1. Introduction
Calf health is critical to the production performance and meat quality of growing cattle, which influences the sustainable development of the beef cattle industry. Early weaning is gradually conducted in rearing calves because it can accelerate the gastrointestinal development of calves and reduce production costs [
1]. Nevertheless, calves experience a significant stress reaction due to the transformation of feed type after weaning. On the other hand, the gastrointestinal tract, particularly the rumen, of weaned calves is in the developmental process and vulnerable to changes in their living environment [
2]. As a result, weaned calves have less resistance to harmful bacteria that can cause the dysfunction of digestive organs [
3]. In addition, the immune system of weaned calves is not fully developed; therefore, calves cannot effectively regulate their immunity to maintain healthy growth. Previously, a study on dairy calves found that weaning could cause an inflammatory reaction and increase concentrations of inflammatory factors, including interleukin-1β (IL-1β), IL-6, and tumor necrosis factor-α (TNF-α) in blood [
4]. These adverse influences caused by weaning can ultimately affect the healthy growth of calves and decrease the productivity of growing cattle. Therefore, mitigating the weaning stress and promoting the healthy growth of weaned beef calves are of great significance for beef cattle farming.
Grape seed extract (Gse) is a kind of polyphenol extracted and isolated from grape seeds, mainly including procyanidin, catechin, gallic acid and epicatechin. Recently, the utilization of Gse has become widespread because of its various antimicrobial, anti-inflammatory and antioxidative abilities [
5]. In dairy cows, the dietary supplementation of 1% grape seed and grape marc meal extract (dry matter basis) had no significant effects on dry matter intake (DMI) but increased the milk production [
6]. In lambs, the addition of grape seed procyanidin in the ration at 20 mg/kg body weight (BW) can increase DMI (1.89 kg/d vs. 1.99 kg/d) and average daily gain (ADG) (280.4 g/d vs. 315.9 g/d) [
7]. Due to being rich in polyphenols, a previous study reported that Gse supplementation could enhance the antioxidant ability and feed efficiency of rabbits under heat stress [
8]. Furthermore, Gse was verified to regulate inflammatory responses by decreasing the gene expressions of pro-inflammatory cytokines in the intestine [
9]. A recent study found that dietary supplementation of Gse could improve energy metabolism and colonic fermentation, thus promoting the healthy growth of pre-weaning dairy calves under heat stress [
10]. Nevertheless, the influences of Gse supplementation on the ruminal fermentation, nutrient digestibility and immunity of weaned calves have not been fully investigated.
In calves, the rumen, which plays an important role in the digestion and absorption of volatile fatty acid (VFA) and ammonia nitrogen (NH
3-N), is immature. The ruminal development is strongly linked to VFA yield and absorption, which is main energy source for ruminants [
11]. An in vitro study demonstrated that Gse could regulate VFA production and affect ruminal fermentation [
12]. As mentioned earlier, weaning can trigger oxidative stress and inflammatory responses and down-regulate the immunity of calves. Previous experiments in cows, calves and lambs reported that the supplementation of grape seed products could reduce inflammatory reactions and improve oxidation resistance, and thus is conducive to promoting production performance [
10,
13,
14]. Although Gse can improve the production performance and antioxidant ability of animals, research on the role of Gse as an effective feed additive in young ruminants is limited. According to previous experiments, we hypothesized that Gse may be a promising feed additive than can promote the healthy growth of weaned cross-breed beef calves. Thus, the present research was carried out to explore the influence of Gse supplementation on growth performance, ruminal fermentation, and nutrient digestibility, as well as the biochemical, antioxidative and immune parameters of serum, in weaned cross-breed beef calves.
