
How Do Alternative Protein Resources Affect the Intestine Morphology and Microbiota of Atlantic Salmon?

 , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. The Salmonid’s Gastrointestinal Tract (GIT)
1.1. Macroscopic Aspects of the Intestine
1.1.1. Mid Intestine (MI)
1.1.2. Distal Intestine (DI)
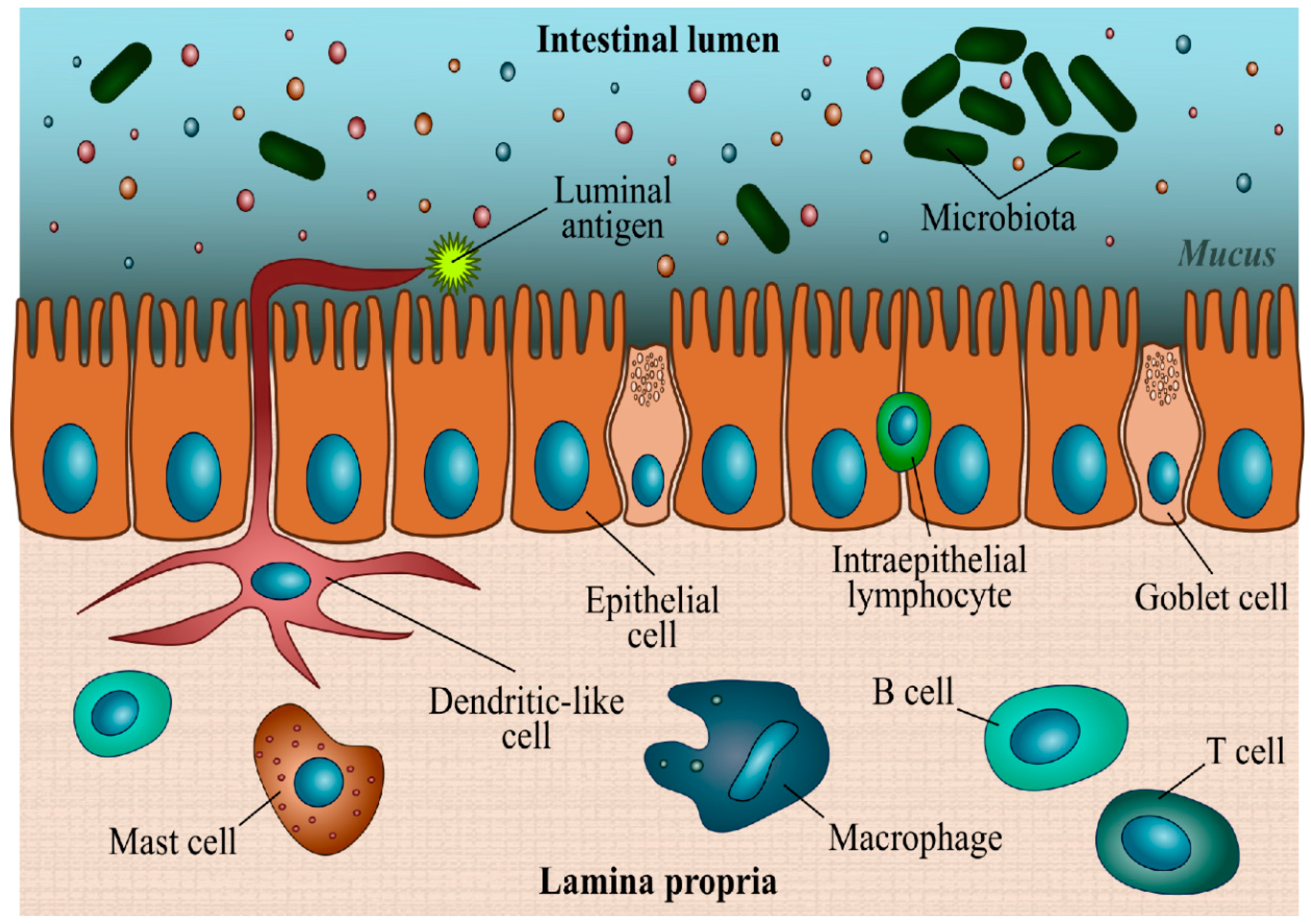
1.2. Microscopic Aspects of the Intestine: Mid and Distal Intestine (MI and DI)
1.3. Microbiota
2. Nutritional Needs of Salmon—“The Protein Problem”
3. Fishmeal (FM) Dietary Replacement—Looking for Potential Candidates

{kind=link}
| Protein Source | Inclusion Level | Mucosa Folds | Mucosa Thickness | Mucosa Cells | Digestive/Absorption Capacity | Inflammatory Process | Overall Histology | References |
|---|---|---|---|---|---|---|---|---|
| Soybean Meal | 10% | ↓ | [50] | |||||
| 10% | − | − | − | ↑ | [51] | |||
| 15% | −− | −− | ↓↓ | [50] | ||||
| 20% | −−− | −−− | ↓↓↓ | [50] | ||||
| 20% | −− | −− | −− | ↓ | ↑↑ | [51] | ||
| 20% | − | ↑ | [52] | |||||
| 20% | − | − | [53] | |||||
| 20% * | + | + | [54] | |||||
| 25% | −−−− | −−−− | ↓↓↓↓ | [50] | ||||
| 30% | − | ↓ | ↑ | − | [55] | |||
| 30% | − | − | [49] | |||||
| 32% | − | − | ↓ | ↑↑ | − | [56,57] | ||
| 35% | − | − | − | ↑ | [58] | |||
| 35% | −−−− | −−−− | ↓↓↓↓↓ | [50] | ||||
| 40% | −− | −− | − | − | [57] | |||
| 50% | − | − | − | ↑ | [59] | |||
| HP48 | 17.8% | − | − | − | [60] | |||
| Whole Field Pea Pea Protein Concentrate | 20% | = | = | = | = | = | [45] | |
| 20% | = | ↓ | = | [61] | ||||
| 35% | − | − | − | ↓ | ↑ | − | [43] | |
| 44% | = | = | = | = | [62] | |||
| Dehulled Faba Faba Bean Concentrate | 20% | = | = | = | = | = | [45] | |
| 17% | − | − | − | ↓ | ↑ | [63] | ||
| 5–20% | = | = | = | ↓ | = | [64] | ||
| 35% | −− | ↓↓ | ↑↑ | [63] | ||||
| 45% | = | = | − | ↑ | − | [64] | ||
| Plant Proteins | 25% | = | = | = | ↑ | [65] | ||
| Poultry Meal | 20% | − | = | [66] | ||||
| 58% | = | = | = | = | [67] | |||
| Feather Meal | 20% | − | = | [62] | ||||
| Insect Meal | 5–15% | = | = | = | [68] | |||
| 5–15% | = | = | [69] | |||||
| 6.25–25% | = | = | = | + | [70] | |||
| 15% | − | − | ↑ | + | [71] | |||
| 25% | = | = | = | = | [17] | |||
| 85% | = | = | = | = | [71] | |||
| Microalgae | ||||||||
| Nannochloropsis oceanica | 20% | = | = | = | ↓ | = | = | [72] |
| Desmodesmus sp. | 20% | = | = | = | = | [73] | ||
| Yeasts | ||||||||
| Candida utilis | 20% | + | [74] | |||||
| 25% | = | = | = | = | [23] |
| Protein Source | Concentration | Intestinal Tract | Effects on Microbiota (MB) | Microbial Population Mostly Influenced | Enteritis | References |
|---|---|---|---|---|---|---|
| Soybean meal | 5–10% | PI and DI mucosa | Yes | Presence of bacteria not normally associated with marine fish (Escherichia and Propionibacterium) | [75] | |
| 20% | MI and DI mucosa | No | Tenericutes dominant Phylum Proteobacteria, Firmicutes and Bacteroidetes. Lactic acid bacteria increase | + | [52] | |
| 20% | MI and DI digesta | Yes | Abundance of Firmicutes and Bacteroides; increase in Proteobacteria | + | [76] | |
| 25% | MI and DI mucosa and digesta | Yes | Increase in Enterococcus spp., mostly Enterococcus faecalis | [41] | ||
| 30% | DI mucosa and digesta | Yes | Proteobacteria, Firmicutes, and Bacteroidetes Abundance of lactic acid bacteria | + | [49] | |
| 30% | DI digesta | No | Dominated by lactic acid bacteria | [77] | ||
| 50% | DI digesta | Yes | Shewanella dominant bacteria. Lactococcus, Aeromonas, Pseudomonas. Lactic acid bacteria may modulate intestinal inflammation | [59] | ||
| Pea protein concentrate | 20% | Allochthonous bacteria: Fecal samples Autochthonous Bacteria: PI and DI mucosa | Yes | Corynebacteriaceaein dominant in the allochthonous bacterial population and Lactobacillaceaein in the autochthonous | [62] | |
| Poultry by-product meal | 20% | Allochthonous bacteria: Fecal samples Autochthonous Bacteria: PI and DI mucosa | Yes | Corynebacteriaceaein dominant in the allochthonous bacterial population and Lactobacillaceaein in the autochthonous | [62] | |
| Insect meal | 60% | DI digesta and mucosa | yes | Increasing of Actinomyces, Bacillus, Brevibacterium, Corynebacterium 1 and Enterococcus | [78] | |
| Yeast Cyberlindnera jadinii | 10% | MI and DI mucosa | yes | Weissella and Leuconostoc slightly increased. Lactobacillus decrease | [79] | |
