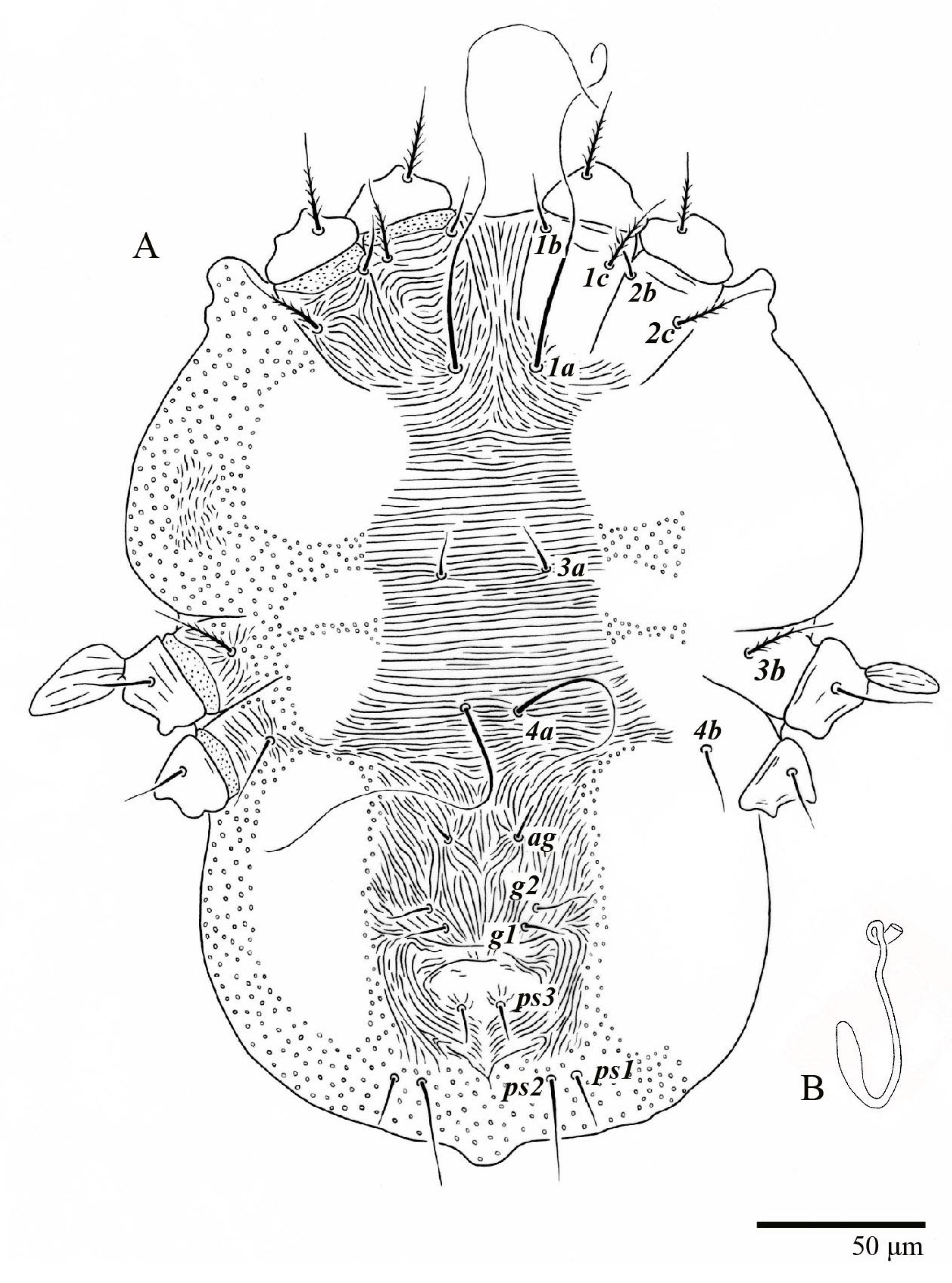
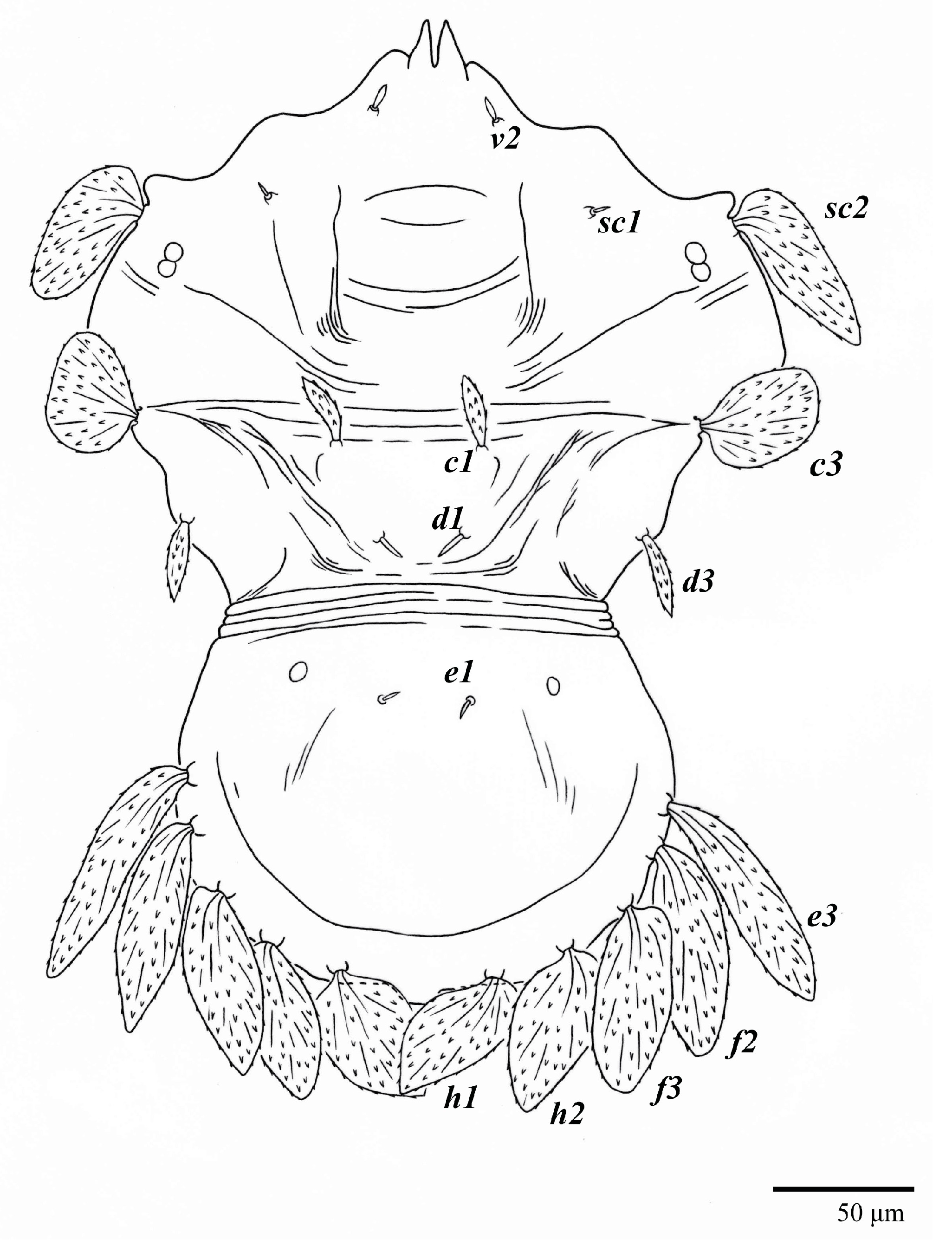
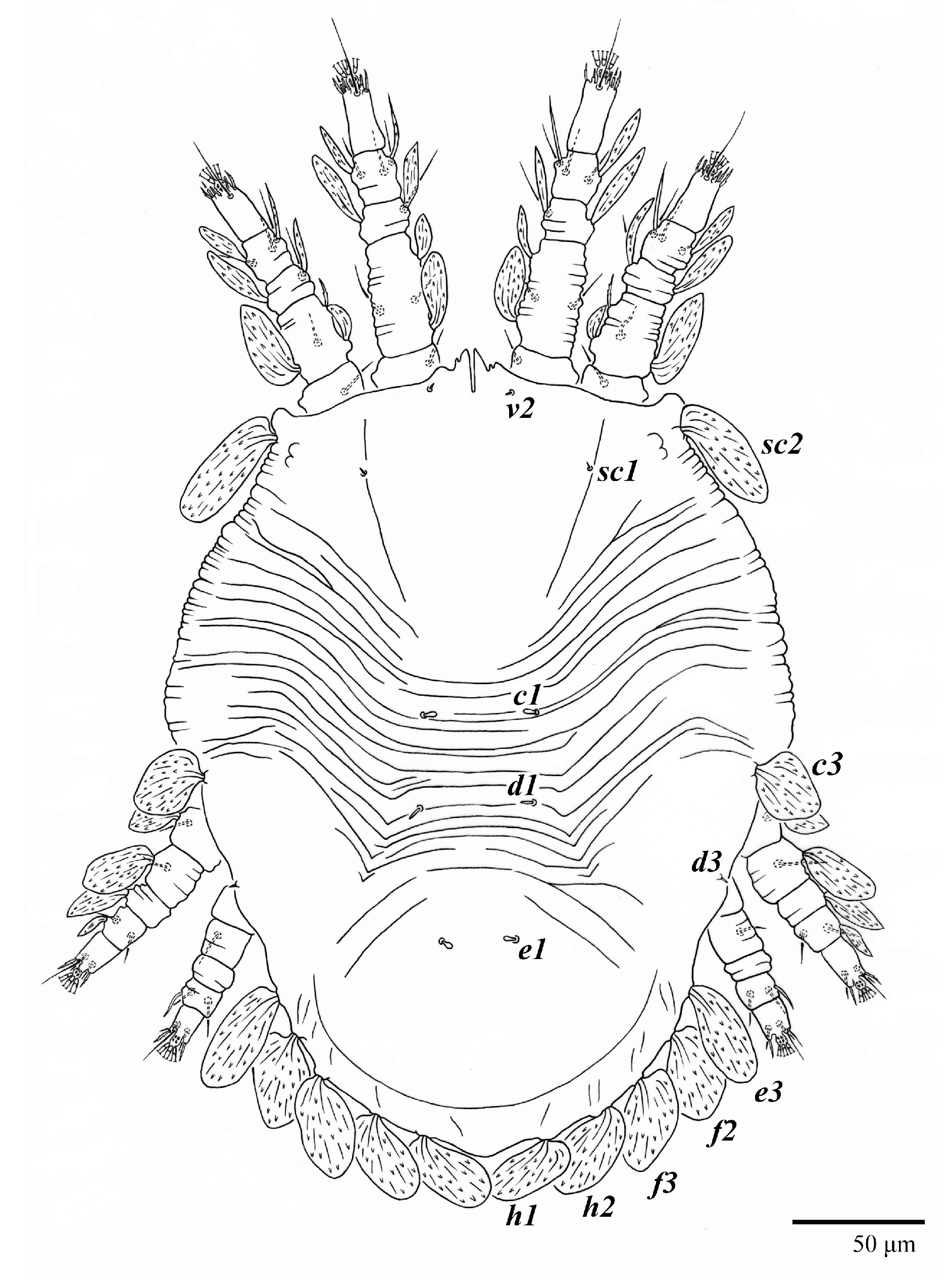
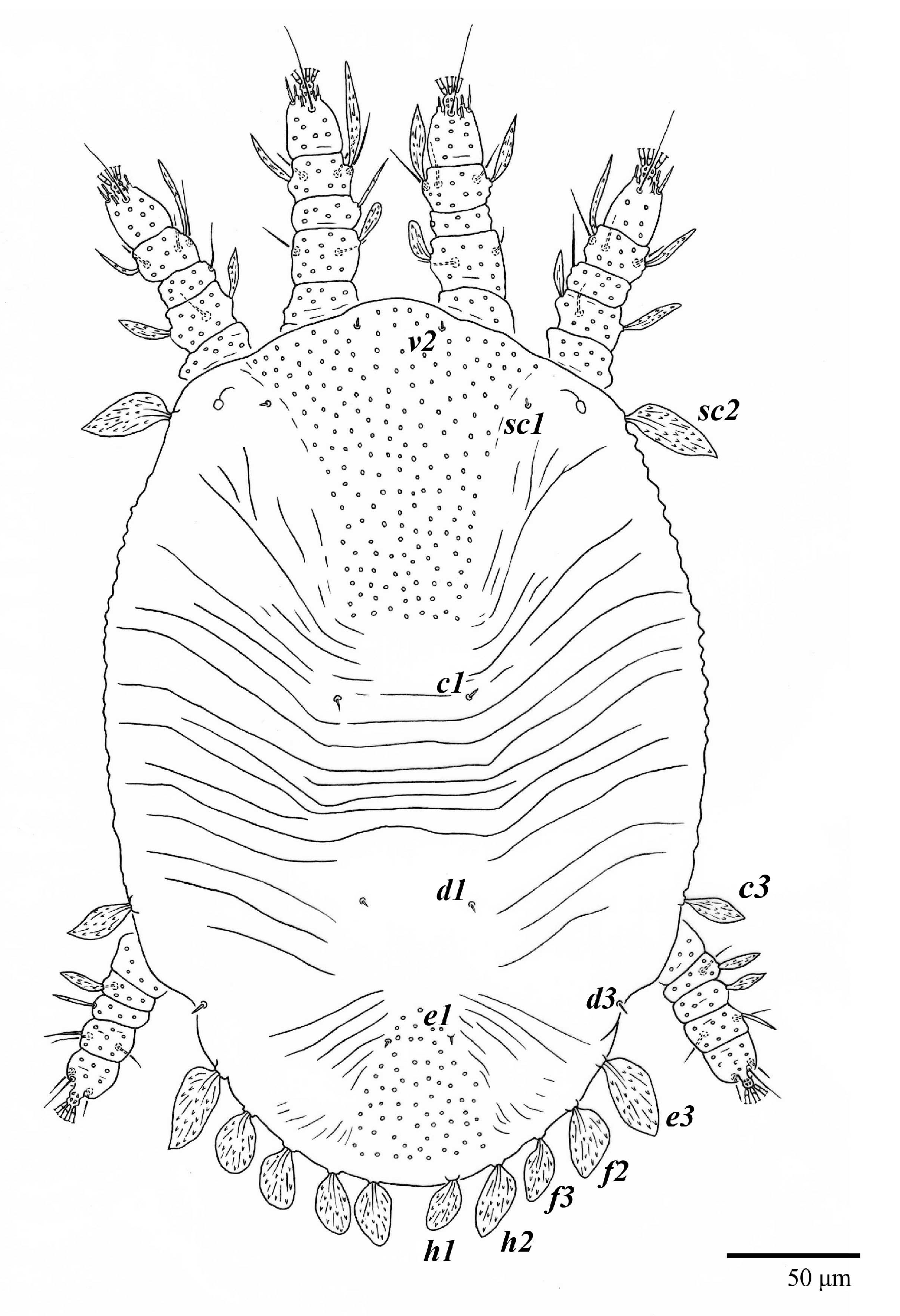
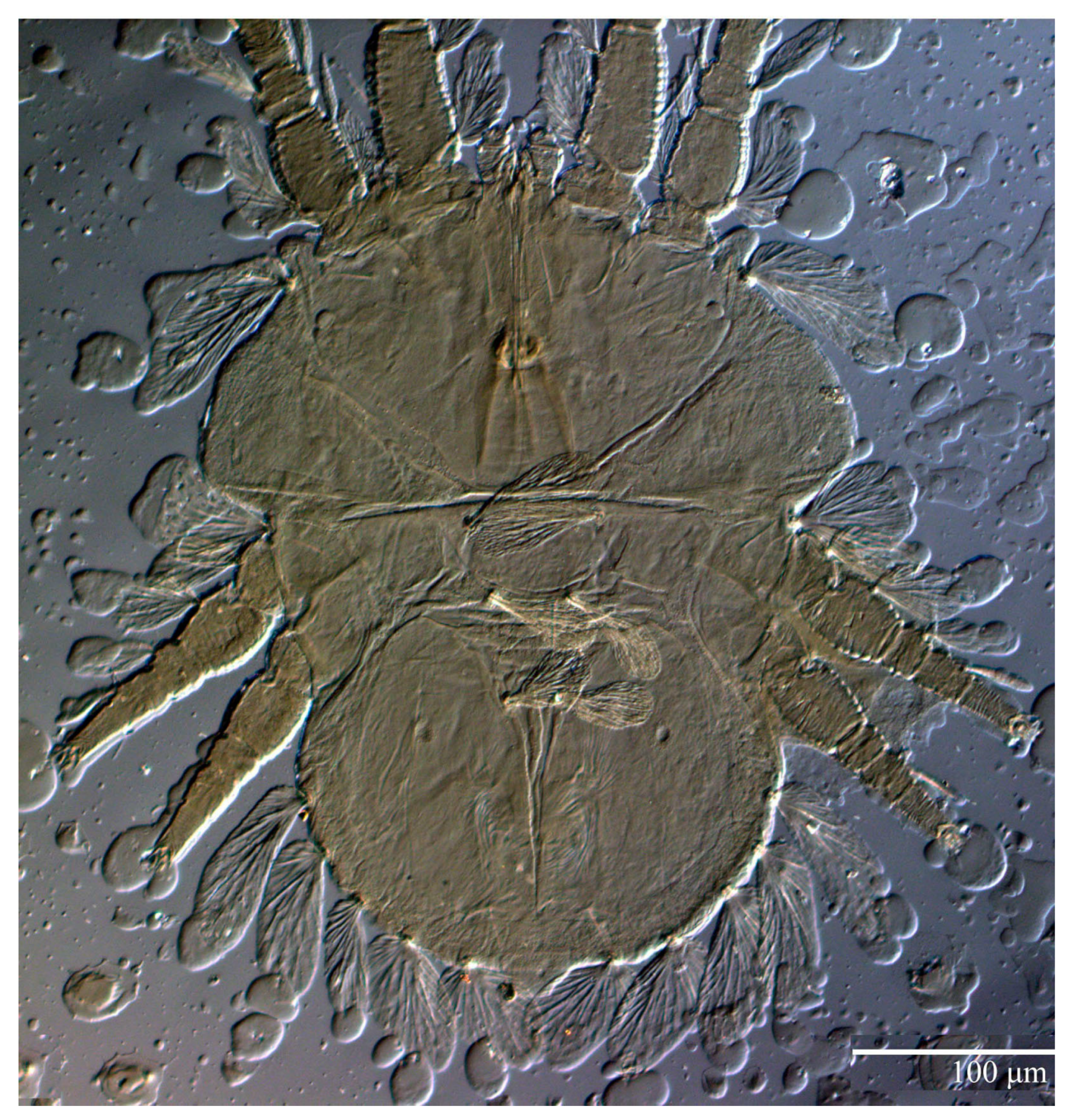
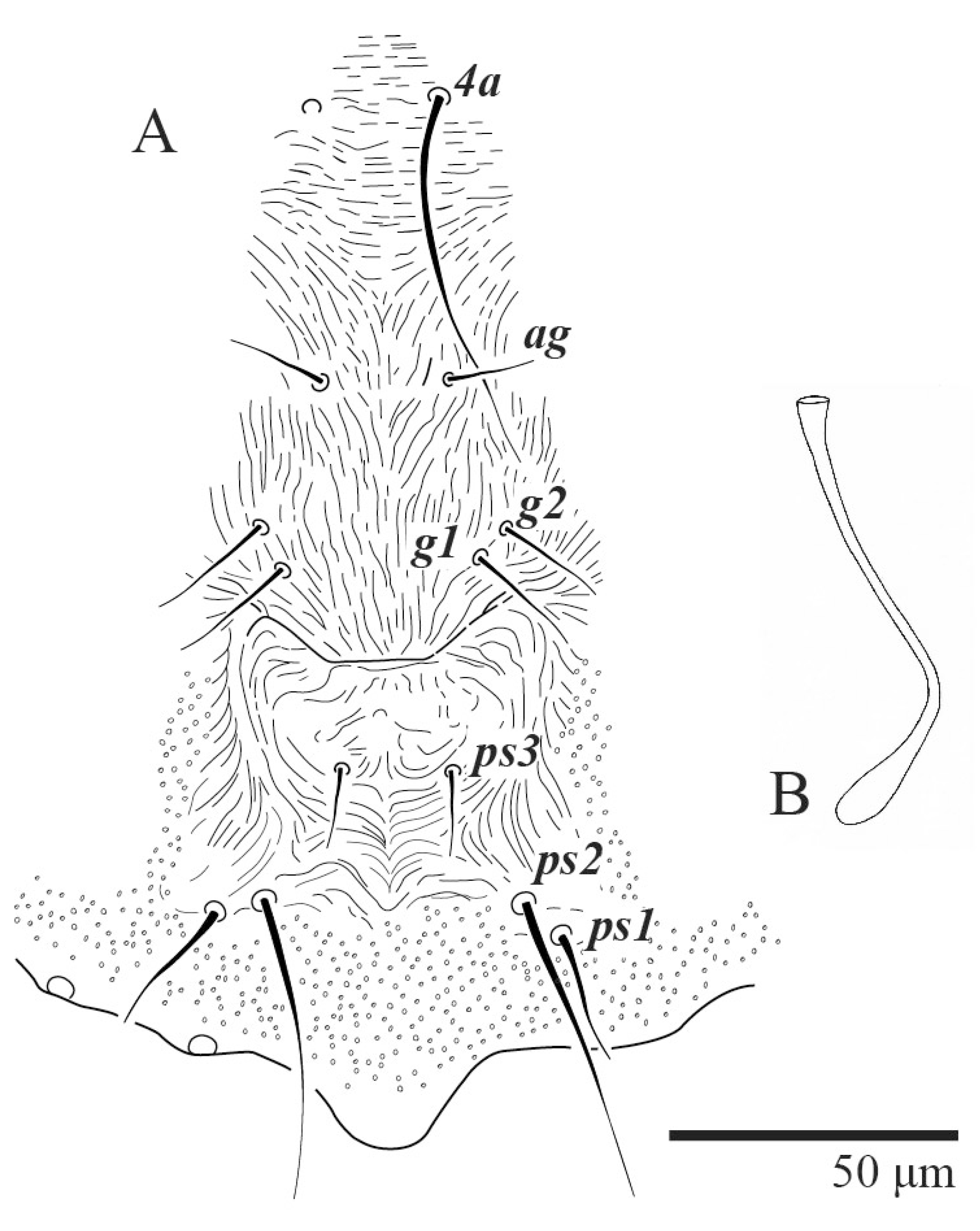
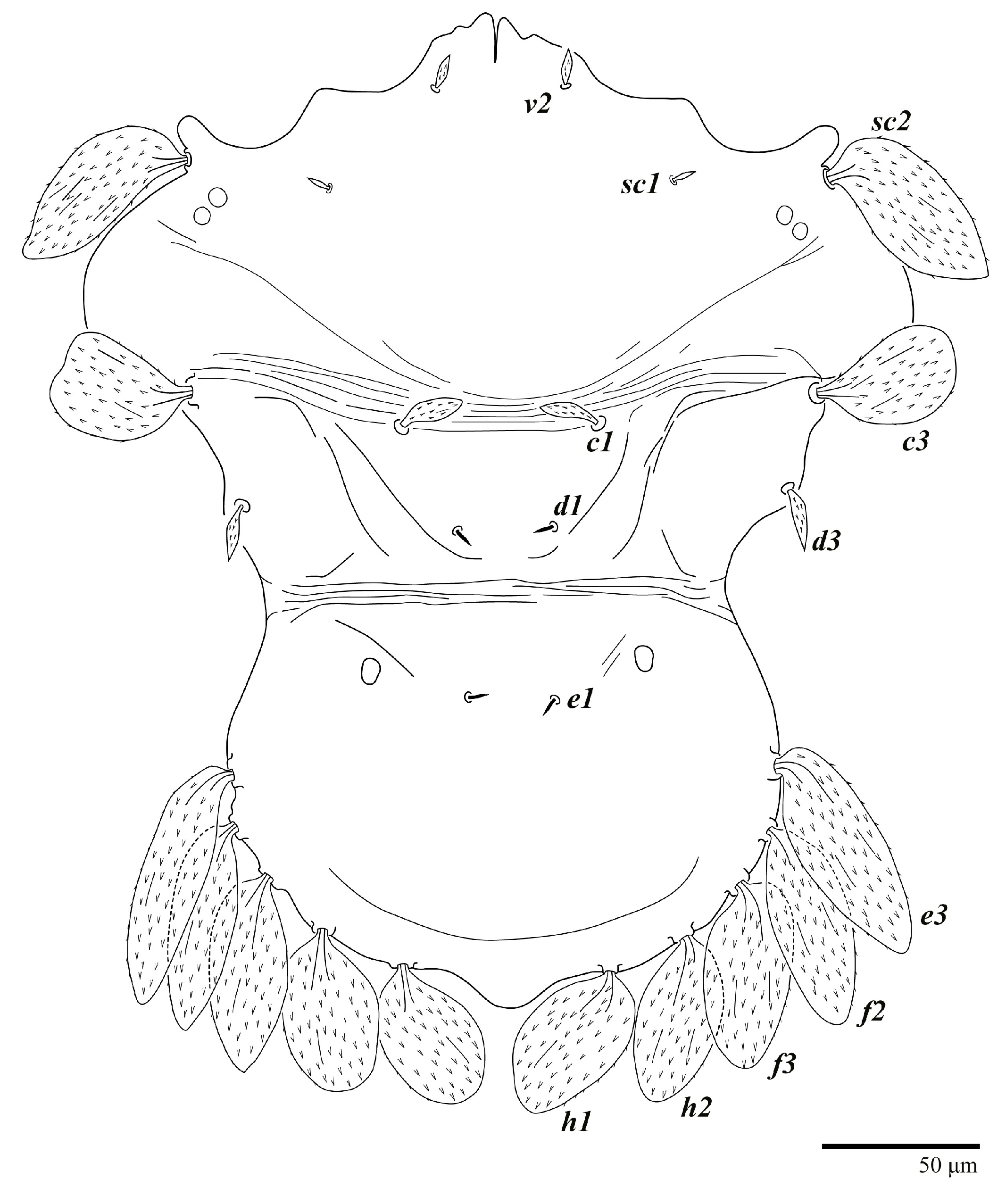
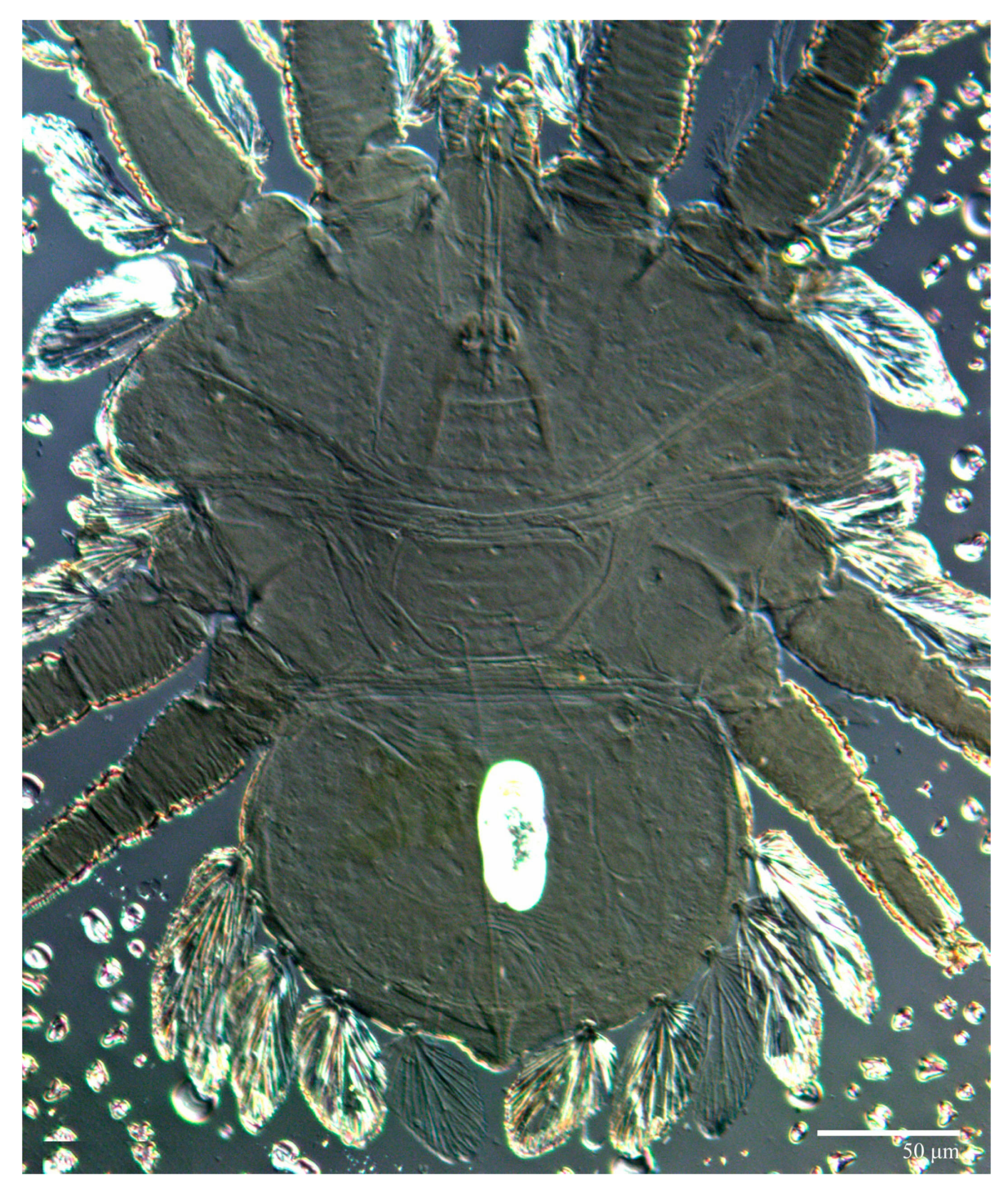
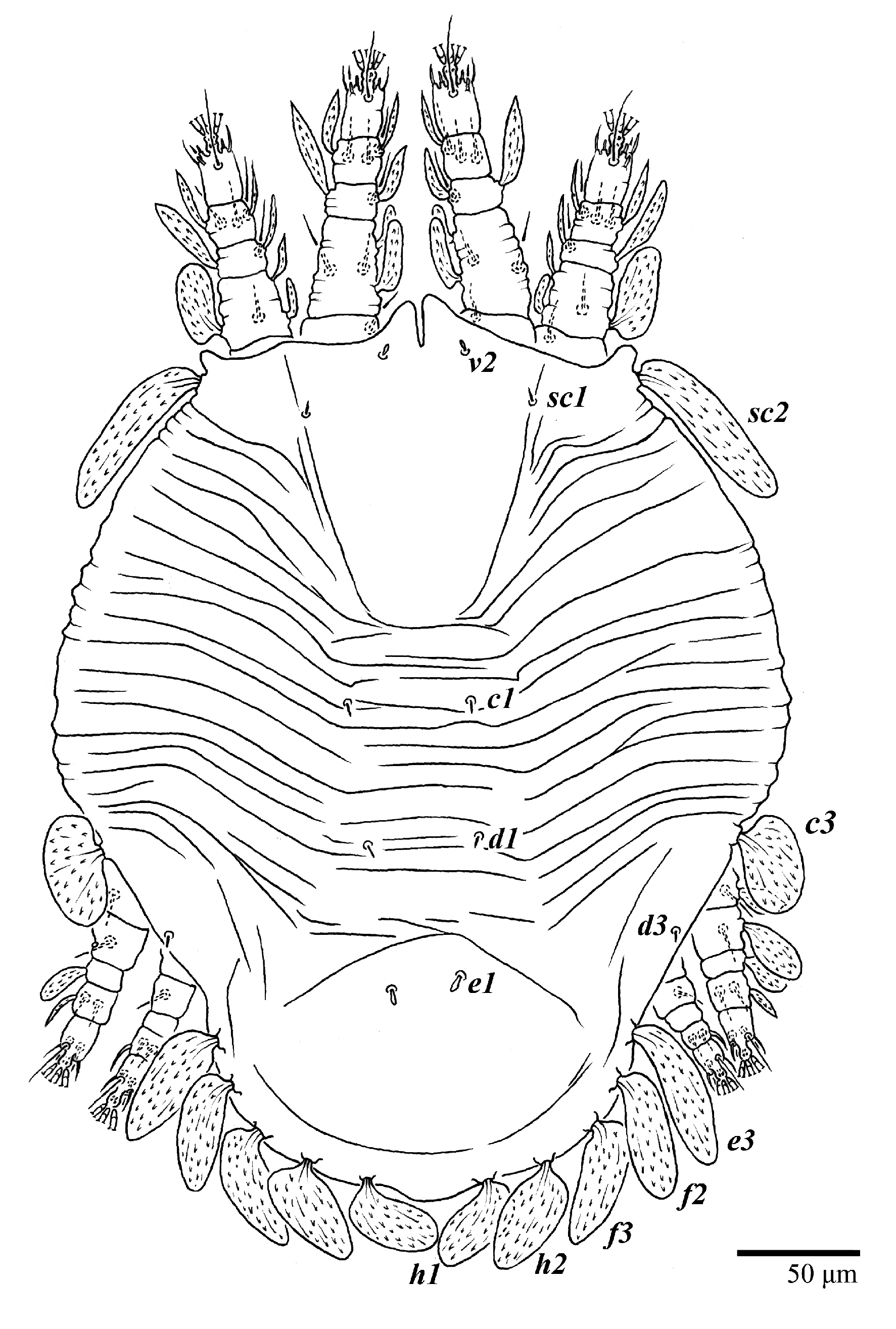
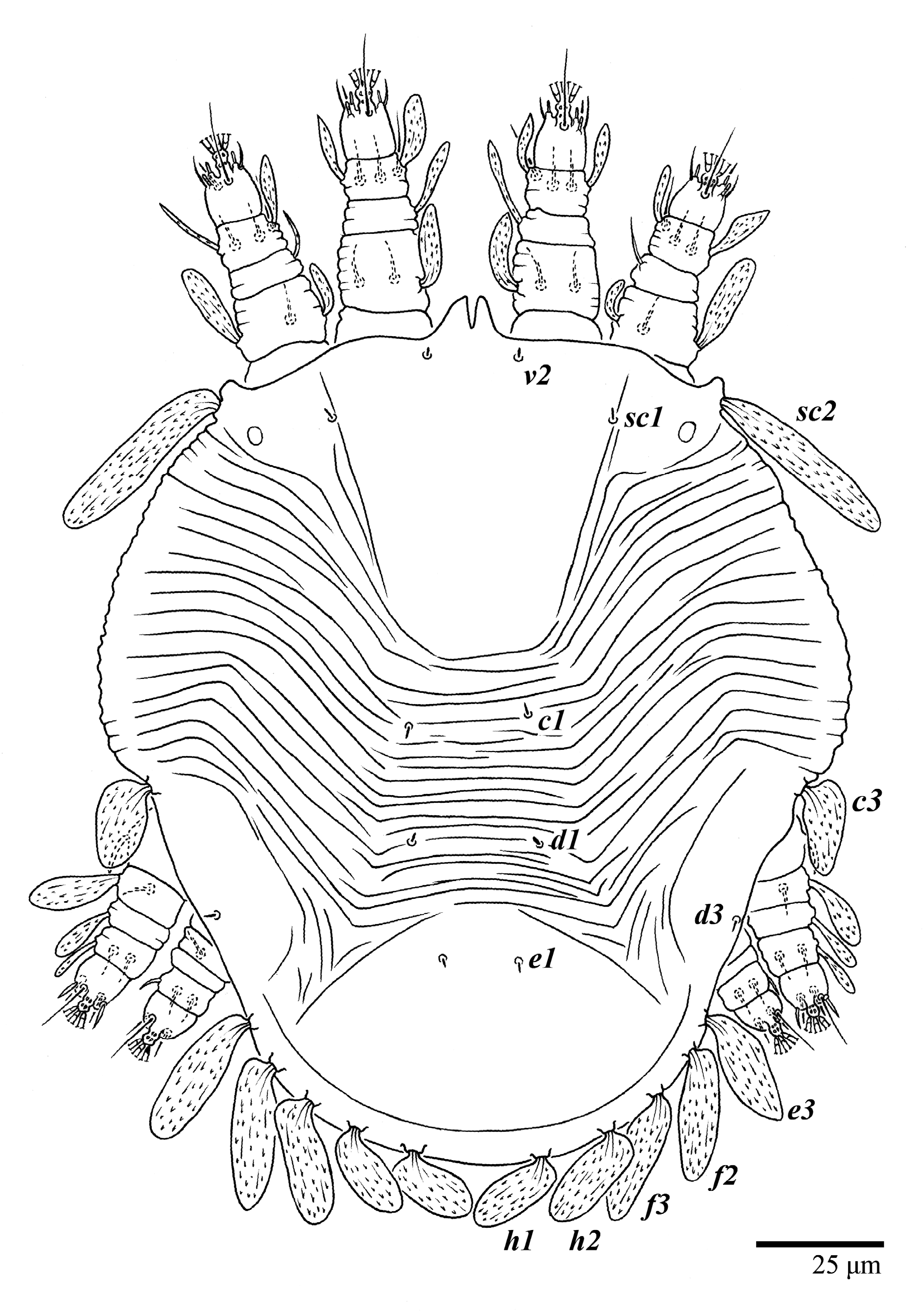
Dorsum (
Figure 1,
Figure 2,
Figure 3 and
Figure 4). Anterior margin of prodorsum with a short median forked projection forming a short notch 27 (20–27). Dorsum smooth, with pair lateral projections anterior to setae
sc2 and single projection between opisthosomal setae
h1 (
Figure 3B). A pair of converging folds from the eyes to near the sejugal furrow on the prodorsum posterior margin. Prodorsal setae
v2 and
sc1 short to minute;