4. Discussion
In calves, the digestive and immune functions are in developmental phases, and they are easily affected by external environments or pathogens. After weaning, calves undergo greatly physiological challenges, resulting in stunted growth, decreased immunity and higher morbidity [
24]. Relieving the weaning stress of weaned calves is of great significance for cattle farming. As a natural antioxidant, Gse has various health benefits, including free radical scavenging activity, antimicrobial and anti-inflammatory activities [
5]. A previous study reported that the dietary supplementation of grape seed procyanidin could increase the final BW, DMI and ADG in finishing lambs [
7]. In our study, Gse supplementation increased the ADG of weaned cross-breed beef calves, while it had no significant difference on final BW and DMI. The reason that different results may be found is because the growth stages (young stage vs. fattening stage) of animals are different. On the other hand, the findings of the present study show that feed efficiency was slightly ameliorated by Gse supplementation. In a previous study, the addition of Gse improved the feed efficiency of calves suffering from heat stress [
25], which is in accordance with our study. The polyphenols in Gse can inhibit the growth of pathogenic bacteria (e.g.,
Escherichia coli and
Salmonella) in the digestive tract of animals [
26], which is helpful for decreasing the incidence rate of diarrhea. The better growth performance of ruminants is commonly related to improved digestive ability and immunity. Thus, the following experiments were carried out to explore the effects of Gse supplementation on ruminal fermentation, nutrient digestibility, antioxidant ability and immunity of weaned beef calves.
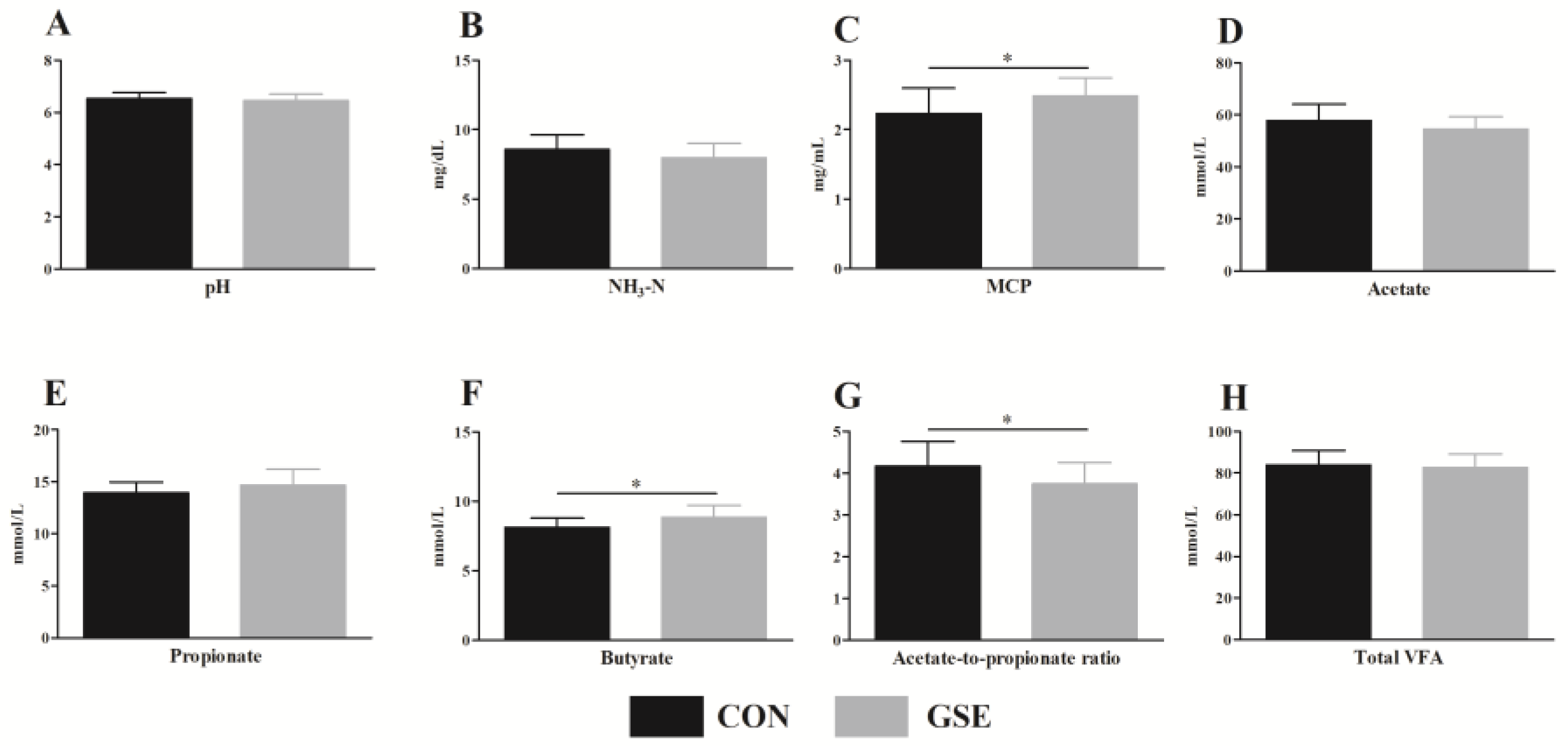
The ruminal pH, which affects the growth and proliferation of microorganisms and regulates VFA production, can be used to assess the healthy status of rumen within a normal range 6 to 7 [