| 20% |
3.1. Soybean Meal (SBM) and Soy Protein Concentrate
3.2. Peas and Pea Protein Concentrate (PPC)
3.3. Faba Bean and Faba Bean Protein Concentrate (BPC)
3.4. Other Plant Protein Sources (PPS)
3.5. Animal Protein: Poultry by-Product Meal (PBM)
3.6. Animal Protein: Insect Meal (IM)
3.6.1. Black Soldier Fly (BSF-Hermetia Illucens)
3.6.2. Yellow Mealworm (Tenebrio Molitor)
3.7. Single-Cell Protein (SCP)
3.7.1. Microalgae
3.7.2. Yeasts
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Løkka, G.; Austbø, L.; Falk, K.; Bjerkås, I.; Koppang, E.O. Intestinal Morphology of the Wild Atlantic Salmon (Salmo salar). J. Morphol. 2013, 274, 859–876. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, B.G.; Smit, H.; Verighina, I.A. The Alimentary Canal and Digestion in Teleosts. In Advances in Marine Biology; Russell, F.S., Yonge, M., Eds.; Academic Press: Cambridge, MA, USA, 1976; Volume 13, pp. 109–239. [Google Scholar]
- Bjørgen, H.; Li, Y.; Kortner, T.M.; Krogdahl, Å.; Koppang, E.O. Anatomy, Immunology, Digestive Physiology and Microbiota of the Salmonid Intestine: Knowns and Unknowns under the Impact of an Expanding Industrialized Production. Fish Shellfish Immunol. 2020, 107, 172–186. [Google Scholar] [CrossRef] [PubMed]
- Harder, W. The Digestive Tract. In Anatomy of Fishes. Part 1: Text; E. Schweizerbart’sche Verlagsbuchhandlung: Stuttgart, Germany, 1975; Volume 62, pp. 128–186. [Google Scholar]
- Rust, M.B. 7—Nutritional Physiology. In Fish Nutrition, 3rd ed.; Halver, J.E., Hardy, R.W., Eds.; Academic Press: San Diego, CA, USA, 2003; pp. 367–452. ISBN 978-0-12-319652-1. [Google Scholar]
- Denstadli, V.; Vegusdal, A.; Krogdahl, Å.; Bakke-McKellep, A.M.; Berge, G.M.; Holm, H.; Hillestad, M.; Ruyter, B. Lipid Absorption in Different Segments of the Gastrointestinal Tract of Atlantic Salmon (Salmo salar L.). Aquaculture 2004, 240, 385–398. [Google Scholar] [CrossRef]
- Burnstock, G. The Morphology of the Gut of the Brown Trout (Salmo trutta). J. Cell Sci. 1959, s3-100, 183–198. [Google Scholar] [CrossRef]
- Sundell, K.; Jutfelt, F.; Ágústsson, T.; Olsen, R.-E.; Sandblom, E.; Hansen, T.; Björnsson, B.T. Intestinal Transport Mechanisms and Plasma Cortisol Levels during Normal and Out-of-Season Parr–Smolt Transformation of Atlantic Salmon. Salmo salar. Aquac. 2003, 222, 265–285. [Google Scholar] [CrossRef]
- Sire, M.F.; Vernier, J.-M. Intestinal Absorption of Protein in Teleost Fish. Comp. Biochem. Physiol. A Physiol. 1992, 103, 771–781. [Google Scholar] [CrossRef]
- Kent, G.; Carr, R. Digestive System. In Comparative Anatomy of the Vertebrates; Kent, G.C., Carr, R.K., Eds.; McGraw-Hill Publishing: New York, NY, USA, 2001; pp. 265–291. [Google Scholar]
- Jutfelt, F. Barrier Function of the Gut. In Encyclopedia of Fish Physiology from Genome to Environment; Academic Press: Cambridge, MA, USA, 2011. [Google Scholar]
- Takei, Y.; Loretz, C.A. 7—The Gastrointestinal Tract as an Endocrine/Neuroendocrine/Paracrine Organ: Organization, Chemical Messengers and Physiological Targets. In Fish Physiology; Grosell, M., Farrell, A.P., Brauner, C.J., Eds.; The Multifunctional Gut of Fish; Academic Press: Cambridge, MA, USA, 2010; Volume 30, pp. 261–317. [Google Scholar]
- Wilson, J.M.; Castro, L.F.C. 1—Morphological Diversity of the Gastrointestinal Tract in Fishes. In Fish Physiology; Grosell, M., Farrell, A.P., Brauner, C.J., Eds.; The Multifunctional Gut of Fish; Academic Press: Cambridge, MA, USA, 2010; Volume 30, pp. 1–55. [Google Scholar]
- Grosell, M. Intestinal Anion Exchange in Marine Teleosts Is Involved in Osmoregulation and Contributes to the Oceanic Inorganic Carbon Cycle. Acta Physiol. 2011, 202, 421–434. [Google Scholar] [CrossRef]
- Pascoli, F.; Negrato, E.; Di Giancamillo, A.; Bertotto, D.; Domeneghini, C.; Simontacchi, C.; Mutinelli, F.; Radaelli, G. Evaluation of Oxidative Stress Biomarkers in Zosterisessor Ophiocephalus from the Venice Lagoon, Italy. Aquat. Toxicol. 2011, 101, 512–520. [Google Scholar] [CrossRef]
- Poussier, P.; Edouard, P.; Lee, C.; Binnie, M.; Julius, M. Thymus-Independent Development and Negative Selection of T Cells Expressing T Cell Receptor Alpha/Beta in the Intestinal Epithelium: Evidence for Distinct Circulation Patterns of Gut- and Thymus-Derived T Lymphocytes. J. Exp. Med. 1992, 176, 187–199. [Google Scholar] [CrossRef] [Green Version]