sc2 large, flattened elongate obovate (
Figure 1 and
Figure 3A); most opisthosomal setae similar to prodorsal setae
sc2, except
d3 is short. Setal measurements:
v2 5 (4–7),
sc1 3 (3–5),
sc2 74 (74–82),
c1 52 (52–58),
c3 36 (36–45),
d1 55 (54–55),
d3 10 (8–10),
e1 50 (48–55),
e3 70 (70–81),
f2 65 (65–70),
f3 61 (60–68),
h1 52 (52–60),
h2 58 (58–65).
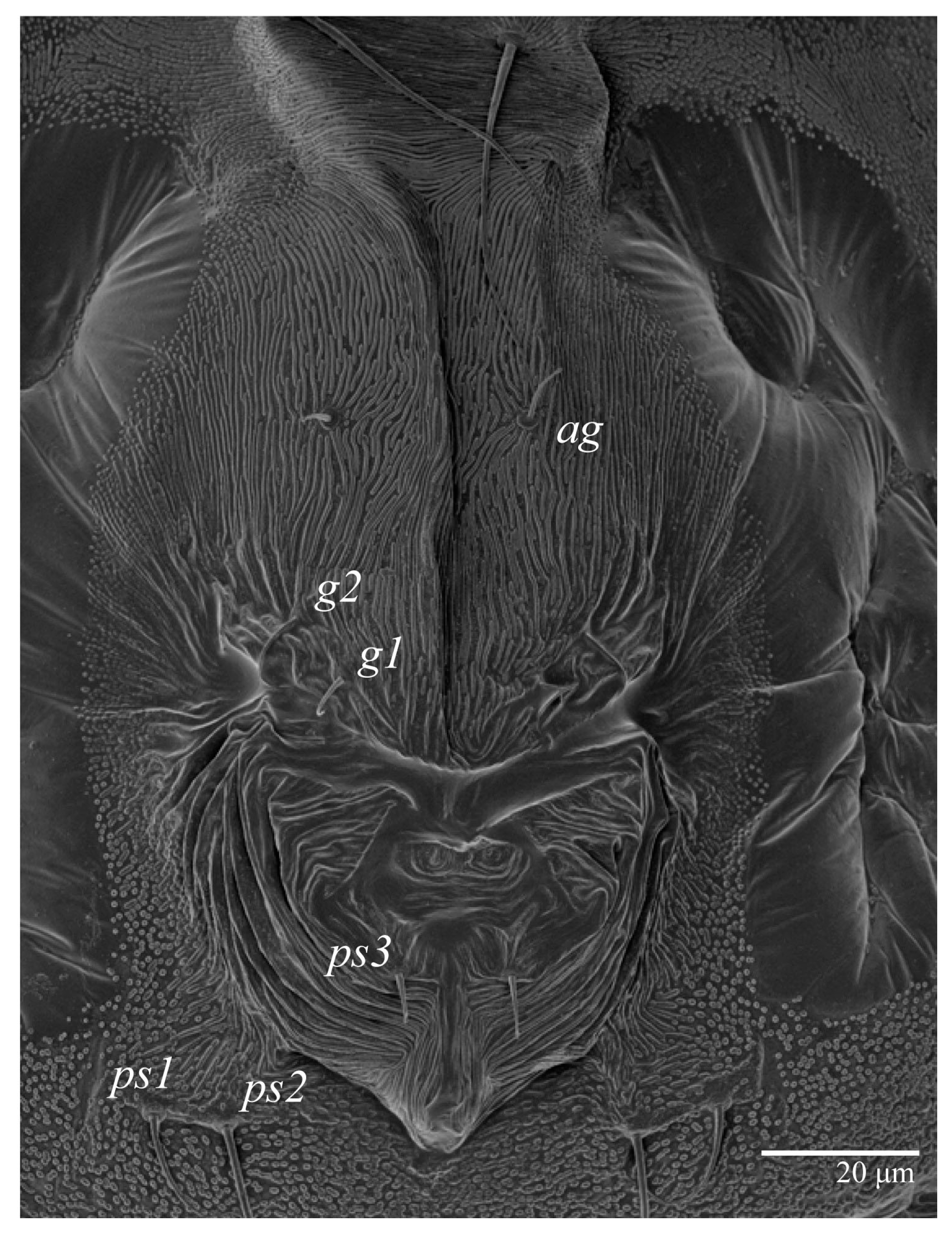
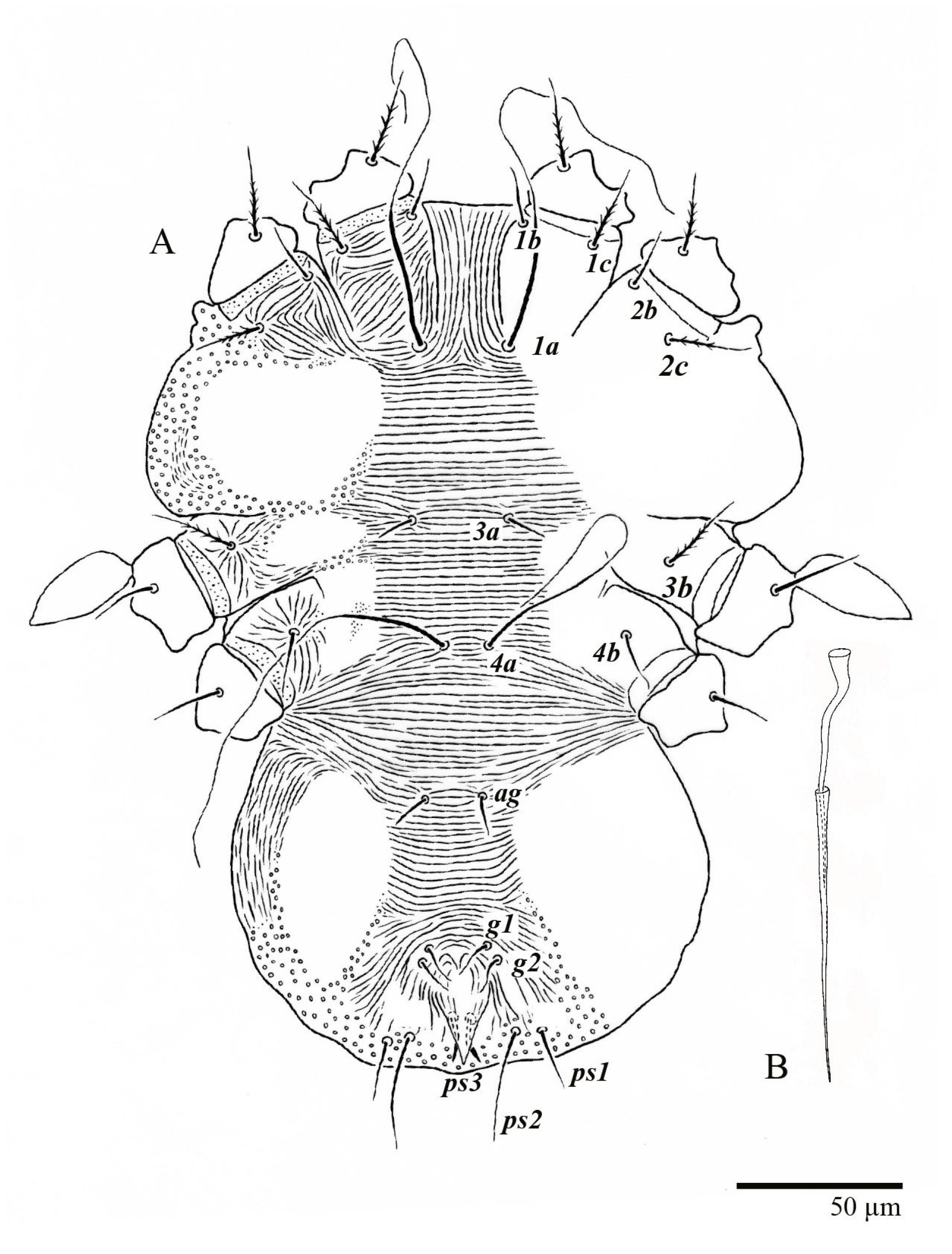
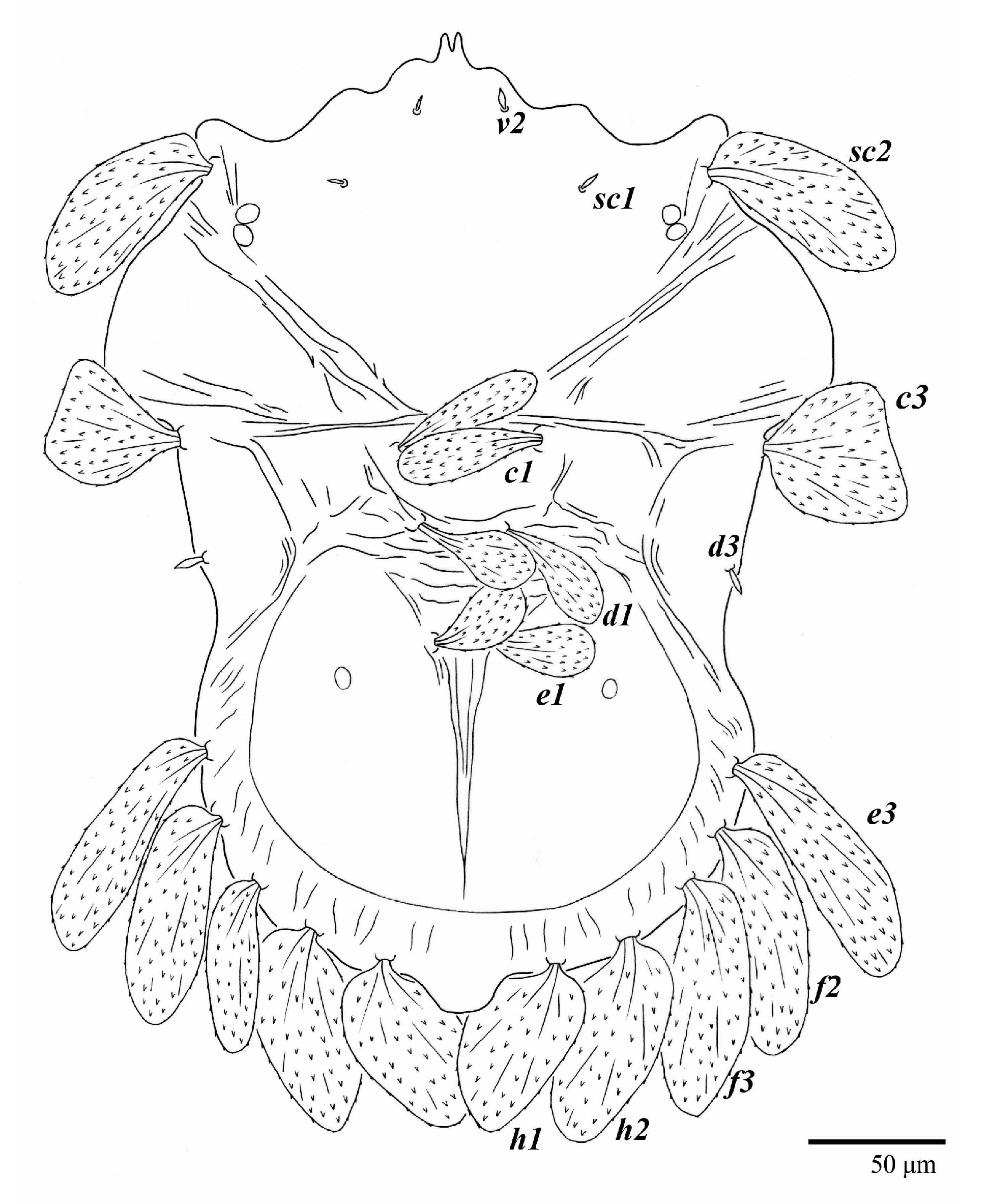
Venter (
Figure 5A and
Figure 6). Ventral integument weakly striate along central region and densely colliculated around lateral body margin; ventral, genital, and anal plates not developed, and entire region membranous and distinctly plicate; ventral setae filiform, with coxal setae
1c,
2c, and
3b barbed; setae
ps2 distinctly longer than
