
Optimised Stable Lighting Strengthens Circadian Clock Gene Rhythmicity in Equine Hair Follicles


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Experimental Protocol and Lighting
2.3. Real-Time Quantitative Polymerase Chain Reaction (qPCR)
2.4. Data Analysis
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Reppert, S.M.; Weaver, D.R. Coordination of circadian timing in mammals. Nature 2002, 418, 935–941. [Google Scholar] [CrossRef]
- Albrecht, U. Timing to Perfection: The Biology of Central and Peripheral Circadian Clocks. Neuron 2012, 74, 246–260. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bedrosian, T.A.; Nelson, R.J. Timing of light exposure affects mood and brain circuits. Transl. Psychiatry 2017, 7, e1017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowrey, P.L.; Takahashi, J.S. Mammalian Circadian Biology: Elucidating Genome-Wide Levels of Temporal Organization. Annu. Rev. Genom. Hum. Genet. 2004, 5, 407–441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Richards, J.; Gumz, M.L. Advances in understanding the peripheral circadian clocks. FASEB J. 2012, 26, 3602–3613. [Google Scholar] [CrossRef] [Green Version]
- Yamazaki, S.; Numano, R.; Abe, M.; Hida, A.; Takahashi, R.-I.; Ueda, M.; Block, G.D.; Sakaki, Y.; Menaker, M.; Tei, H. Resetting Central and Peripheral Circadian Oscillators in Transgenic Rats. Science 2000, 288, 682–685. [Google Scholar] [CrossRef] [Green Version]
- Geyfman, M.; Andersen, B. How the Skin Can Tell Time. J. Invest. Dermatol. 2009, 129, 1063–1066. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0022202X15343414 (accessed on 6 May 2023). [CrossRef] [Green Version]
- Yoo, S.-H.; Yamazaki, S.; Lowrey, P.L.; Shimomura, K.; Ko, C.H.; Buhr, E.D.; Siepka, S.M.; Hong, H.-K.; Oh, W.J.; Yoo, O.J.; et al. PERIOD2::LUCIFERASE real-time reporting of circadian dynamics reveals persistent circadian oscillations in mouse peripheral tissues. Proc. Natl. Acad. Sci. USA 2004, 101, 5339–5346. [Google Scholar] [CrossRef]
- Murphy, B.A.; Vick, M.M.; Sessions, D.R.; Cook, R.F.; Fitzgerald, B.P. Evidence of an oscillating peripheral clock in an equine fibroblast cell line and adipose tissue but not in peripheral blood. J. Comp. Physiol. A 2006, 192, 743–751. [Google Scholar] [CrossRef]
- Martin, A.-M.; Elliott, J.A.; Duffy, P.; Blake, C.M.; Ben Attia, S.; Katz, L.M.; Browne, J.A.; Gath, V.; McGivney, B.A.; Hill, E.W.; et al. Circadian regulation of locomotor activity and skeletal muscle gene expression in the horse. J. Appl. Physiol. 2010, 109, 1328–1336. [Google Scholar] [CrossRef] [Green Version]
- Watts, L.M.; A Browne, J.; A Murphy, B. Investigation of a non-invasive method of assessing the equine circadian clock using hair follicle cells. J. Circadian Rhythm. 2012, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Giannetto, C.; Cannella, V.; Giudice, E.; Guercio, A.; Piccione, G. Behavioral and physiological processes in horses and their linkage with peripheral clock gene expression: A preliminary study. J. Vet. Behav. 2019, 34, 37–41. [Google Scholar] [CrossRef]
- Giannetto, C.; Fazio, F.; Alberghina, D.; Giudice, E.; Piccione, G. Clock Genes Expression in Peripheral Leukocytes and Plasma Melatonin Daily Rhythm in Horses. J. Equine Vet. Sci. 2020, 84, 102856. Available online: https://linkinghub.elsevier.com/retrieve/pii/S0737080619306057 (accessed on 13 July 2023). [CrossRef]
- Giannetto, C.; Fazio, F.; Giudice, E.; Mazzullo, G.; Piccione, G. Physiological role of circadian clock gene on the energetic me-tabolism in horses. J. Vet. Behav. 2022, 47, 29–34. [Google Scholar] [CrossRef]
- Brainard, G.C.; Sliney, D.; Hanifin, J.P.; Glickman, G.; Byrne, B.; Greeson, J.M.; Jasser, S.; Gerner, E.; Rollag, M.D. Sensitivity of the Human Circadian System to Short-Wavelength (420-nm) Light. J. Biol. Rhythm. 2008, 23, 379–386. [Google Scholar] [CrossRef] [Green Version]
- Berson, D.M.; Dunn, F.A.; Takao, M. Phototransduction by Retinal Ganglion Cells That Set the Circadian Clock. Science 2002, 295, 1070–1073. [Google Scholar] [CrossRef] [Green Version]
- Hattar, S.; Liao, H.-W.; Takao, M.; Berson, D.M.; Yau, K.-W. Melanopsin-containing retinal ganglion cells: Architecture, projections, and intrinsic photosensitivity. Science 2002, 295, 1065–1070. [Google Scholar] [CrossRef] [Green Version]