- Lock, E.R.; Arsiwalla, T.; Waagbø, R. Insect Larvae Meal as an Alternative Source of Nutrients in the Diet of Atlantic Salmon (Salmo salar) Postsmolt. Aquac. Nutr. 2016, 22, 1202–1213. [Google Scholar] [CrossRef]
- Bjørgen, H.; Hellberg, H.; Løken, O.M.; Gunnes, G.; Koppang, E.O.; Dale, O.B. Tumor Microenvironment and Stroma in Intestinal Adenocarcinomas and Associated Metastases in Atlantic Salmon Broodfish (Salmo salar). Vet. Immunol. Immunopathol. 2019, 214, 109891. [Google Scholar] [CrossRef]
- Tafalla, C.; Leal, E.; Yamaguchi, T.; Fischer, U. T Cell Immunity in the Teleost Digestive Tract. Dev. Comp. Immunol. 2016, 64, 167–177. [Google Scholar] [CrossRef]
- Ng, A.N.Y.; de Jong-Curtain, T.A.; Mawdsley, D.J.; White, S.J.; Shin, J.; Appel, B.; Dong, P.D.S.; Stainier, D.Y.R.; Heath, J.K. Formation of the Digestive System in Zebrafish: III. Intestinal Epithelium Morphogenesis. Dev. Biol. 2005, 286, 114–135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallace, K.N.; Akhter, S.; Smith, E.M.; Lorent, K.; Pack, M. Intestinal Growth and Differentiation in Zebrafish. Mech. Dev. 2005, 122, 157–173. [Google Scholar] [CrossRef] [PubMed]
- Whittamore, J.M. Osmoregulation and Epithelial Water Transport: Lessons from the Intestine of Marine Teleost Fish. J. Comp. Physiol. B 2012, 182, 1–39. [Google Scholar] [CrossRef] [PubMed]
- Sahlmann, C.; Djordjevic, B.; Lagos, L.; Mydland, L.T.; Morales-Lange, B.; Øvrum Hansen, J.; Ånestad, R.; Mercado, L.; Bjelanovic, M.; Press, C.M.; et al. Yeast as a Protein Source during Smoltification of Atlantic Salmon (Salmo salar L.), Enhances Performance and Modulates Health. Aquaculture 2019, 513, 734396. [Google Scholar] [CrossRef]
- Dhanasiri, A.K.S.; Brunvold, L.; Brinchmann, M.F.; Korsnes, K.; Bergh, Ø.; Kiron, V. Changes in the Intestinal Microbiota of Wild Atlantic Cod Gadus Morhua L. Upon Captive Rearing. Microb. Ecol. 2011, 61, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Ringø, E.; Zhou, Z.; Vecino, J.L.G.; Wadsworth, S.; Romero, J.; Krogdahl, Å.; Olsen, R.e.; Dimitroglou, A.; Foey, A.; Davies, S.; et al. Effect of Dietary Components on the Gut Microbiota of Aquatic Animals. A Never-Ending Story? Aquac. Nutr. 2016, 22, 219–282. [Google Scholar] [CrossRef] [Green Version]
- Ringø, E.; Olsen, R.E.; Mayhew, T.M.; Myklebust, R. Electron Microscopy of the Intestinal Microflora of Fish. Aquaculture 2003, 227, 395–415. [Google Scholar] [CrossRef]
- Wang, J.; Jaramillo-Torres, A.; Li, Y.; Kortner, T.M.; Gajardo, K.; Brevik, Ø.J.; Jakobsen, J.V.; Krogdahl, Å. Microbiota in Intestinal Digesta of Atlantic Salmon (Salmo salar), Observed from Late Freshwater Stage until One Year in Seawater, and Effects of Functional Ingredients: A Case Study from a Commercial Sized Research Site in the Arctic Region. Anim. Microbiome 2021, 3, 14. [Google Scholar] [CrossRef]
- Falcinelli, S.; Picchietti, S.; Rodiles, A.; Cossignani, L.; Merrifield, D.L.; Taddei, A.R.; Maradonna, F.; Olivotto, I.; Gioacchini, G.; Carnevali, O. Lactobacillus Rhamnosus Lowers Zebrafish Lipid Content by Changing Gut Microbiota and Host Transcription of Genes Involved in Lipid Metabolism. Sci. Rep. 2015, 5, 9336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Semova, I.; Carten, J.D.; Stombaugh, J.; Mackey, L.C.; Knight, R.; Farber, S.A.; Rawls, J.F. Microbiota Regulate Intestinal Absorption and Metabolism of Fatty Acids in the Zebrafish. Cell Host Microbe 2012, 12, 277–288. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ray, A.K.; Ghosh, K.; Ringø, E. Enzyme-Producing Bacteria Isolated from Fish Gut: A Review. Aquac. Nutr. 2012, 18, 465–492. [Google Scholar] [CrossRef]
- Austin, B. The Bacterial Microflora of Fish, Revised. Sci. World J. 2006, 6, 931–945. [Google Scholar] [CrossRef] [Green Version]
- López Nadal, A.; Ikeda-Ohtsubo, W.; Sipkema, D.; Peggs, D.; McGurk, C.; Forlenza, M.; Wiegertjes, G.F.; Brugman, S. Feed, Microbiota, and Gut Immunity: Using the Zebrafish Model to Understand Fish Health. Front. Immunol. 2020, 11, 114. [Google Scholar]
- FAO: Nutritional Requirements. Available online: https://www.fao.org/fishery/affris/species-profiles/atlantic-salmon/nutritional-requirements/en/ (accessed on 19 May 2023).