ps1. Setal measurements:
1a 105 (100–135),
1b 13 (13–16),
1c 30 (25–30),
2b 22 (22–26),
2c 38 (30–39),
3a 18 (15–18),
3b 32 (31–36),
4a 145 (115–145),
4b 22 (18–22),
ag 10 (10–11),
g1 16 (14–16),
g2 16 (15–18),
ps1 28 (22–28),
ps2 53 (50–55),
ps3 12 (10–13).
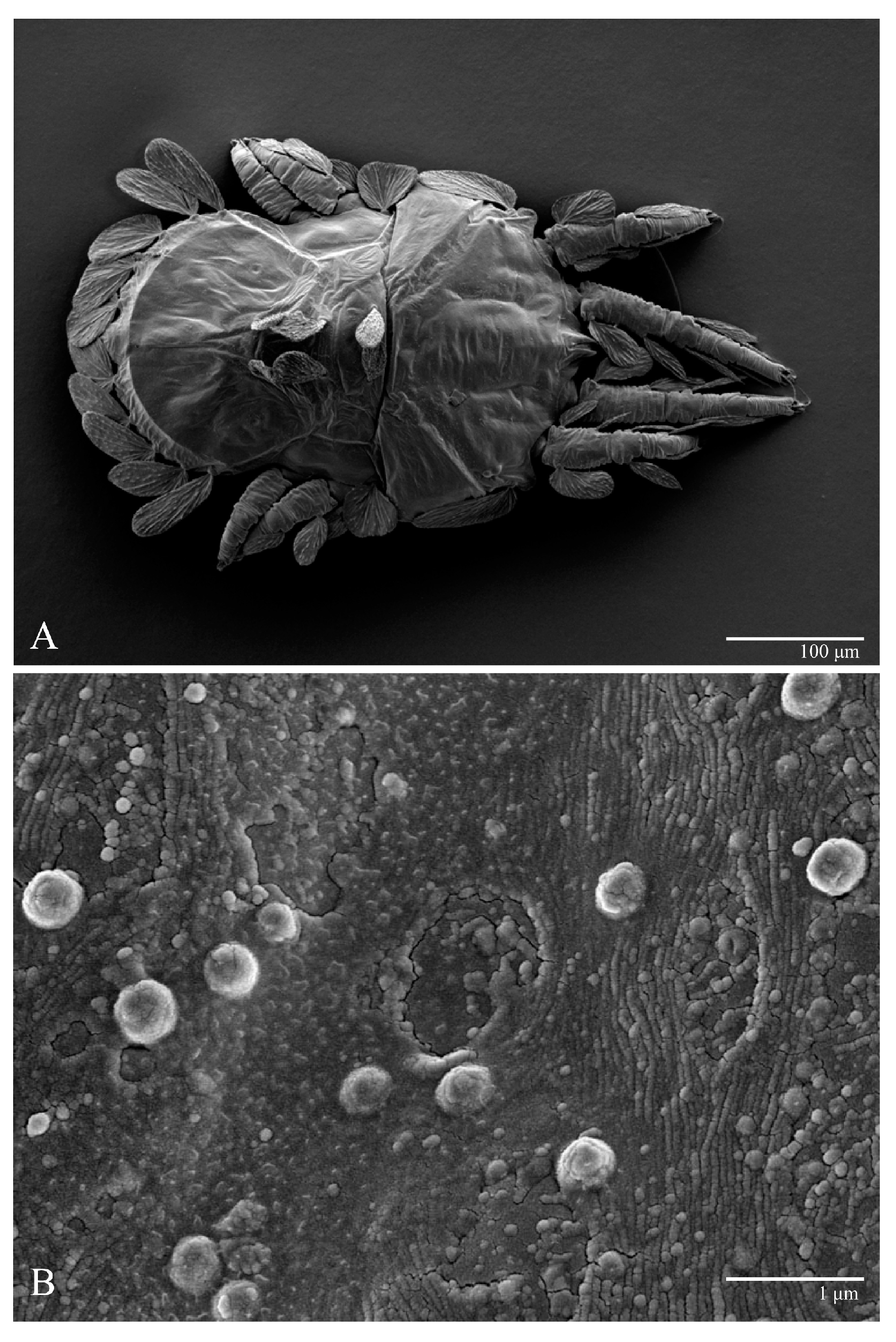
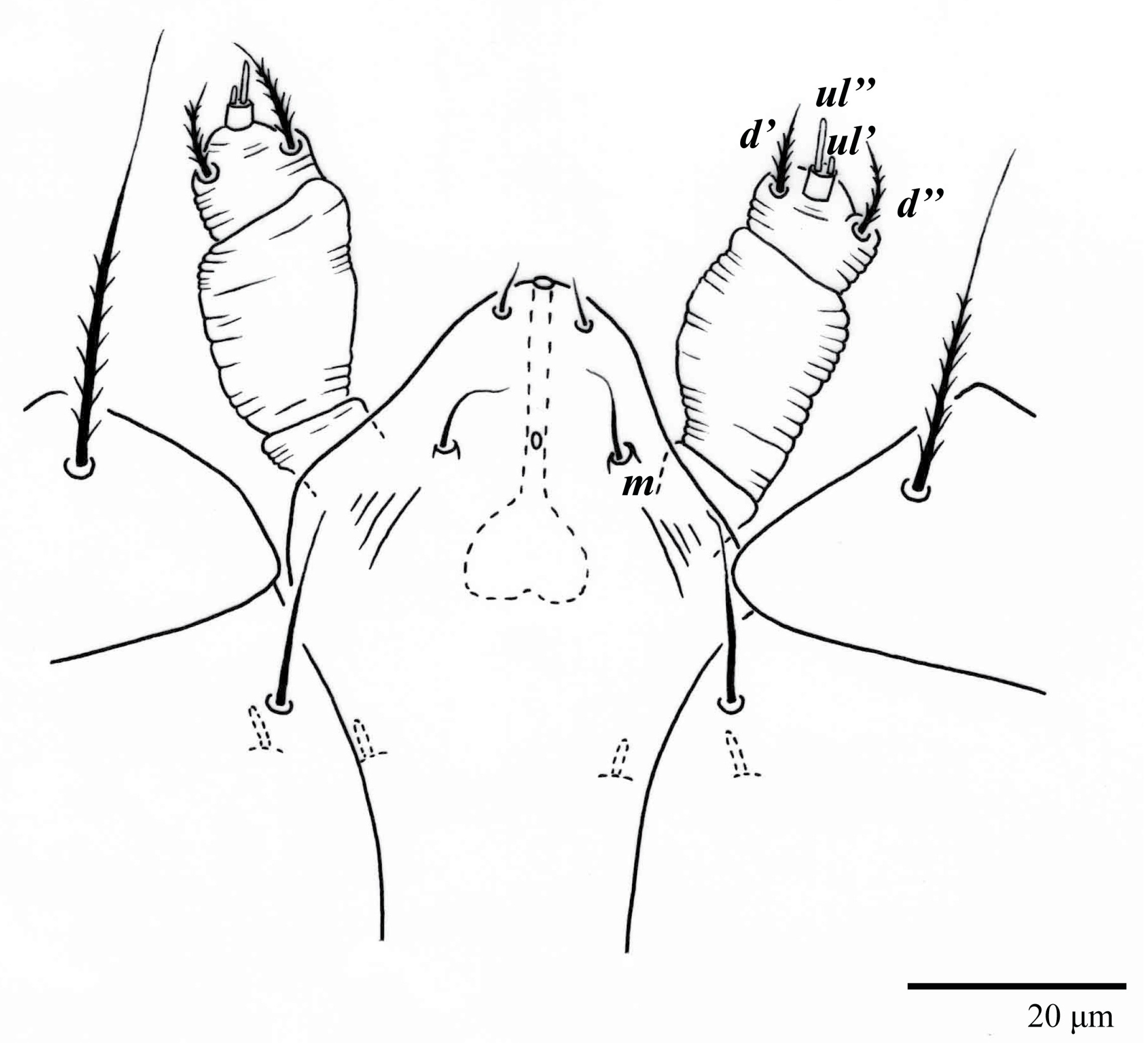
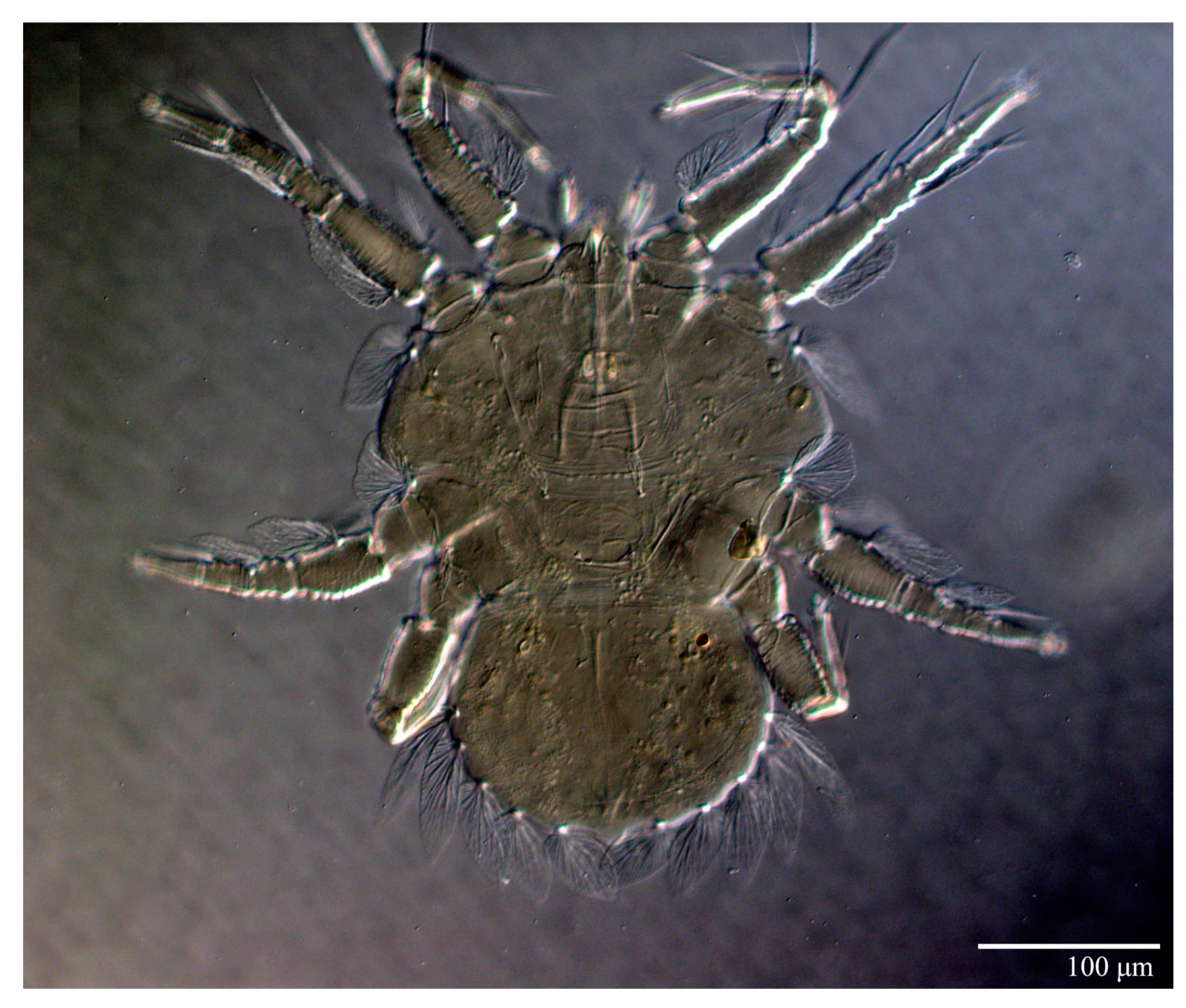
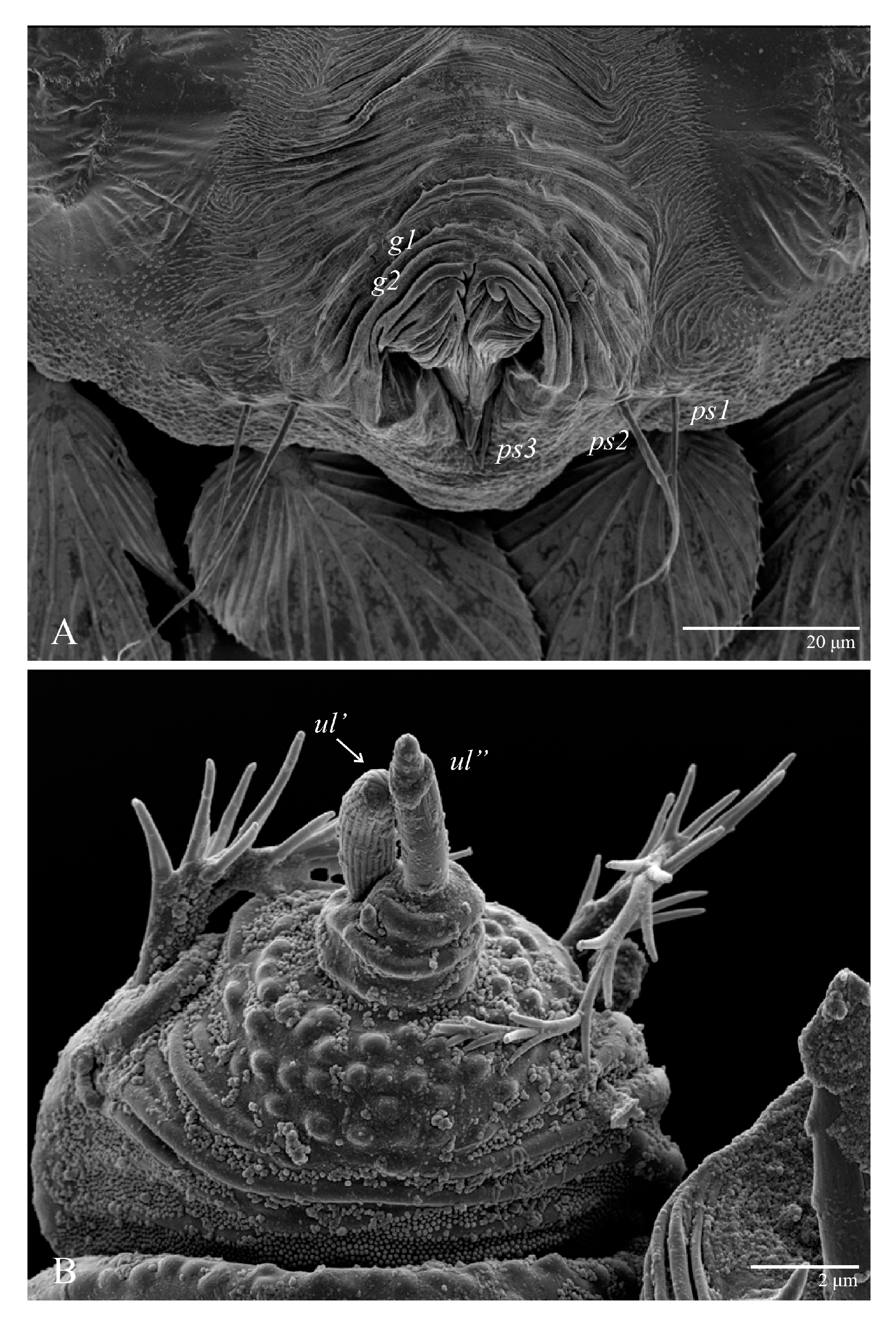
Gnathosoma (
Figure 7,
Figure 8 and
Figure 9). Palps four segmented, setal formula: 0, 0, 2, 2; tibia with two setae,
d′ 7 (6–8),
d″ 6 (5–6), tarsus with one eupathidium 5 (3–5) and one solenidion 1 (1–2). Ventral setae
m 8 (6–8); distance between setae
m–m 14 (13–16). Tips of cheliceral stylets with a few bluntly rounded lateral projections (
Figure 9).
Spermatheca (
Figure 5B). Duct length ca. 75–85, terminating in smooth rounded bulb.