27]. Previously, a study found that supplementing low forage diets with grape marc could decrease ruminal pH [
28]. Nevertheless, in our study, the ruminal pH values of the CON and GSE groups were within the normal range from 6.45 to 6.53, indicating that Gse supplementation had no negative effects on ruminal fermentation. The difference in research results may be associated with the dietary ratio of concentrate to roughage. Furthermore, calves fed Gse increased MCP content in the rumen. Plant polyphenols can reduce the degradation of protein in the rumen and increase intestinally absorbable dietary protein [
29], which can explain the improvement in ruminal MCP. In the rumen, NH
3-N is the main raw material that synthesizes MCP. An in vitro study reported that polyphenols can reduce the ruminal NH
3-N content [
30]. In our research, the NH
3-N concentration was similar between the two treatments. This suggests that the microbial communities in calves were still in developmental period.
In the present study, Gse supplementation significantly increased the ruminal butyrate content of weaned cross-breed beef calves. Butyrate is the main energy source of the ruminal epithelium, and ruminal development is closely related to the butyrate content produced by microorganisms [
31]. Therefore, the elevated content of butyrate found in this study might be helpful for the ruminal development of weaned cross-breed beef calves. In addition, we found that the ratio of acetate to propionate in the GSE group was reduced compared to the CON group, indicating that Gse supplementation promoted the ruminal propionate fermentation of calves. In ruminants, propionate is an important precursor of gluconeogenesis [
32]. Ruminal propionate fermentation can provide more energy for the body. Moreover, a previous study reported that grape seed meal could up-regulate the relative abundance of butyrate-producing bacteria, which contributed to the increased content of butyrate in the rumen [