- Lall, S.P.; Anderson, S. Amino Acid Nutrition of Salmonids: Dietary Requirements and Bioavailablity. Cah. Options Mediterr. 2005, 63, 73–90. [Google Scholar]
- Francis, G.; Makkar, H.P.S.; Becker, K. Antinutritional Factors Present in Plant-Derived Alternate Fish Feed Ingredients and Their Effects in Fish. Aquaculture 2001, 199, 197–227. [Google Scholar] [CrossRef]
- Gatlin, D.M.; Barrows, F.T.; Brown, P.; Dabrowski, K.; Gaylord, T.G.; Hardy, R.W.; Herman, E.; Hu, G.; Krogdahl, Å.; Nelson, R.; et al. Expanding the Utilization of Sustainable Plant Products in Aquafeeds: A Review. Aquac. Res. 2007, 38, 551–579. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). Fisheries and Aquaculture Department—Species Fact Sheets. Available online: www.fao.org (accessed on 19 May 2023).
- Crespi, V. The State of World Fisheries and Aquaculture; FAO: Rome, Italy, 2020; ISBN 978-92-5-132692-3. [Google Scholar]
- Tacon, A.G.J.; Metian, M. Global Overview on the Use of Fish Meal and Fish Oil in Industrially Compounded Aquafeeds: Trends and Future Prospects. Aquaculture 2008, 285, 146–158. [Google Scholar] [CrossRef]
- Gillund, F.; Myhr, A.I. Perspectives on Salmon Feed: A Deliberative Assessment of Several Alternative Feed Resources. J. Agric. Environ. Ethics 2010, 23, 527–550. [Google Scholar] [CrossRef]
- Bakke-McKellep, A.M.; Koppang, E.O.; Gunnes, G.; Sanden, M.; Hemre, G.-I.; Landsverk, T.; Krogdahl, A. Histological, Digestive, Metabolic, Hormonal and Some Immune Factor Responses in Atlantic Salmon, Salmo salar L., Fed Genetically Modified Soybeans. J. Fish Dis. 2007, 30, 65–79. [Google Scholar] [CrossRef] [PubMed]
- Dale, O.B.; Tørud, B.; Kvellestad, A.; Koppang, H.S.; Koppang, E.O. From Chronic Feed-Induced Intestinal Inflammation to Adenocarcinoma with Metastases in Salmonid Fish. Cancer Res. 2009, 69, 4355–4362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Penn, M.H.; Bendiksen, E.Å.; Campbell, P.; Krogdahl, Å. High Level of Dietary Pea Protein Concentrate Induces Enteropathy in Atlantic Salmon (Salmo salar L.). Aquaculture 2011, 310, 267–273. [Google Scholar] [CrossRef]
- Urán, P.A.; Schrama, J.W.; Rombout, J.H.W.M.; Obach, A.; Jensen, L.; Koppe, W.; Verreth, J.A.J. Soybean Meal-Induced Enteritis in Atlantic Salmon (Salmo salar L.) at Different Temperatures. Aquac. Nutr. 2008, 14, 324–330. [Google Scholar] [CrossRef]
- Aslaksen, M.A.; Kraugerud, O.F.; Penn, M.; Svihus, B.; Denstadli, V.; Jørgensen, H.Y.; Hillestad, M.; Krogdahl, Å.; Storebakken, T. Screening of Nutrient Digestibilities and Intestinal Pathologies in Atlantic Salmon, Salmo salar, Fed Diets with Legumes, Oilseeds, or Cereals. Aquaculture 2007, 272, 541–555. [Google Scholar] [CrossRef]
- Thiessen, D.L.; Campbell, G.L.; Adelizi, P.D. Digestibility and Growth Performance of Juvenile Rainbow Trout (Oncorhynchus Mykiss) Fed with Pea and Canola Products. Aquac. Nutr. 2003, 9, 67–75. [Google Scholar] [CrossRef]
- Gouveia, A.; Davies, S.J. Inclusion of an Extruded Dehulled Pea Seed Meal in Diets for Juvenile European Sea Bass (Dicentrarchus Labrax). Aquaculture 2000, 182, 183–193. [Google Scholar] [CrossRef]
- Shubin, A.V.; Demidyuk, I.V.; Komissarov, A.A.; Rafieva, L.M.; Kostrov, S.V. Cytoplasmic Vacuolization in Cell Death and Survival. Oncotarget 2016, 7, 55863–55889. [Google Scholar] [CrossRef] [Green Version]
- Gajardo, K.; Jaramillo-Torres, A.; Kortner, T.M.; Merrifield, D.L.; Tinsley, J.; Bakke, A.M.; Krogdahl, Å. Alternative Protein Sources in the Diet Modulate Microbiota and Functionality in the Distal Intestine of Atlantic Salmon (Salmo salar). Appl. Environ. Microbiol. 2016, 83, e02615-16. [Google Scholar] [CrossRef] [Green Version]
- Krogdahl, Å.; Bakke-McKellep, A.M.; Baeverfjord, G. Effects of Graded Levels of Standard Soybean Meal on Intestinal Structure, Mucosal Enzyme Activities, and Pancreatic Response in Atlantic Salmon (Salmo salar L.). Aquac. Nutr. 2003, 9, 361–371. [Google Scholar] [CrossRef]
- Urán, P.A.; Schrama, J.W.; Rombout, J.H.W.M.; Taverne-Thiele, J.J.; Obach, A.; Koppe, W.; Verreth, J. a J. Time-Related Changes of the Intestinal Morphology of Atlantic Salmon, Salmo salar L., at Two Different Soybean Meal Inclusion Levels. J. Fish Dis. 2009, 32, 733–744. [Google Scholar] [CrossRef] [PubMed]
- Booman, M.; Forster, I.; Vederas, J.C.; Groman, D.B.; Jones, S.R.M. Soybean Meal-Induced Enteritis in Atlantic Salmon (Salmo salar) and Chinook Salmon (Oncorhynchus tshawytscha) but Not in Pink Salmon (O. Gorbuscha). Aquaculture 2018, 483, 238–243. [Google Scholar] [CrossRef]