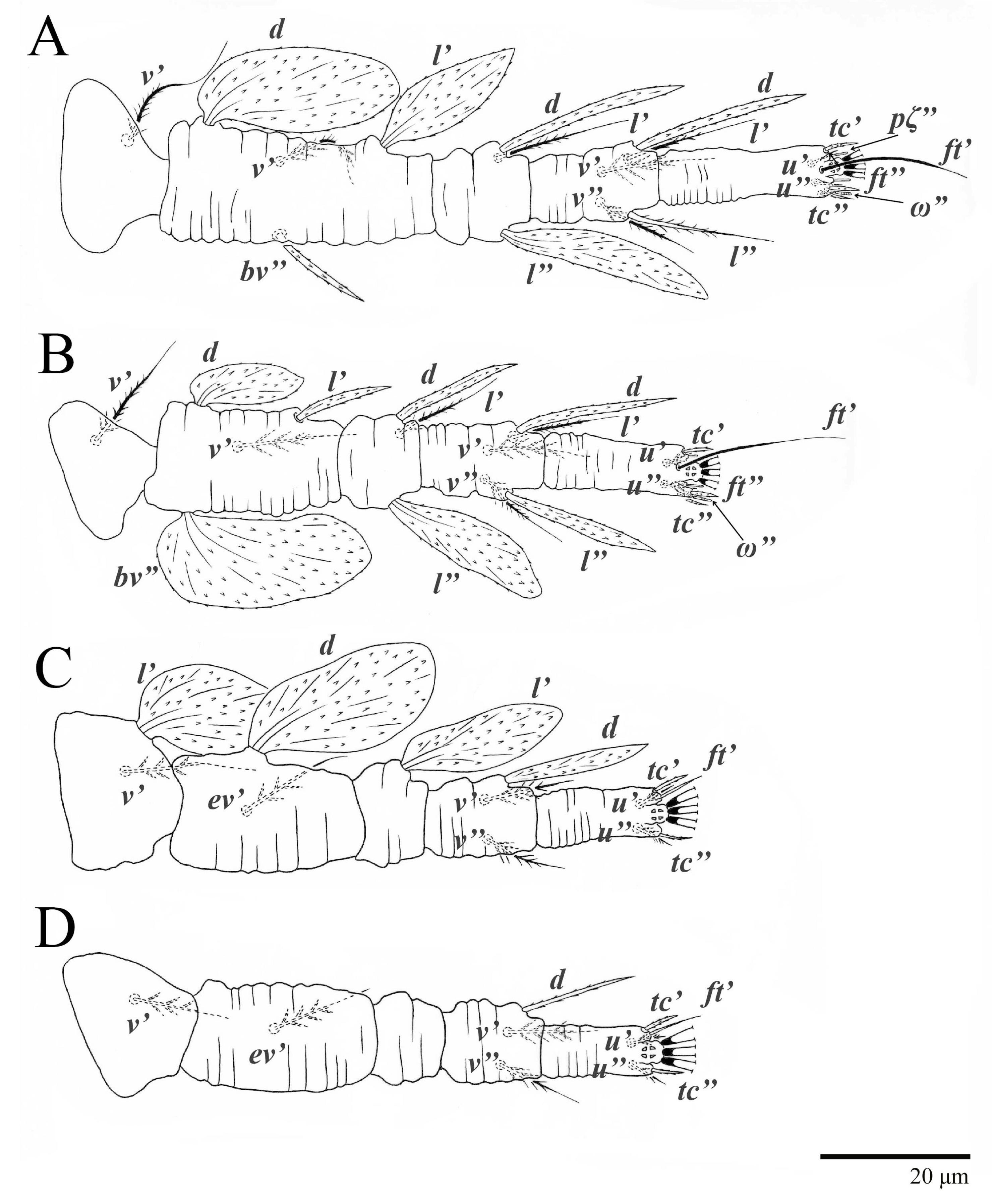
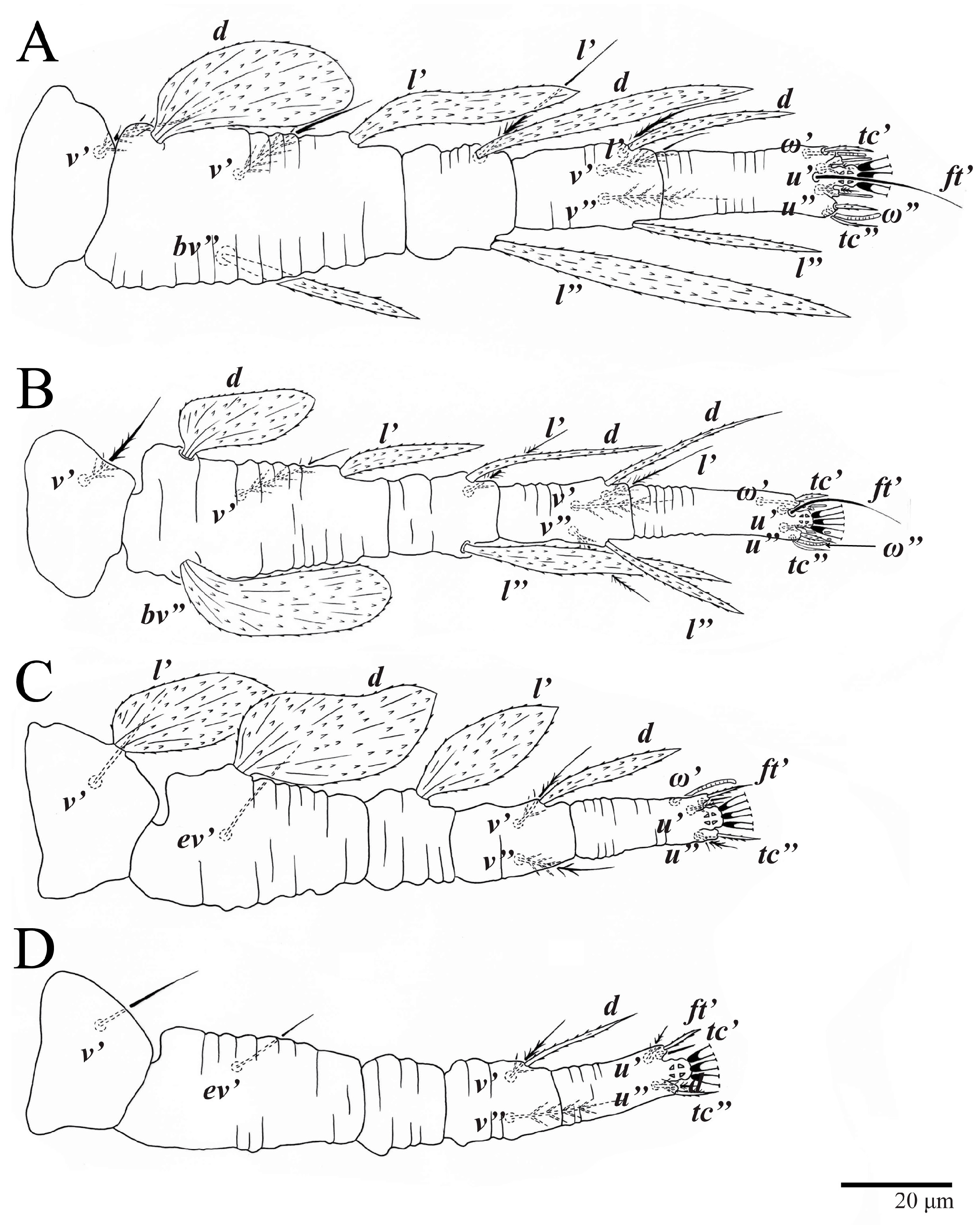
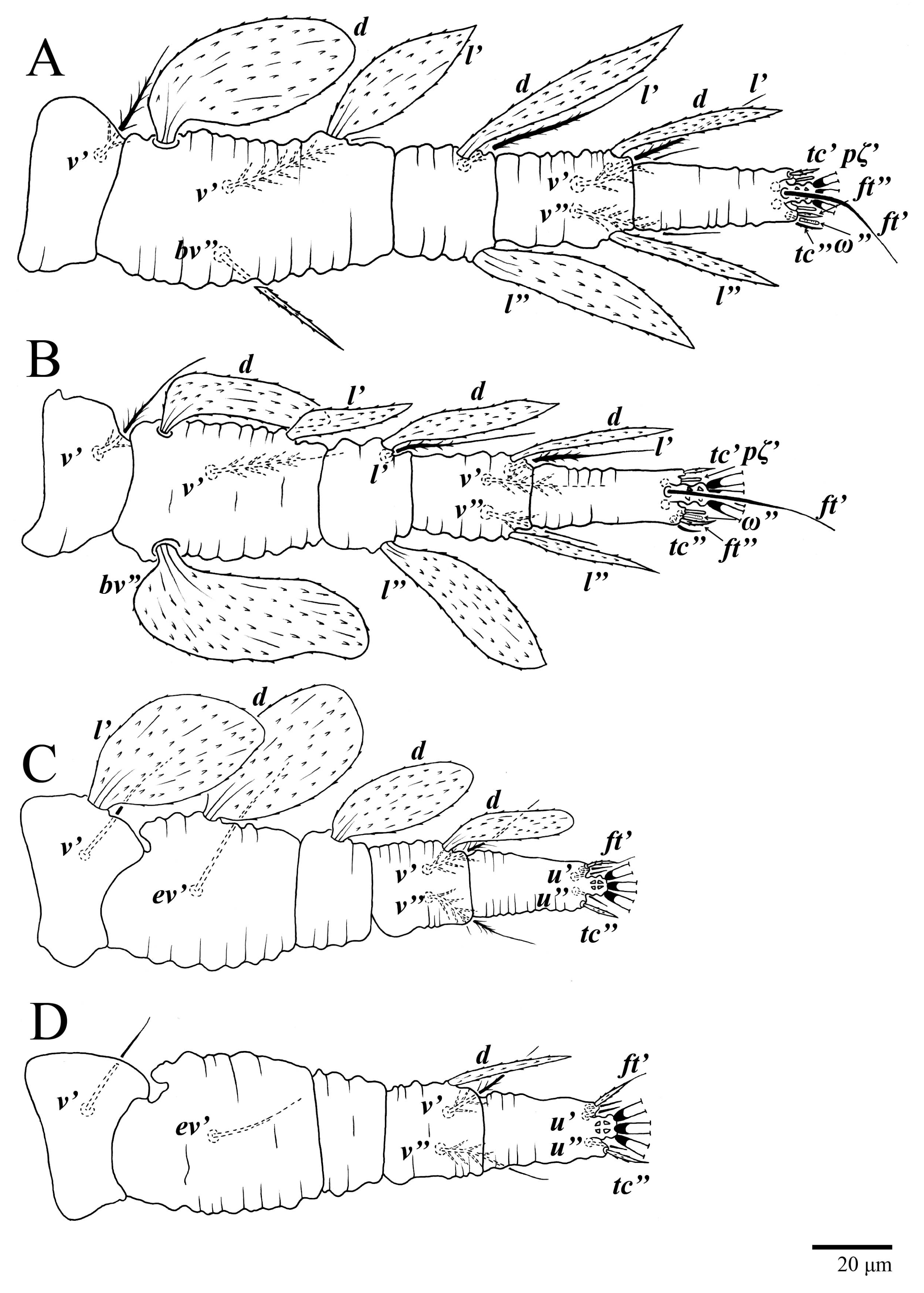
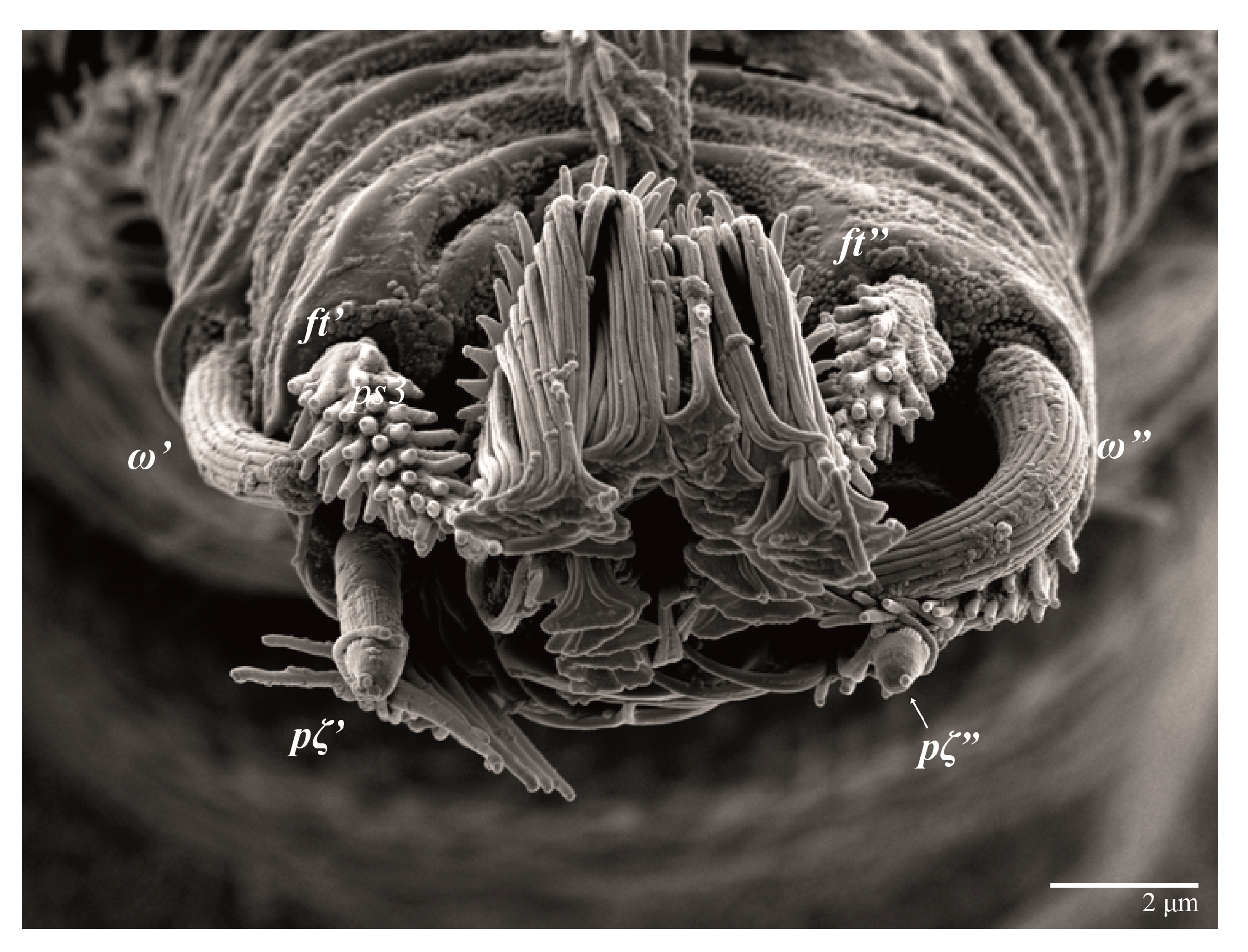
Legs (
Figure 10). Setation (from coxae to tarsi): I 3–1–4–3–5–8(1), II 2–1–4–3–5–8(1), III 1–2–2–1–3–5, IV 1–1–1–0–3–5. Tarsi I–II each with one solenidion
ω” 7 (6–8) (for both tarsi I and tarsi II) and two eupathidia
pζ′–pζ” (5–6, 5–6; 5, 5–6, respectively); femur I with setae
d obovate and
l′ broadly lanceolate; femur II with setae
d narrowly obovate,
l′ lanceolate, and
bv” obovate to broadly falcate. Femora, genua, and tibiae with setae
d inserted in lateral position. Detail of the development of leg chaetotaxy in
Table 1.
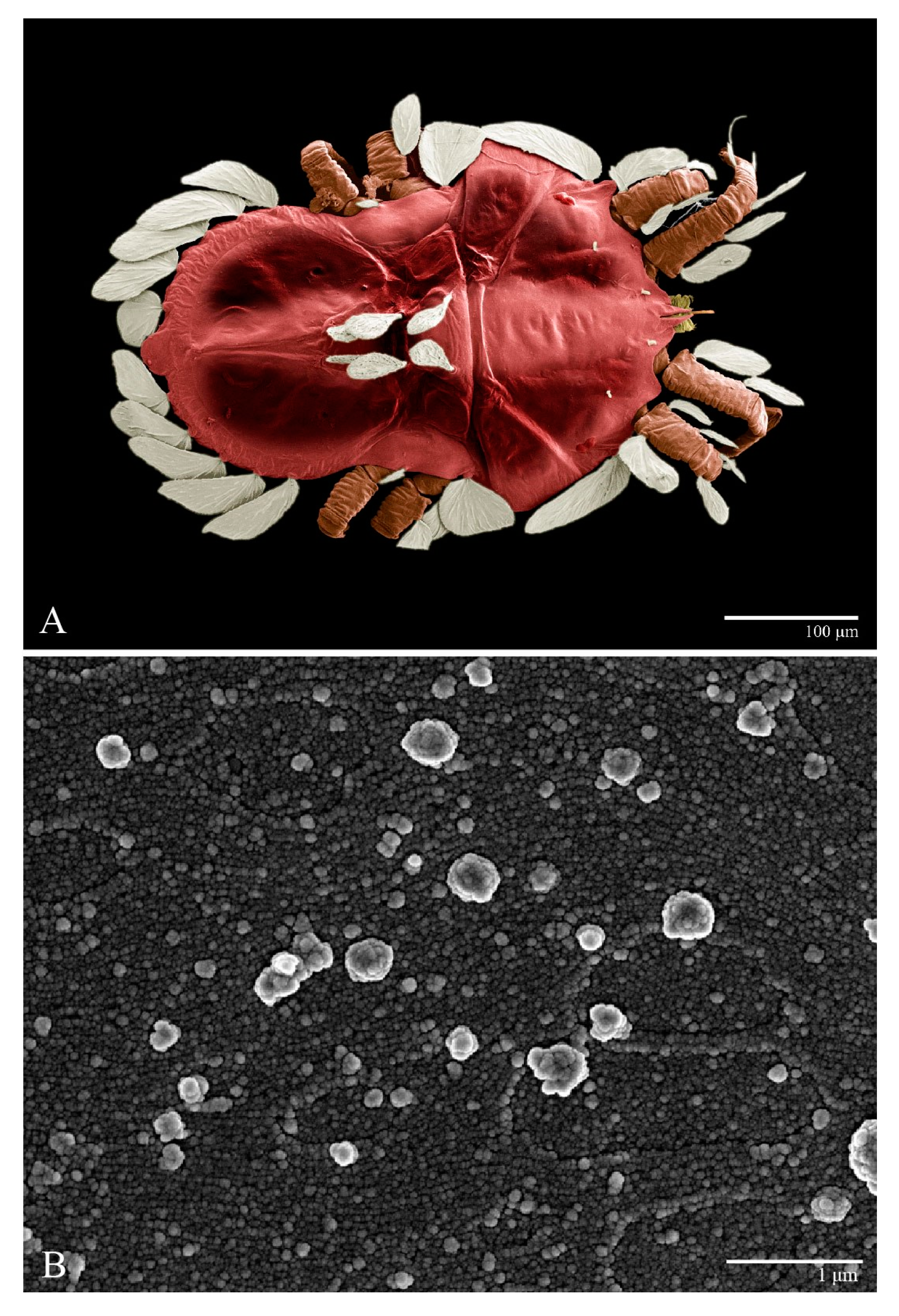
Microplates (