33]. In the future, the effects of Gse on microbial communities in the rumen of calves require an in-depth investigation.
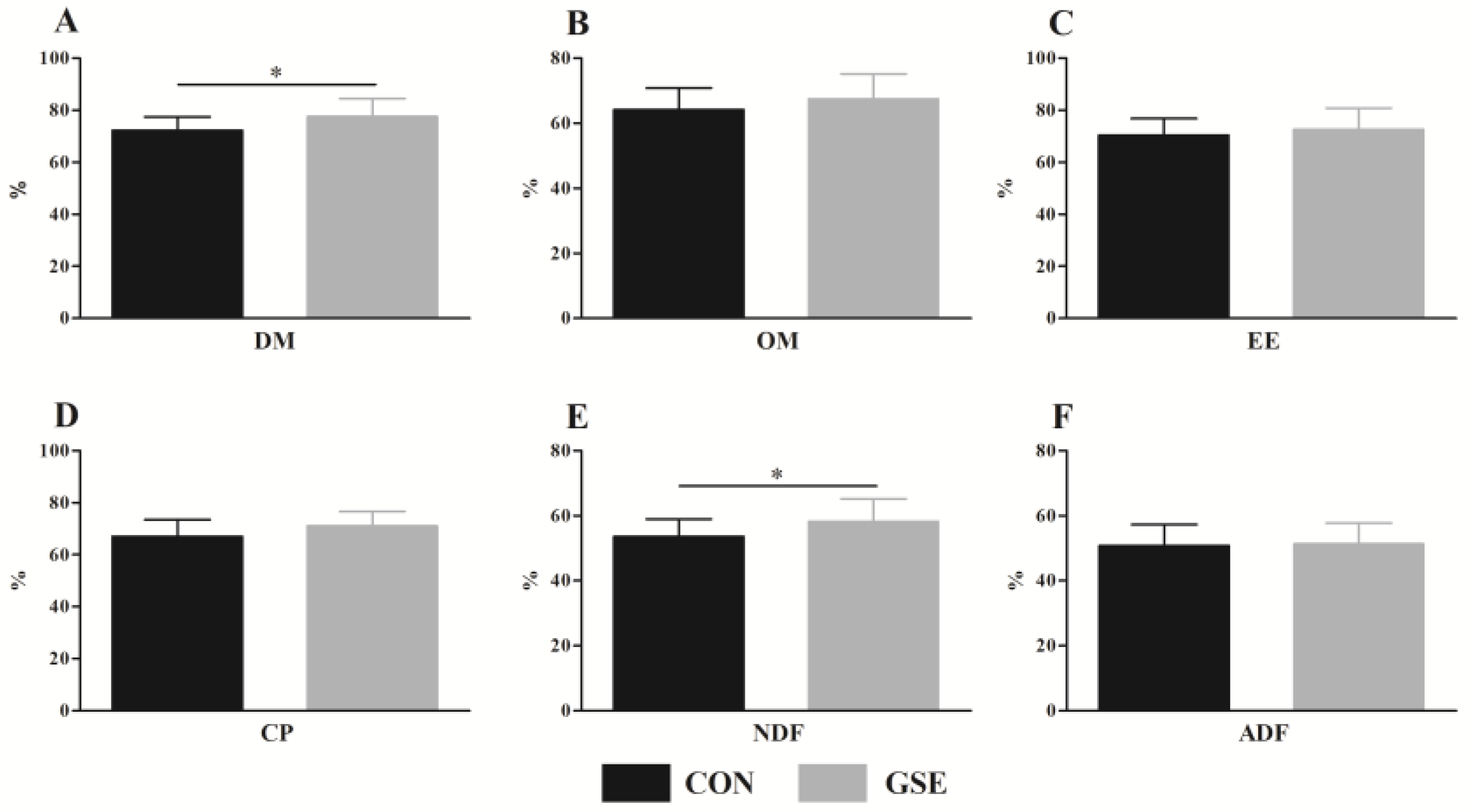
In addition to rumen fermentation, nutrient digestibility could also directly affect the growth performance of calves. DM and OM digestibility are important indexes that can be used to measure animal diets [
34]. In our study, Gse supplementation significantly increased DM digestibility, indicating that the calves in the GSE group could obtain more nutrients and promote growth, which matched with growth performance data. Moreover, the NDF digestibility of calves fed with Gse significantly increased, which was in line with the study by Juráček et al. [
35], who reported that the dietary supplementation of grape pomace could improve the NDF digestibility of wethers. In dairy calves, the supplementation of gallic acid, an ingredient of Gse, in the starter feed of calves, is confirmed to up-regulate the relative abundance of fecal
Ruminococcaceae,
Bacteroides and
Christensenellaceae [
36], which have essential functions in cellulose degradation and VFA production. We speculated that the positive effects of grape seed products on NDF digestibility might be related to the regulation of microbiota in the digestive tract. Forage can be utilized by ruminal microorganisms to generate MCP and small peptides that are easy to absorb by small intestine to increase nutrient digestibility [
37]. Interestingly, Gse supplementation slightly increased the CP digestibility, which was consistent with MCP result mentioned earlier. Generally, improved nutrient digestibility improves ADG, as seen in growth performance results. Lastly, the activity of digestive enzymes plays a vital role in nutrient digestion. Thus, future research should be carried out to elucidate the effects of Gse supplementation on the digestive enzyme activity of calves.