- Reveco-Urzua, F.E.; Hofossæter, M.; Kovi, M.R.; Mydland, L.T.; Ånestad, R.; Sørby, R.; Press, C.M.; Lagos, L.; Øverland, M. Candida Utilis Yeast as a Functional Protein Source for Atlantic Salmon (Salmo salar L.): Local Intestinal Tissue and Plasma Proteome Responses. PLoS ONE 2019, 14, e0218360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimalan, N.; Sørensen, S.L.; Fečkaninová, A.; Koščová, J.; Mudroňová, D.; Gancarčíková, S.; Vatsos, I.N.; Bisa, S.; Kiron, V.; Sørensen, M. Supplementation of Lactic Acid Bacteria Has Positive Effects on the Mucosal Health of Atlantic Salmon (Salmo salar) Fed Soybean Meal. Aquac. Rep. 2023, 28, 101461. [Google Scholar] [CrossRef]
- Nordrum, S.; Bakke-McKellep, A.M.; Krogdahl, A.; Buddington, R.K. Effects of Soybean Meal and Salinity on Intestinal Transport of Nutrients in Atlantic Salmon (Salmo salar L.) and Rainbow Trout (Oncorhynchus mykiss). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2000, 125, 317–335. [Google Scholar] [CrossRef]
- Refstie, S.; Baeverfjord, G.; Seim, R.R.; Elvebø, O. Effects of Dietary Yeast Cell Wall β-Glucans and MOS on Performance, Gut Health, and Salmon Lice Resistance in Atlantic Salmon (Salmo salar) Fed Sunflower and Soybean Meal. Aquaculture 2010, 305, 109–116. [Google Scholar] [CrossRef]
- Baeverfjord, G.; Krogdahl, A. Development and Regression of Soybean Meal Induced Enteritis in Atlantic Salmon, Salmo salar L., Distal Intestine: A Comparison with the Intestines of Fasted Fish. J. Fish Dis. 1996, 19, 375–387. [Google Scholar] [CrossRef]
- Bakke-McKellep, A.M.; Press, C.M.; Baeverfjord, G.; Krogdahl, Å.; Landsverk, T. Changes in Immune and Enzyme Histochemical Phenotypes of Cells in the Intestinal Mucosa of Atlantic Salmon, Salmo salar L., with Soybean Meal-Induced Enteritis. J. Fish Dis. 2000, 23, 115–127. [Google Scholar] [CrossRef]
- Navarrete, P.; Fuentes, P.; la Fuente, L.D.; Barros, L.; Magne, F.; Opazo, R.; Ibacache, C.; Espejo, R.; Romero, J. Short-Term Effects of Dietary Soybean Meal and Lactic Acid Bacteria on the Intestinal Morphology and Microbiota of Atlantic Salmon (Salmo salar). Aquac. Nutr. 2013, 19, 827–836. [Google Scholar] [CrossRef]
- Nordvi, M.F.; Løvmo, S.D.; Whatmore, P.; Sundh, H.; Sigholt, T.; Olsen, R.E. Low Intestinal Inflammation Model (HP48) in Atlantic Salmon (Salmo salar) and Inflammatory Mitigation by Bactocell. Aquaculture 2023, 563, 738920. [Google Scholar] [CrossRef]
- Øverland, M.; Sørensen, M.; Storebakken, T.; Penn, M.; Krogdahl, Å.; Skrede, A. Pea Protein Concentrate Substituting Fish Meal or Soybean Meal in Diets for Atlantic Salmon (Salmo salar)—Effect on Growth Performance, Nutrient Digestibility, Carcass Composition, Gut Health, and Physical Feed Quality. Aquaculture 2009, 288, 305–311. [Google Scholar] [CrossRef]
- Hartviksen, M.; Vecino, J.L.G.; Ringø, E.; Bakke, A.-M.; Wadsworth, S.; Krogdahl, Å.; Ruohonen, K.; Kettunen, A. Alternative Dietary Protein Sources for Atlantic Salmon (Salmo salar L.) Effect on Intestinal Microbiota, Intestinal and Liver Histology and Growth. Aquac. Nutr. 2014, 20, 381–398. [Google Scholar] [CrossRef] [Green Version]
- De Santis, C.; Martin, S.A.M.; Dehler, C.E.; Iannetta, P.P.M.; Leeming, D.; Tocher, D.R. Influence of Dietary Inclusion of a Wet Processed Faba Bean Protein Isolate on Post-Smolt Atlantic Salmon (Salmo salar). Aquaculture 2016, 465, 124–133. [Google Scholar] [CrossRef]
- De Santis, C.; Ruohonen, K.; Tocher, D.R.; Martin, S.A.M.; Król, E.; Secombes, C.J.; Bell, J.G.; El-Mowafi, A.; Crampton, V.O. Atlantic Salmon (Salmo salar) Parr as a Model to Predict the Optimum Inclusion of Air Classified Faba Bean Protein Concentrate in Feeds for Seawater Salmon. Aquaculture 2015, 444, 70–78. [Google Scholar] [CrossRef]
- Tacchi, L.; Secombes, C.J.; Bickerdike, R.; Adler, M.A.; Venegas, C.; Takle, H.; Martin, S.A. Transcriptomic and Physiological Responses to Fishmeal Substitution with Plant Proteins in Formulated Feed in Farmed Atlantic Salmon (Salmo salar). BMC Genom. 2012, 13, 363. [Google Scholar] [CrossRef] [Green Version]
- Hartviksen, M.; Bakke, A.M.; Vecino, J.G.; Ringø, E.; Krogdahl, Å. Evaluation of the Effect of Commercially Available Plant and Animal Protein Sources in Diets for Atlantic Salmon (Salmo salar L.): Digestive and Metabolic Investigations. Fish Physiol. Biochem. 2014, 40, 1621–1637. [Google Scholar] [CrossRef]
- Hu, H.; Kortner, T.M.; Gajardo, K.; Chikwati, E.; Tinsley, J.; Krogdahl, Å. Intestinal Fluid Permeability in Atlantic Salmon (Salmo salar L.) Is Affected by Dietary Protein Source. PLoS ONE 2016, 11, e0167515. [Google Scholar] [CrossRef] [Green Version]
- Mikołajczak, Z.; Mazurkiewicz, J.; Rawski, M.; Kierończyk, B.; Józefiak, A.; Świątkiewicz, S.; Józefiak, D. Black Soldier Fly Full-Fat Meal in Atlantic Salmon Nutrition—Part B: Effects on Growth Performance, Feed Utilization, Selected Nutriphysiological Traits and Production Sustainability in Pre-Smolts. Ann. Anim. Sci. 2022. ahead-of-print. [Google Scholar] [CrossRef]