Figure 2B). The microplate layer forms a reticulate network of thick ridges covered in small, single, irregularly-shaped wax-like crystals or masses.
Color. The body is mostly orange with the margin of prodorsum and opisthosoma with dark spots, eyes red, and legs orange. The dorsal body setae and leg setae white to translucent.
Body measurements: distance between setae, v2–h1 285–310, sc2–sc2 200–220; other measurements: v2–v2 45–50, sc1–sc1 105–120, c1–c1 50–58, c3–c3 190–215, d1–d1 27–30, d3–d3 155–175, e1–e1 27–33, e3–e3 165–175, f2–f2 160–170, f3–f3 140–145, h1–h1 55–58, h2–h2 105–115.
Dorsum (
Figure 11 and
Figure 12). Anterior margin of prodorsum with a short median forked projection forming a short notch. The dorsum is smooth, with a pair of lateral projections anterior to setae
sc2 and a single projection between opisthosomal setae
h1. Many dorsal setae similar in general form to those of female, except
c1,
d1, and
e1 short to minute,
d3 longer, and setae along posterior margin of opisthosoma (especially
e3) narrower and more elongated than those of the female. Setal measurements:
v2 5–7,
sc1 4–6,
sc2 60–67,
c1 29–30,
c3 40–45,
d1 8–10,
d3 19–27,
e1 5–7,
e3 77–80,
f2 60–72,
f3 60–63,
h1 49–50,
h2 53–55.
Venter (
Figure 13 and
Figure 14A). Ventral integument weakly striate along central region and densely colliculated along lateral body margin; ventral setae filiform, with coxal setae
1c,
2c, and
3b barbed; setae
ps2 distinctly longer than
ps1; setae
ps3 thickened and inserted ventrally on the elongate tapered anal valves. Setal measurements:
1a 100–105,
1b 18–21,
1c 27–30,
2b 23–29,
2c 34–35,
3a 17–23,
3b 40–42,
4a 120–130,
4b 23–30,
ag 14–15,
g1 12–13,
g2 14–17,
ps1 21–23,
ps2 42–55,
ps3 13–14.