As important parameters associated with the health of animals, serum biochemistry reflects the physiological metabolism of the body and changes in various organ functions [
38]. The serum contents of TP, ALB, GLB, and UN are essential indexes of protein metabolism, and changes in GLU, TG, and NEFA concentrations in serum are closely related to lipid and energy metabolism. Moreover, the ALP, ALT and AST concentrations in serum reflect hepatic functions [
38]. In this study, the serum concentrations of TP, ALB, GLB, UN, ALP, ALT, and AST were similar between two groups, suggesting that Gse supplementation did not have negative effects on protein metabolism and the hepatic function of weaned beef calves. In laboratory animals, one study showed that Gse could protect the liver when its function was experimentally damaged [
39]. In ruminants, Nudda et al. [
40] found that dietary supplementation with grape seed did not have significant effects on UN, ALT, and AST concentrations in blood of dairy ewes, in accordance with our results. Our study also found that Gse supplementation increased serum GLU and TG contents, while decreasing NEFA content in calves, indicating that Gse could improve energy metabolism. The reason for this may be related to the ruminal propionate fermentation caused by Gse supplementation, and ruminal propionate fermentation enhances the gluconeogenesis process, which can result in an elevated GLU content in the sera of calves. However, the potential mechanism of action still needs investigation. TG is the the most abundant and productive energy substance in the body, and under stress, the body will decompose TG into NEFA to supply energy for physiological activities via blood circulation [
41]. Gse supplementation improved the negative energy balance of calves induced by weaning, which was beneficial for promoting growth.
Weaning calves are exposed to oxidative stress [
42]. The GSH-Px, CAT, SOD, MDA, and T-AOC are the most important indexes used to determine the antioxidant ability of animals to protect themselves against oxidative damage [
1]. In this experiment, Gse supplementation increased the serum activity of GSH-Px, CAT, SOD, and T-AOC and reduced MDA concentration, suggesting that Gse could improve the antioxidant ability of weaned cross-breed beef calves. In calves and lambs, previous studies reported that dietary supplementation with grape seed products effectively increased he serum antioxidative enzymes [
25,
43], which is consistent with our results. Plant polyphenols have been demonstrated to protect animals from oxidative damage via the Nrf2/Keap1 signaling pathway [
44]. In addition, catechin, gallic acid and procyanidins, which are the main phenolic ingredients of Gse, inhibit the production of reactive oxygen species and free radicals mainly by scavenging hydrogen peroxide and free radicals and preventing macromolecular damage [
5]. A recent study found that lambs supplemented with grape seed procyanidin had an enhanced activity of CAT, SOD, and T-AOC and reduced MDA content in muscle [
7], which is essentially in line with the results of our study. Our results show that Gse supplementation has positive effects on CAT, SOD, MDA, and T-AOC in calves under weaning stress.
Due to undeveloped digestive and immune functions, calves usually have reduced immunity [
24]. Serum immunoglobulin content reflects the immune function of animals. At present, research on the effects of Gse on the immunity of ruminants remains scare. In our study for heightening the immunologic function of calves. The polyphenols of Gse can inhi, calves fed Gse had increased serum immunoglobulin (IgA, IgG and IgM) contents. Serum immunoglobulins have the key function of humoral immunity. Therefore, elevated immunoglobulin contents were beneficial for the growth of harmful microorganisms, including
Escherichia coli and
Salmonella, in the digestive tract of animals [
26], which might explain the positive influence of Gse on immunity.
After weaning, due to the changes in feeding methods and feedstuffs, the integrity of tight junction protein that forms the gastrointestinal epithelium in animals is damaged. As a result, lipopolysaccharides enter into the blood circulation because of elevated gastrointestinal permeability, cause the release of pro-inflammatory cytokines, and trigger local and systemic inflammation [
45]. As key pro-inflammatory factors, TNF-α, IL-1β, and IL-6 regulate inflammatory reactions of the body. IL-10 can inhibit the production of pro-inflammatory factors and the combination of factors with receptors, thus decreasing the damage of inflammatory reactions in the body [
46]. In the present study, the dietary supplementation with Gse reduced the contents of TNF-α and IL-6 and slightly increased the IL-10 content in serum of calves, indicating that Gse alleviated the inflammatory responses of beef calves. In preweaning dairy calves, Gse is verified to reduce the blood concentration of TNF-α and attenuate the inflammatory response under heat stress condition [
25], which were consistent with our findings. Another study found that grape seed product could decrease the level of pro-inflammatory cytokines by suppressing the NF-κB signaling pathway [
47]. The beneficial influence of Gse on the regulation of inflammatory factors may be achieved by inhibiting the NF-κB signaling pathway. Future research should pay more attention to the potential mechanisms of Gse in the immunity of calves.


 ,
, 



{kind=link}
{kind=link}