- Mikołajczak, Z.; Mazurkiewicz, J.; Rawski, M.; Kierończyk, B.; Józefiak, A.; Świątkiewicz, S.; Józefiak, D. Black Soldier Fly Full-Fat Meal in Atlantic Salmon Nutrition—Part A: Effects on Growth Performance, Feed Utilization, Selected Nutriphysiological Traits and Production Sustainability in Fries. Ann. Anim. Sci. 2022. ahead-of-print. [Google Scholar] [CrossRef]
- Weththasinghe, P.; Lagos, L.; Cortés, M.; Hansen, J.Ø.; Øverland, M. Dietary Inclusion of Black Soldier Fly (Hermetia illucens) Larvae Meal and Paste Improved Gut Health but Had Minor Effects on Skin Mucus Proteome and Immune Response in Atlantic Salmon (Salmo salar). Front. Immunol. 2021, 12, 599530. [Google Scholar] [CrossRef]
- Li, Y.; Kortner, T.M.; Chikwati, E.M.; Belghit, I.; Lock, E.-J.; Krogdahl, Å. Total Replacement of Fish Meal with Black Soldier Fly (Hermetia Illucens) Larvae Meal Does Not Compromise the Gut Health of Atlantic Salmon (Salmo salar). Aquaculture 2020, 520, 734967. [Google Scholar] [CrossRef]
- Sørensen, S.L.; Ghirmay, A.; Gong, Y.; Dahle, D.; Vasanth, G.; Sørensen, M.; Kiron, V. Growth, Chemical Composition, Histology and Antioxidant Genes of Atlantic Salmon (Salmo salar) Fed Whole or Pre-Processed Nannochloropsis Oceanica and Tetraselmis Sp. Fishes 2021, 6, 23. [Google Scholar] [CrossRef]
- Kiron, V.; Sørensen, M.; Huntley, M.; Vasanth, G.K.; Gong, Y.; Dahle, D.; Palihawadana, A.M. Defatted Biomass of the Microalga, Desmodesmus Sp., Can Replace Fishmeal in the Feeds for Atlantic Salmon. Front. Mar. Sci. 2016, 3, 67. [Google Scholar] [CrossRef] [Green Version]
- Hansen, J.; Hofossæter, M.; Sahlmann, C.; Ånestad, R.; Reveco-Urzua, F.E.; Press, C.M.; Mydland, L.T.; Øverland, M. Effect of Candida Utilis on Growth and Intestinal Health of Atlantic Salmon (Salmo salar) Parr. Aquaculture 2019, 511, 734239. [Google Scholar] [CrossRef]
- Green, T.J.; Smullen, R.; Barnes, A.C. Dietary Soybean Protein Concentrate-Induced Intestinal Disorder in Marine Farmed Atlantic Salmon, Salmo salar Is Associated with Alterations in Gut Microbiota. Vet. Microbiol. 2013, 166, 286–292. [Google Scholar] [CrossRef] [PubMed]
- Reveco, F.E.; Øverland, M.; Romarheim, O.H.; Mydland, L.T. Intestinal Bacterial Community Structure Differs between Healthy and Inflamed Intestines in Atlantic Salmon (Salmo salar L.). Aquaculture 2014, 420–421, 262–269. [Google Scholar] [CrossRef]
- Agboola, J.O.; Rocha, S.D.C.; Mensah, D.D.; Hansen, J.Ø.; Øyås, O.; Lapeña, D.; Mydland, L.T.; Arntzen, M.Ø.; Horn, S.J.; Øverland, M. Effect of Yeast Species and Processing on Intestinal Microbiota of Atlantic Salmon (Salmo salar) Fed Soybean Meal-Based Diets in Seawater. Anim. Microbiome 2023, 5, 21. [Google Scholar] [CrossRef]
- Li, Y.; Gajardo, K.; Jaramillo-Torres, A.; Kortner, T.M.; Krogdahl, Å. Consistent Changes in the Intestinal Microbiota of Atlantic Salmon Fed Insect Meal Diets. Anim. Microbiome 2022, 4, 8. [Google Scholar] [CrossRef]
- Leeper, A.; Ekmay, R.; Knobloch, S.; Skírnisdóttir, S.; Varunjikar, M.; Dubois, M.; Smárason, B.Ö.; Árnason, J.; Koppe, W.; Benhaïm, D. Torula Yeast in the Diet of Atlantic Salmon Salmo salar and the Impact on Growth Performance and Gut Microbiome. Sci. Rep. 2022, 12, 567. [Google Scholar] [CrossRef]
- Urán, P.A.; Aydin, R.; Schrama, J.W.; Verreth, J.A.J.; Rombout, J.H.W.M. Soybean Meal-Induced Uptake Block in Atlantic Salmon Salmo salar Distal Enterocytes. J. Fish Biol. 2008, 73, 2571–2579. [Google Scholar] [CrossRef]
- Bruce, T.J.; Neiger, R.D.; Brown, M.L. Gut Histology, Immunology and the Intestinal Microbiota of Rainbow Trout, Oncorhynchus Mykiss (Walbaum), Fed Process Variants of Soybean Meal. Aquac. Res. 2018, 49, 492–504. [Google Scholar] [CrossRef]
- Miao, S.; Zhao, C.; Zhu, J.; Hu, J.; Dong, X.; Sun, L. Dietary Soybean Meal Affects Intestinal Homoeostasis by Altering the Microbiota, Morphology and Inflammatory Cytokine Gene Expression in Northern Snakehead. Sci. Rep. 2018, 8, 113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Merrifield, D.L.; Dimitroglou, A.; Bradley, G.; Baker, R.T.M.; Davies, S.J. Soybean Meal Alters Autochthonous Microbial Populations, Microvilli Morphology and Compromises Intestinal Enterocyte Integrity of Rainbow Trout, Oncorhynchus mykiss (Walbaum). J. Fish Dis. 2009, 32, 755–766. [Google Scholar] [CrossRef] [PubMed]
- Buttle, L.G.; Burrells, A.C.; Good, J.E.; Williams, P.D.; Southgate, P.J.; Burrells, C. The Binding of Soybean Agglutinin (SBA) to the Intestinal Epithelium of Atlantic Salmon, Salmo salar and Rainbow Trout, Oncorhynchus Mykiss, Fed High Levels of Soybean Meal. Vet. Immunol. Immunopathol. 2001, 80, 237–244. [Google Scholar] [CrossRef] [PubMed]