Gnathosoma (
Figure 14B). Palps four segmented, setal formula: 0, 0, 2, 2; tibia with two setae,
d′ 7–8,
d″ 7–8, tarsus with one eupathidium 5–6 and one solenidion 6. Ventral setae
m 6–7; distance between setae
m–m 13–14.
Legs (
Figure 15). Setation (from coxae to tarsi): I 3–1–4–3–5–8(2), II 2–1–4–3–5–8(2), III 1–2–2–1–3–5(1), IV 1–1–1–0–3–5. Tarsi I–II each with two solenidia (one abaxial, one adaxial), tarsi I
ω″ 10–11,
ω′ 16–17, tarsi II
ω″ 11–12
, ω′ 14–15 and two eupathidia
pζ′–
pζ” (6–7, 7; 5–6, 5–6), and tarsus III with one solenidion (paraxial and ventrolateral)
ω′ 13–15. Leg setae similar to that of the female; seta
l” on genu I distinctly elongated. Detail of the development of leg chaetotaxy in
Table 1.
Aedeagus (
Figure 13B). As figured; ca. 130 long.
Body measurements: distance between setae v2–h1 335–365, sc2–sc2 165–180; other measurements: v2–v2 37–40, sc1–sc1 93–105, c1–c1 42–55, c3–c3 235–265, d1–d1 40–50, d3–d3 200–215, e1–e1 18–28, e3–e3 155–175, f2–f2 145–160, f3–f3 120–135, h1–h1 45–50, h2–h2 87–95.
Dorsum (
Figure 16). Anterior margin of prodorsum with a short median forked projection forming a short notch; pair of lateral projections anterior and adjacent to setae
sc2 present; projection not formed (or rudimentary) between setae
h1. Prodorsal region smooth; region between setae
sc2–
c3 with transverse plicae and folds; region posterior to setae
d1–
d3 smooth. Dorsal setae similar in general form to that of females, except setae
c1,
d1 and
e1 short to minute. Setal measurements:
v2 3–4,
sc1 2–3,
sc2 60–64,
c1 3–5,
c3 32–35,
d1 2–3,
d3 3–4,
e1 3–4,
e3 45–54,
f2 40–42,
f3 41–42,
h1 35–36,
h2 38–45.
Gnathosoma. Palps similar to those of female, setal formula: 0, 0, 2, 2; tibia with two setae, d′ 4–5, d″ 4–5, tarsus with one eupathidium 3–4 and one minute solenidion, 1 long. Ventral setae m 4–5; distance between setae m–m 10–11.
Venter. Cuticle covered with fine and mostly transverse striae. Coxal, genital, and anal setae fine. Setal lengths: 1a 80–100, 1b 10–15, 1c 10–12, 2b 10–14, 2c 18–20, 3a 12–13, 3b 12–17, 4a 50–80, 4b 12–21, ag 7–8, g1 8–9, ps1 12–14, ps2 25–27, ps3 9–10. Setae g2 absent.
Legs (
Figure 16). Setation (from coxae to tarsi): I 3–1–4–3–5–8(1), II 2–1–4–3–5–8(1), III 1–2–2–1–3–5, IV 1–0–1–0–3–5. Leg chaetotaxy similar to that of the female, except by trochanter IV nude; tarsi I–II each with one solenidion
ω” (tarsi I 4–5 and tarsi II 5), and two eupathidia
pζ′–
pζ” (4–5, 5; 4–5, 4–5, respectively). Detail of the development of leg chaetotaxy in
Table 1.
Body measurements: distance between setae v2–h1 275–290, sc2–sc2 135–145; other measurements: v2–v2 32–35, sc1–sc1 80–83, c1–c1 40–43, c3–c3 185–195, d1–d1 35–38, d3–d3 150–155, e1–e1 22–25, e3–e3 120–130, f2–f2 110–115, f3–f3 90–95, h1–h1 30–33, h2–h2 62–65.
Dorsum (
Figure 17). Anterior margin of prodorsum with a short median forked projection forming a short notch; pair of lateral body projections anterior and adjacent to setae
sc2 present. Prodorsal region smooth; region between setae
sc2–
c3 with transverse striations and region posterior to setae
c3 smooth; dorsal setae similar to that of the female, except setae
c1,
d1, and
e1 short. Setal measurements:
v2 2–3,
sc1 2–3,
sc2 40–44,
c1 3–4,
c3 24–25,
d1 3–4,
d3 3–4,
e1 2–3,
e3 30–32,
f2 27–28,
f3 24–26,
h1 22–25,
h2 24–25.
Gnathosoma. Palps similar to those of the female, setal formula: 0, 0, 2, 2; tibia with two setae, d′ 4–5, d″ 3–4, tarsus with one eupathidium 3–4 and one solenidion, 1 long. Ventral setae m 4–5; distance between setae m–m 10–12.
Venter. Cuticle covered with fine and mostly transverse striae. Coxal, genital and anal setae fine. Setal measurements: 1a 65–67, 1b 10–11, 1c 9–12, 2c 12–13, 3a 10–13, 3b 14–17, ag 6–7, ps1 7–9, ps2 13–15, ps3 7–8. Setae 2b, 4a, 4b, g1 and g2 absent.
Legs (
Figure 17). Setation (from coxae to tarsi): I 3–0–3–1–5–6(1), II 1–0–3–1–5–6(1), III 1–0–2–0–3–5, IV 0–0–1–0–3–3. Tarsi I–II each with one solenidion
ω” 4–5 (for both tarsi I and tarsi II) and two eupathidia
pζ′–pζ” (all 3–4). Detail of the development of leg chaetotaxy in
Table 1.
Body measurements: distance between setae v2–h1 220–230, sc2–sc2 115–125; other measurements: v2–v2 22–25, sc1–sc1 70–73, c1–c1 32–38, c3–c3 140–150, d1–d1 30–38, d3–d3 110–115, e1–e1 16–18, e3–e3 100–105, f2–f2 86–88, f3–f3 67–70, h1–h1 20–23, h2–h2 40–45.
Dorsum (
Figure 18). Prodorsal region with colliculated integument anteromedially; region between setae
sc2–
c3 with oblique and transverse folds; pygidial region posterior to setae
e1 with colliculated integument; dorsal setae similar in general form to those of females except much smaller and setae
c1,
d1, and
e1 minute. Setal measurements:
v2 2–3,
sc1 1–2,
sc2 25–26,
c1 2–4,
c3 16–18,
d1 2–3,
d3 2–3,
e1 2–3,
e3 22–23,
f2 16–20,
f3 15–18,
h1 15–17,
h2 16–17.
Gnathosoma. Palps similar to those of female, setal formula: 0, 0, 2, 2; tibia with two setae, d′ 3–4, d″ 5, tarsus with one eupathidium 3–4 and one minute solenidion, 1 long. Setae m absent.
Venter. Cuticle covered with fine and mostly transverse striae. Coxal, genital, and anal setae fine. Setal measurements: 1a 55–65, 1b 7–8, 3a 10–11, ps1 5–7, ps2 10–11, ps3 5–6. Setae 1c, 2b, 2c, 3b, 4a, 4b, ag, g1, and g2 absent.
Legs (
Figure 18). Setation (from coxae to tarsi): I 2–0–3–1–5–6(1), II 0–0–3–1–5–6(1), III 0–0–2–1–3–3. Tarsi I–II each with one solenidion
ω” 3–4 (for both tarsi I and II) and two eupathidia
pζ′–pζ” (3–4, 3–4; 3–4, 3–4, respectively). Cuticle of all legs covered with colliculated sculpturing. Detail of the development of leg chaetotaxy in
Table 1.
Etymology. The specific name parameekeri refers to the morphological similarity of this species and U. meekeri (De Leon), the type species of the genus.
Differential diagnosis. This new species resembles Ultratenuipalpus meekeri (De Leon) (herein redescribed) as they both have dorsal setae of a similar shape and length and the same leg and palp chaetotaxy in all developmental stages. These two species also share several other characteristics, such as the pair of lateral projections anterior to setae sc2 and a single posterior projection between opisthosomal setae h1. However, the two species can be separated: the prodorsum is distinctly broader in adult females and males (measured at the widest point between setae sc1 and c1) in U. meekeri (325–345) than U. parameekeri (290–315) (in females); notch in anterior forked projection is shorter in U. meekeri (8–13) than in U. parameekeri (20–27) (in females); e3 is narrower and more lanceolate on male (and to a lesser extent on females) U. meekeri than in U. parameekeri; l” on ti I on U. meekeri is thicker than on U. parameekeri; d on fe II is longer and more falcate on female U. meekeri than on U. parameekeri; c3 in larvae is narrower and more lanceolate in U. meekeri than U. parameekeri. In addition to the morphological differences, the molecular analyses confirmed that U. parameekeri and U. meekeri represent distinct species, with a 15.7% difference between their COI sequences.
DNA Barcoding. DNA was successfully amplified and the mitochondrial cytochrome C oxidase subunit I gene (COI) sequenced from one specimen of
U. parameekeri collected on
Cyclosorus interruptus (Thelypteridaceae) from Pindorama, São Paulo, Brazil; sequence data have been deposited in GenBank (
https://www.ncbi.nlm.nih.gov/, accessed on 15 January 2023), with the following accession code: female, 398 base pairs (GenBank: OQ533138).
Type material examined. Holotype: female collected on ferns Rumohra adiantiforme (Dryopteridaceae) from Ilha do Cardoso, São Paulo, Brazil, 22 March 2017, coll. G.C.O. Piccoli (DZSJRP). Paratypes: 3 females, 1 protonymph, and 2 larvae, with the same data as the holotype (DZSJRP); 4 females, 3 males, 4 deutonymphs, 5 protonymphs, and 2 larvae collected on Psychotria nuda (Rubiaceae) from Ilha do Cardoso, São Paulo, Brazil, 22 March 2017, coll. G.C.O. Piccoli (DZSJRP); 2 females and 2 males collected on P. nuda from Ilha do Cardoso, São Paulo, Brazil, 22 March 2017, coll. G.C.O. Piccoli (NMNH); 4 females and 1 deutonymph collected on ferns C. interruptus from Pindorama, São Paulo, Brazil, 15 December 2002, coll. R. Kishimoto (DZSJRP).
Other material examined. 1 female and 1 larva collected on ferns C. interruptus from Pindorama, São Paulo, Brazil, 15 March 2003, coll. P. Demite (DZSJRP); 2 females, 2 deutonymphs, 3 protonymphs, and 1 larva collected on ferns C. interruptus from Pindorama, São Paulo, Brazil, 15 December 2002, coll. R. Kishimoto (DZSJRP); 2 females, 2 deutonymphs, 3 protonymphs, and 1 larva collected on ferns C. interruptus from Pindorama, São Paulo, Brazil, 15 March 2005, coll. P. Demite (DZSJRP, USNM).
3.2. Redescription of Ultratenuipalpus meekeri (De Leon, 1957)
Tenuipalpus meekeri De Leon: De Leon [
13]—original designation
Ultratenuipalpus meekeri (De Leon): Mitrofanov [
14]
Diagnosis. Female: As per genus, in addition to: prodorsal setae v2, sc1 minute to short, and sc2 large, flattened, obovate to ovate; dorsal opisthosoma with 10 pairs of setae (f2 present); most of the dorsal opisthosomal setae large, flattened, obovate to ovate, except setae d3 is distinctly short and c3 is almost orbicular; pair lateral projections anterior to setae sc2 and single posterior projection between opisthosomal setae h1 present; palp four segmented, setal formula 0, 0, 2, 2. Male: Opisthosoma narrower than that of females, with a distinct transverse constriction (waist) between setae d1 and e1; many dorsal setae similar to those of the female, except c1 much smaller, d1 and e1 short to minute, and v2 and d3 longer. Tarsi I–II each with two solenidia (ω′ paraxial and ventrolateral; ω′ antiaxial); tarsus III with one solenidion ω′ paraxial and ventrolateral. Immatures: with lateral body projections anterior to setae sc2 present (except absent in larvae); single posterior projection between setae h1 absent; dorsal setae similar in general form to those of the female, except c1, d1, and e1 minute. Larvae with anterior margin colliculated and central prodorsum smooth; pygidial region of posterior opisthosoma with colliculated integument.
Body measurements: distance between setae v2–h1 390 (375–390), sc2–sc2 230 (230–235); other measurements: v2–v2 42 (42–48), sc1–sc1 115 (115), c1–c1 65 (65–68), c3–c3 290 (260–290), d1–d1 43 (37–43), d3–d3 250 (235–250), e1–e1 28 (25–30), e3–e3 235 (230–240), f2–f2 225 (215–225), f3–f3 190 (185–195), h1–h1 72 (72–78), h2–h2 135 (130–140).