- Gajardo, K.; Rodiles, A.; Kortner, T.M.; Krogdahl, Å.; Bakke, A.M.; Merrifield, D.L.; Sørum, H. A High-Resolution Map of the Gut Microbiota in Atlantic Salmon (Salmo salar): A Basis for Comparative Gut Microbial Research. Sci. Rep. 2016, 6, 30893. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sanden, M.; Berntssen, M.H.G.; Krogdahl, Å.; Hemre, G.-I.; Bakke-McKellep, A.-M. An Examination of the Intestinal Tract of Atlantic Salmon, Salmo salar L., Parr Fed Different Varieties of Soy and Maize. J. Fish Dis. 2005, 28, 317–330. [Google Scholar] [CrossRef] [PubMed]
- Krogdahl; Bakke-Mckellep; RØed; Baeverfjord. Feeding Atlantic Salmon Salmo salar L. Soybean Products: Effects on Disease Resistance (Furunculosis), and Lysozyme and IgM Levels in the Intestinal Mucosa. Aquac. Nutr. 2000, 6, 77–84. [Google Scholar] [CrossRef] [Green Version]
- Sørensen, S.L.; Park, Y.; Gong, Y.; Vasanth, G.K.; Dahle, D.; Korsnes, K.; Phuong, T.H.; Kiron, V.; Øyen, S.; Pittman, K.; et al. Nutrient Digestibility, Growth, Mucosal Barrier Status, and Activity of Leucocytes From Head Kidney of Atlantic Salmon Fed Marine- or Plant-Derived Protein and Lipid Sources. Front. Immunol. 2021, 11, 623726. [Google Scholar] [CrossRef]
- Knudsen, D.; Urán, P.; Arnous, A.; Koppe, W.; FRØKIÆR†, H. Saponin-Containing Subfractions of Soybean Molasses Induce Enteritis in the Distal Intestine of Atlantic Salmon. J. Agric. Food Chem. 2007, 55, 2261–2267. [Google Scholar] [CrossRef]
- Nikmaram, N.; Leong, S.Y.; Koubaa, M.; Zhu, Z.; Barba, F.J.; Greiner, R.; Oey, I.; Roohinejad, S. Effect of Extrusion on the Anti-Nutritional Factors of Food Products: An Overview. Food Control. 2017, 79, 62–73. [Google Scholar] [CrossRef]
- Catalán, N.; Villasante, A.; Wacyk, J.; Ramírez, C.; Romero, J. Fermented Soybean Meal Increases Lactic Acid Bacteria in Gut Microbiota of Atlantic Salmon (Salmo salar). Probiotics Antimicrob. Proteins 2018, 10, 566–576. [Google Scholar] [CrossRef]
- Krogdahl, Å.; Kortner, T.M.; Jaramillo-Torres, A.; Gamil, A.A.A.; Chikwati, E.; Li, Y.; Schmidt, M.; Herman, E.; Hymowitz, T.; Teimouri, S.; et al. Removal of Three Proteinaceous Antinutrients from Soybean Does Not Mitigate Soybean-Induced Enteritis in Atlantic Salmon (Salmo salar, L). Aquaculture 2020, 514, 734495. [Google Scholar] [CrossRef]
- Bakke-McKellep, A.M.; Penn, M.H.; Salas, P.M.; Refstie, S.; Sperstad, S.; Landsverk, T.; Ringø, E.; Krogdahl, Å. Effects of Dietary Soyabean Meal, Inulin and Oxytetracycline on Intestinal Microbiota and Epithelial Cell Stress, Apoptosis and Proliferation in the Teleost Atlantic Salmon (Salmo salar L.). Br. J. Nutr. 2007, 97, 699–713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Shemy, H.; Abdel-Rahim, E.; Shaban, O.; Ragab, A.; Carnovale, E.; Fujita, K. Comparison of Nutritional and Antinutritional Factors in Soybean and Fababean Seeds with or without Cortex. Soil Sci. Plant Nutr. 2000, 46, 515–524. [Google Scholar]
- Kamunde, C.; Sappal, R.; Melegy, T.M. Brown Seaweed (AquaArom) Supplementation Increases Food Intake and Improves Growth, Antioxidant Status and Resistance to Temperature Stress in Atlantic Salmon, Salmo salar. PLoS ONE 2019, 14, e0219792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Norambuena, F.; Hermon, K.; Skrzypczyk, V.; Emery, J.A.; Sharon, Y.; Beard, A.; Turchini, G.M. Algae in Fish Feed: Performances and Fatty Acid Metabolism in Juvenile Atlantic Salmon. PLoS ONE 2015, 10, e0124042. [Google Scholar] [CrossRef] [Green Version]
- Johny, A.; Berge, G.M.; Bogevik, A.S.; Krasnov, A.; Ruyter, B.; Fæste, C.K.; Østbye, T.-K.K. Sensitivity to Dietary Wheat Gluten in Atlantic Salmon Indicated by Gene Expression Changes in Liver and Intestine. Genes 2020, 11, 1339. [Google Scholar] [CrossRef] [PubMed]
- Rawles, S.D.; Thompson, K.R.; Brady, Y.J.; Metts, L.S.; Aksoy, M.Y.; Gannam, A.L.; Twibell, R.G.; Ostrand, S.; Webster, C.D. Effects of Replacing Fish Meal with Poultry By-Product Meal and Soybean Meal and Reduced Protein Level on the Performance and Immune Status of Pond-Grown Sunshine Bass (Morone chrysops × M. Saxatilis). Aquac. Nutr. 2011, 17, e708–e721. [Google Scholar] [CrossRef]
- Cammack, J.; Tomberlin, J. The Impact of Diet Protein and Carbohydrate on Select Life-History Traits of The Black Soldier Fly Hermetia Illucens (L.) (Diptera: Stratiomyidae). Insects 2017, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- van Huis, A. Potential of Insects as Food and Feed in Assuring Food Security. Annu. Rev. Entomol. 2013, 58, 563–583. [Google Scholar] [CrossRef]
- Wang, Y.-S.; Shelomi, M. Review of Black Soldier Fly (Hermetia illucens) as Animal Feed and Human Food. Foods 2017, 6, 91. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belghit, I.; Liland, N.S.; Gjesdal, P.; Biancarosa, I.; Menchetti, E.; Li, Y.; Waagbø, R.; Krogdahl, Å.; Lock, E.-J. Black Soldier Fly Larvae Meal Can Replace Fish Meal in Diets of Sea-Water Phase Atlantic Salmon (Salmo salar). Aquaculture 2019, 503, 609–619. [Google Scholar] [CrossRef]
- Li, Y.; Kortner, T.M.; Chikwati, E.M.; Munang’andu, H.M.; Lock, E.-J.; Krogdahl, Å. Gut Health and Vaccination Response in Pre-Smolt Atlantic Salmon (Salmo salar) Fed Black Soldier Fly (Hermetia illucens) Larvae Meal. Fish Shellfish Immunol. 2019, 86, 1106–1113. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Bruni, L.; Jaramillo-Torres, A.; Gajardo, K.; Kortner, T.M.; Krogdahl, Å. Differential Response of Digesta- and Mucosa-Associated Intestinal Microbiota to Dietary Insect Meal during the Seawater Phase of Atlantic Salmon. Anim. Microbiome 2021, 3, 1–18. [Google Scholar] [CrossRef]
- Shafique, L.; Abdel-Latif, H.M.R.; Hassan, F.; Alagawany, M.; Naiel, M.A.E.; Dawood, M.A.O.; Yilmaz, S.; Liu, Q. The Feasibility of Using Yellow Mealworms (Tenebrio molitor): Towards a Sustainable Aquafeed Industry. Animals 2021, 11, 811. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.W.; Karpol, A.; Friedman, S.; Maru, B.T.; Tracy, B.P. Recent Advances in Single Cell Protein Use as a Feed Ingredient in Aquaculture. Curr. Opin. Biotechnol. 2020, 61, 189–197. [Google Scholar] [CrossRef]
- Becker, W. Microalgae in Human and Animal Nutrition. In Handbook of Microalgal Culture; John Wiley & Sons, Ltd.: Hoboken, NJ, USA, 2003; pp. 312–351. ISBN 978-0-470-99528-0. [Google Scholar]
- Sørensen, M.; Gong, Y.; Bjarnason, F.; Vasanth, G.K.; Dahle, D.; Huntley, M.; Kiron, V. Nannochloropsis Oceania-Derived Defatted Meal as an Alternative to Fishmeal in Atlantic Salmon Feeds. PLoS ONE 2017, 12, e0179907. [Google Scholar] [CrossRef] [Green Version]
- Gong, Y.; Sørensen, S.L.; Dahle, D.; Nadanasabesan, N.; Dias, J.; Valente, L.M.P.; Sørensen, M.; Kiron, V. Approaches to Improve Utilization of Nannochloropsis Oceanica in Plant-Based Feeds for Atlantic Salmon. Aquaculture 2020, 522, 735122. [Google Scholar] [CrossRef]
- Grammes, F.; Reveco, F.E.; Romarheim, O.H.; Landsverk, T.; Mydland, L.T.; Øverland, M. Candida Utilis and Chlorella Vulgaris Counteract Intestinal Inflammation in Atlantic Salmon (Salmo salar L.). PLoS ONE 2013, 8, e83213. [Google Scholar] [CrossRef] [Green Version]
- Agboola, J.O.; Øverland, M.; Skrede, A.; Hansen, J.Ø. Yeast as Major Protein-Rich Ingredient in Aquafeeds: A Review of the Implications for Aquaculture Production. Rev. Aquac. 2021, 13, 949–970. [Google Scholar] [CrossRef]
- Øverland, M.; Karlsson, A.; Mydland, L.T.; Romarheim, O.H.; Skrede, A. Evaluation of Candida utilis, Kluyveromyces marxianus and Saccharomyces cerevisiae Yeasts as Protein Sources in Diets for Atlantic Salmon (Salmo salar). Aquaculture 2013, 402–403, 1–7. [Google Scholar] [CrossRef]
- Naylor, R.L. Managing Food Production Systems for Resilience. In Principles of Ecosystem Stewardship: Resilience-Based Natural Resource Management in a Changing World; Folke, C., Kofinas, G.P., Chapin, F.S., Eds.; Springer: New York, NY, USA, 2009; pp. 259–280. ISBN 978-0-387-73033-2. [Google Scholar]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aidos, L.; Mirra, G.; Pallaoro, M.; Herrera Millar, V.R.; Radaelli, G.; Bazzocchi, C.; Modina, S.C.; Di Giancamillo, A. How Do Alternative Protein Resources Affect the Intestine Morphology and Microbiota of Atlantic Salmon? Animals 2023, 13, 1922. https://doi.org/10.3390/ani13121922
Aidos L, Mirra G, Pallaoro M, Herrera Millar VR, Radaelli G, Bazzocchi C, Modina SC, Di Giancamillo A. How Do Alternative Protein Resources Affect the Intestine Morphology and Microbiota of Atlantic Salmon? Animals. 2023; 13(12):1922. https://doi.org/10.3390/ani13121922
Chicago/Turabian StyleAidos, Lucia, Giorgio Mirra, Margherita Pallaoro, Valentina Rafaela Herrera Millar, Giuseppe Radaelli, Chiara Bazzocchi, Silvia Clotilde Modina, and Alessia Di Giancamillo. 2023. "How Do Alternative Protein Resources Affect the Intestine Morphology and Microbiota of Atlantic Salmon?" Animals 13, no. 12: 1922. https://doi.org/10.3390/ani13121922
APA StyleAidos, L., Mirra, G., Pallaoro, M., Herrera Millar, V. R., Radaelli, G., Bazzocchi, C., Modina, S. C., & Di Giancamillo, A. (2023). How Do Alternative Protein Resources Affect the Intestine Morphology and Microbiota of Atlantic Salmon? Animals, 13(12), 1922. https://doi.org/10.3390/ani13121922