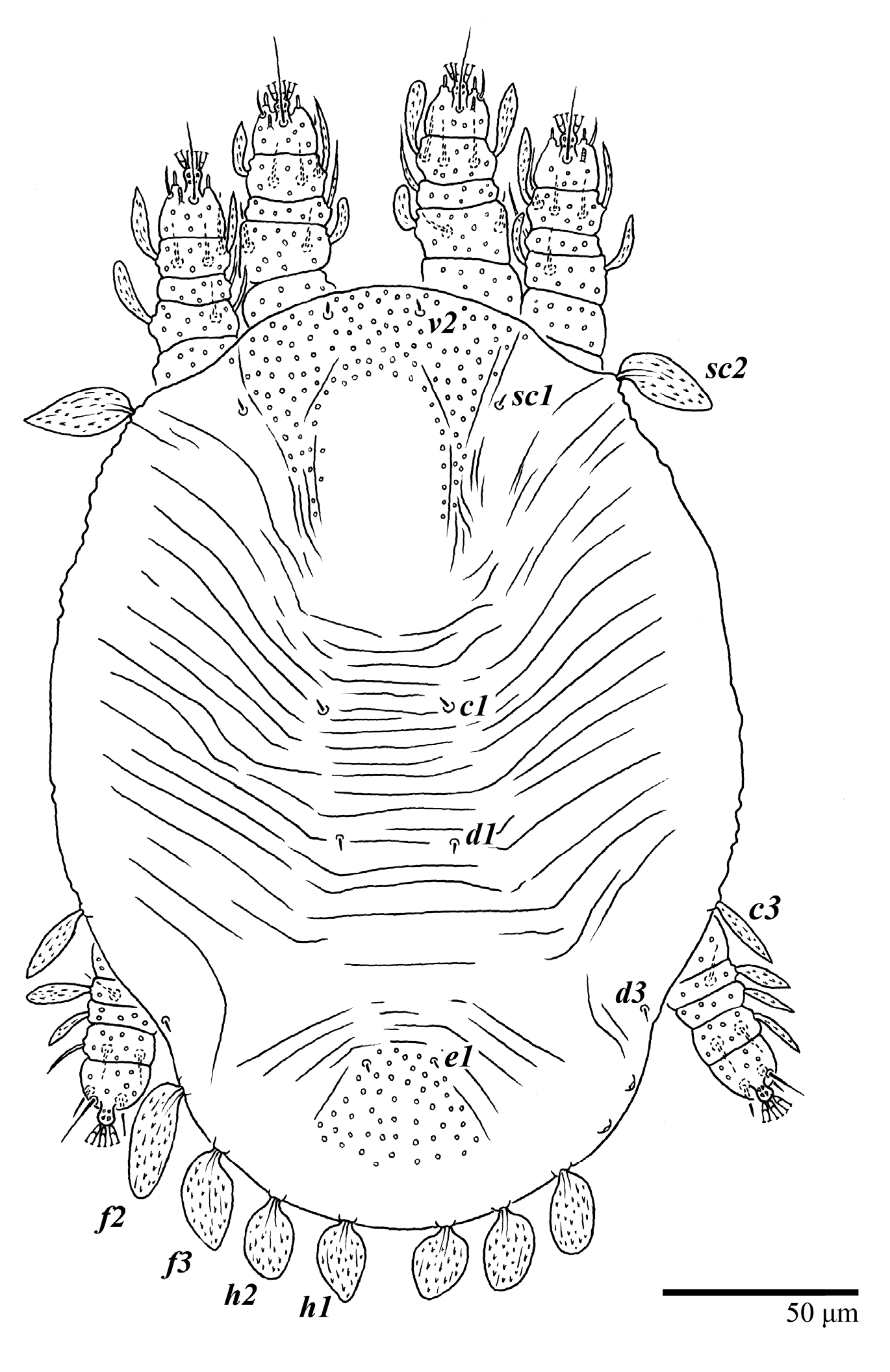
Dorsum (
Figure 19,
Figure 20 and
Figure 21). Anterior margin of prodorsum with a short median forked projection forming a short notch 8 (8–13). Dorsum smooth, with pair of lateral projections anterior to setae
sc2 and a single projection between opisthosomal setae
h1 present. A pair of converging folds from the eyes to near the sejugal furrow on the prodorsum posterior margin. Prodorsal setae
v2 and
sc1 short to minute;
sc2 large, flattened elongated obovate (
Figure 19 and
Figure 20A); most opisthosomal setae similar to prodorsal setae
sc2, except
d3 short. Setal measurements:
v2 8 (4–8),
sc1 4 (4–10),
sc2 83 (83–94),
c1 65 (65–69),
c3 57 (54–57),
d1 55 (55–58),
d3 14 (14–15),
e1 48 (39–48),
e3 95 (92–95),
f2 80 (80–84),
f3 77 (77–83),
h1 63 (62–66),
h2 70 (70–73).
Venter (
Figure 22A). Ventral integument weakly striate along central region and densely colliculated around lateral body margin; ventral, genital, and anal plates not developed, entire region membranous and distinctly plicate; ventral setae filiform, with coxal setae
1c,
2c, and
3b barbed; setae
ps2 distinctly longer than
ps1. Setal measurements:
1a 105 (105–115),
1b 19 (12–19),
1c 29 (26–29),
2b 27 (27–28),
2c 47 (41–47),
3a 20 (20–21),
3b 43 (37–43),
4a 105 (95–115),
4b 26 (23–26),
ag 15 (15–17),
g1 19 (15–19),
g2 17 (17–20),
ps1 15 (12–15),
ps2 48 (48–60),
ps3 31 (23–31).
Gnathosoma (
Figure 23). Palps four segmented, setal formula: 0, 0, 2, 2; tibia with two setae,
d′ 7 (7–11),
d″ 8 (7–8), tarsus with one eupathidium 5 (5) and one solenidion, 1 (1) long. Ventral setae
m 7 (7–8); distance between setae
m–
m 15 (13–15).
Spermatheca (
Figure 22B). Duct length ca. 70–85, terminating in smooth rounded bulb.
Legs (
Figure 24). Setation (from coxae to tarsi): I 3–1–4–3–5–8(1), II 2–1–4–3–5–8(1), III 1–2–2–1–3–5, IV 1–1–1–0–3–5. Tarsi I–II each with one solenidion
ω” 9 (8–9) (for both tarsi I and tarsi II) and two eupathidia
pζ′–
pζ” (7, 7; 7, 6–7, respectively); femur I with setae
d obovate and
l′ broadly lanceolate; femur II with setae
d elongate obovate to weakly falcate,
l′ lanceolate and
bv” obovate to broadly falcate. Femora, genua, and tibiae with setae
d inserted in lateral position. Detail of the development of leg chaetotaxy in
Table 1.
Color (
Figure 20A). The body is reddish with the central region becoming darker, eyes red, and legs orange. Dorsal body setae and legs setae are white.
Body measurements: distance between setae v2–h1 280, sc2–sc2 210; other measurements: v2–v2 43, sc1–sc1 110, c1–c1 65, c3–c3 205, d1–d1 30, d3–d3 165, e1–e1 28, e3–e3 175, f2–f2 170, f3–f3 150, h1–h1 63, h2–h2 110.
Dorsum (
Figure 25 and
Figure 26). Anterior margin of prodorsum with a short median forked projection forming a short notch. Dorsum smooth, with pair lateral projections anterior to setae
sc2 and a single projection between opisthosomal setae
h1 present. Prodorsum with a pair of converging folds from the eyes to near the sejugal furrow on the posterior margin. Dorsal setae similar in general form to those of the female, except
c1,
d1, and
e1 small to minute, and
d3 longer. Setal measurements:
v2 10,
sc1 8,
sc2 65,
c1 23,
c3 49,
d1 6,
d3 18,
e1 5,
e3 74,
f2 70,
f3 66,
h1 53,
h2 57.
Venter (
Figure 27 and
Figure 28A). Ventral integument weakly striated along central region and densely colliculated around lateral margin of body; ventral setae filiform; coxal setae
1c,
2c, and
3b barbed; setae
ps2 distinctly longer than
ps1; setae
ps3 thickened and inserted ventrodistally on elongated, tapered anal valves. Setal measurements:
1a 85,
1b 22,
1c 28,
2b 26,
2c 35,
3a 21,
3b 35,
4a 90,
4b 23,
ag 20,
g1 19,
g2 16,
ps1 24,
ps2 60,
ps3 16.
Gnathosoma (
Figure 28B). Palps four segmented, setal formula: 0, 0, 2, 2; tibia with two setae,
d′ 8,
d″ 7, tarsus with one eupathidium 5 and one solenidion 6. Ventral setae
m 8; distance between setae
m–
m 14.
Legs. Setation (from coxae to tarsi): I 3–1–4–3–5–8(2), II 2–1–4–3–5–8(2), III 1–2–2–1–3–5(1), IV 1–1–1–0–3–5. Tarsi I–II (
Figure 29) each with two solenidia (one abaxial, one adaxial), tarsi I
ω′ 12,
ω″ 9, tarsi II
ω′ 13,
ω″ 9, and two eupathidia
pζ′–
pζ” (all 6–7), and tarsus III with one solenidion (paraxial and ventrolateral)
ω′ 12. Leg setae similar to that of the female. Detail of the development of leg chaetotaxy in
Table 1.
Body size measurements: distance between setae v2–h1 310–350, sc2–sc2 170–190; other measurements: v2–v2 37–40, sc1–sc1 95–105, c1–c1 43–53, c3–c3 220–270, d1–d1 45–55, d3–d3 200–225, e1–e1 25–30, e3–e3 160–180, f2–f2 145–170, f3–f3 120–145, h1–h1 40–55, h2–h2 83–105.
Dorsum (
Figure 30). Anterior margin of prodorsum with a short median forked projection forming a short notch; a pair of body projections anterior and adjacent to setae
sc2 present; posterior projection between setae
h1 absent. Prodorsum with central region smooth; region between setae
sc2–
c3 with transverse folds and plicae; region posterior to setae
e1 smooth. Dorsal setae similar in general form to those of the female, except setae
c1,
d1, and
e1 are short to minute. Setal measurements:
v2 3–5,
sc1 4–5,
sc2 64–78,
c1 6–12,
c3 36–41,
d1 5–7,
d3 5–7,
e1 4–8,
e3 52–64,
f2 51–55,
f3 47–55,
h1 36–42,
h2 43–51.
Gnathosoma. Palps similar to those of female, setal formula: 0, 0, 2, 2; tibia with two setae, d′ 6–7, d″ 5–6, tarsus with one eupathidium 4–5 and one minute solenidion 1. Ventral setae m 5–7; distance between setae m–m 12–13.
Venter (
Figure 31). Cuticle covered with fine and mostly transverse striae; with band of a colliculated cuticle around posterior body margin. Coxal, genital, and anal setae fine. Setal lengths:
1a 75–90,
1b 9–12,
1c 11–15,
2b 10–18,
2c 13–16,
3a 10–15,
3b 16–17,
4a 60–80,
4b 11–12,
ag 11–15,
g1 8–11,
ps1 9–10,
ps2 27–33,
ps3 15–17. Setae
g2 absent.
Legs (
Figure 30). Setation (from coxae to tarsi): I 3–1–4–3–5–8(1), II 2–1–4–3–5–8(1), III 1–2–2–1–3–5, IV 1–0–1–0–3–5. Leg chaetotaxy similar to that of the female, except by trochanter IV nude; tarsi I–II each with one solenidion
ω” (tarsi I 5–6 and tarsi II 5, 6) and two eupathidia
pζ′–
pζ” (5–6, 5–6; 5, 5 respectively). Detail of the development of leg chaetotaxy in
Table 1.
Body size measurements: distance between setae v2–h1 230, sc2–sc2 150; other measurements: v2–v2 28, sc1–sc1 85, c1–c1 35, c3–c3 190, d1–d1 25, d3–d3 155, e1–e1 23, e3–e3 130, f2–f2 120, f3–f3 100, h1–h1 38, h2–h2 73.
Dorsum (
Figure 32). Anterior margin of prodorsum with a short median forked projection forming a short notch; a pair of lateral body projections anterior and adjacent to setae
sc2 present; posterior projection between setae
h1 absent. Prodorsum with central region smooth; region between setae
sc2–
c3 with transverse folds and plicae; region posterior to setae
e1 smooth; dorsal setae similar in general form to those of the female, except setae
c1,
d1, and
e1 short to minute. Setal measurements:
v2 3,
sc1 2,
sc2 54,
c1 4,
c3 27,
d1 4,
d3 4,
e1 4,
e3 45,
f2 38,
f3 35,
h1 30,
h2 35.
Gnathosoma. Palps similar to those of the female, setal formula: 0, 0, 2, 2; tibia with two setae, d′ 4, d″ 4, tarsus with one eupathidium 3 and one minute solenidion, 1 long. Ventral setae m 5; distance between setae m–m 12.
Venter. Cuticle covered with fine and mostly transverse striae. Coxal, genital, and anal setae fine. Setal measurements: 1a 70, 1b 9, 1c 8, 2c 13, 3a 14, 3b 11, ag 10, ps1 8, ps2 16, ps3 10. Setae 2b, 4a, 4b, g1, and g2 absent.
Legs (
Figure 32). Setation (from coxae to tarsi): I 3–0–3–1–5–6(1), II 1–0–3–1–5–6(1), III 1–0–2–1–3–5, IV 0–0–1–0–3–3. Tarsi I–II each with one solenidion
ω” 4 (for both tarsi I and tarsi II) and two eupathidia
pζ′–
pζ” (all 5). Detail of the development of leg chaetotaxy in
Table 1.
Body size measurements: distance between setae v2–h1 225, sc2–sc2 120; other measurements: v2–v2 33, sc1–sc1 68, c1–c1 33, c3–c3 160, d1–d1 30, d3–d3 120, e1–e1 18, e3–e3 115, f2–f2 105, f3–f3 88, h1–h1 23, h2–h2 58.
Dorsum (
Figure 33). Prodorsal region with broad band of a colliculated integument anteromedially between setae
sc1; region between setae
sc2–
c3 with oblique and transverse folds and plicae; pygidial region posterior to setae
e1 with a small region of colliculated integuments; dorsal setae similar in general form to those of the female, except setae
c1,
d1, and
e1 are short to minute. Setal measurements:
v2 3,
sc1 3,
sc2 28,
c1 4,
c3 23,
d1 2,
d3 3,
e1 3,
e3 missing,
f2 30,
f3 25,
h1 21,
h2 21.
Gnathosoma. Palps similar to those of female, setal formula: 0, 0, 2, 2; tibia with two setae, d′ 3, d″ 5, tarsus with one eupathidium 3 and one minute solenidion, 1 long. Setae m absent.
Venter. Cuticle covered with fine and mostly transverse striae. Coxal, genital, and anal setae fine. Setal measurements: 1a 50, 1b 7, 3a 10, ps1 7, ps2 11, ps3 6. Setae 1c, 2b, 2c, 3b, 4a, 4b, ag, g1, and g2 absent.
Legs (
Figure 33). Setation (from coxae to tarsi): I 2–0–3–1–5–6(1), II 0–0–3–1–5–6(1), III 0–0–2–1–3–3. Tarsi I–II each with one solenidion
ω” 3 (for both tarsi I and II) and two eupathidia
pζ′–
pζ” (5, 5; 4, 4, respectively). Cuticles of all legs covered with colliculated cuticles. Detail of the development of leg chaetotaxy in
Table 1.
Remarks. The new specimens examined in this study have similar body and setal measurements to those of the type specimens. In addition, the palp and leg chaetotaxy of those specimens match those of the type specimens.
DNA Barcoding. DNA was successfully amplified and the mitochondrial cytochrome C oxidase subunit I gene (COI) sequenced from one specimen of
U. meekeri collected on
Acrostichum danaeifolium (Pteridaceae) from Tecpan de Galeana, Guerrero State, Mexico; sequence data have been deposited in GenBank (
https://www.ncbi.nlm.nih.gov/, accessed on 15 January 2023), with the following accession code: female, 446 base pairs (GenBank: OQ533137).
Type material examined: Holotype: the female collected on a fern in a mangrove swamp, from San Blas, Nayarit State, Mexico, 21 March 1957, coll. D. De Leon, was deposited in the Museum of Comparative Zoology (MCZ), Harvard University. Paratypes: 2 females, 1 male, 3 deutonymphs, 1 protonymph, and 1 larva, with the same data as the holotype, were deposited in the National Insect and Mite Collection, National Museum of Natural History (NMNH), Smithsonian Institution.
Other material examined: Non-type material: 5 females collected on ferns A. danaeifolium in a mangrove swamp, from Tecpan de Galeana, Guerrero State, Mexico, 5 September 2017, coll. G. Otero-Colina (USNM, DZJSRP).


 ,
, 

































{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}