- Brainard, G.C.; Hanifin, J.P.; Greeson, J.; Byrne, B.; Glickman, G.; Gerner, E.; Rollag, M.D. Action Spectrum for Melatonin Regulation in Humans: Evidence for a Novel Circadian Photoreceptor. J. Neurosci. 2001, 21, 6405–6412. [Google Scholar] [CrossRef] [Green Version]
- Pevet, P.; Challet, E. Melatonin: Both master clock output and internal time-giver in the circadian clocks network. J. Physiol. 2011, 105, 170–182. [Google Scholar] [CrossRef]
- Walsh, C.; Prendergast, R.; Sheridan, J.; Murphy, B. Blue light from light-emitting diodes directed at a single eye elicits a dose-dependent suppression of melatonin in horses. Vet. J. 2013, 196, 231–235. [Google Scholar] [CrossRef]
- Murphy, B.A.; Walsh, C.M.; Woodward, E.M.; Prendergast, R.L.; Ryle, J.P.; Fallon, L.H.; Troedsson, M.H.T. Blue light from individual light masks directed at a single eye advances the breeding season in mares. Equine Vet. J. 2013, 46, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Bedrosian, T.A.; Vaughn, C.A.; Galan, A.; Daye, G.; Weil, Z.; Nelson, R.J. Nocturnal Light Exposure Impairs Affective Responses in a Wavelength-Dependent Manner. J. Neurosci. 2013, 33, 13081–13087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lockley, S.W.; Gooley, J.J. Circadian Photoreception: Spotlight on the Brain. Curr. Biol. 2006, 16, R795–R797. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stenvers, D.J.; van Dorp, R.; Foppen, E.; Mendoza, J.; Opperhuizen, A.-L.; Fliers, E.; Bisschop, P.H.; Meijer, J.H.; Kalsbeek, A.; Deboer, T. Dim light at night disturbs the daily sleep-wake cycle in the rat. Sci. Rep. 2016, 6, 35662. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Z.; Wang, H.-J.; Wang, D.-R.; Qu, W.-M.; Huang, Z.-L. Red light at intensities above 10 lx alters sleep–wake behavior in mice. Light. Sci. Appl. 2017, 6, e16231. [Google Scholar] [CrossRef] [Green Version]
- Opperhuizen, A.-L.; Stenvers, D.J.; Jansen, R.D.; Foppen, E.; Fliers, E.; Kalsbeek, A. Light at night acutely impairs glucose tolerance in a time-, intensity- and wavelength-dependent manner in rats. Diabetologia 2017, 60, 1333–1343. [Google Scholar] [CrossRef] [Green Version]
- Higuchi, S.; Fukuda, T.; Kozaki, T.; Takahashi, M.; Miura, N. Effectiveness of a Red-visor Cap for Preventing Light-induced Melatonin Suppression during Simulated Night Work. J. Physiol. Anthr. 2011, 30, 251–258. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.; O’Brien, C.; Elliott, J. Red light at night permits the nocturnal rise of melatonin production in horses. Vet. J. 2019, 252, 105360. [Google Scholar] [CrossRef]
- Sakamoto, K.; Nagase, T.; Fukui, H.; Horikawa, K.; Okada, T.; Tanaka, H.; Sato, K.; Miyake, Y.; Ohara, O.; Kako, K.; et al. Multitissue Circadian Expression of Rat periodHomolog (rPer2) mRNA Is Governed by the Mammalian Circadian Clock, the Suprachiasmatic Nucleus in the Brain. J. Biol. Chem. 1998, 273, 27039–27042. [Google Scholar] [CrossRef] [Green Version]
- Fonken, L.K.; Aubrecht, T.G.; Meléndez-Fernández, O.H.; Weil, Z.M.; Nelson, R.J. Dim light at night disrupts molecular circadian rhythms and increases body weight. J. Biol. Rhythm. 2013, 28, 262–271. [Google Scholar] [CrossRef]
- Polidarová, L.; Sládek, M.; Soták, M.; Pácha, J.; Sumová, A. Hepatic, Duodenal, and Colonic Circadian Clocks Differ in their Persistence under Conditions of Constant Light and in their Entrainment by Restricted Feeding. Chrono-Int. 2011, 28, 204–215. [Google Scholar] [CrossRef]
- Benloucif, S.; Guico, M.J.; Reid, K.J.; Wolfe, L.F.; L’hermite-Balériaux, M.; Zee, P.C. Stability of Melatonin and Temperature as Circadian Phase Markers and Their Relation to Sleep Times in Humans. J. Biol. Rhythm. 2005, 20, 178–188. [Google Scholar] [CrossRef] [Green Version]
- Refinetti, R.; Menaker, M. The circadian rhythm of body temperature. Physiol. Behav. 1992, 51, 613–637. [Google Scholar] [CrossRef]
- Murphy, B.A.; Elliott, J.A.; Sessions, D.R.; Vick, M.M.; Kennedy, E.L.; Fitzgerald, B.P. Rapid phase adjustment of melatonin and core body temperature rhythms following a 6-h advance of the light/dark cycle in the horse. J. Circadian Rhythm. 2007, 5, 5. [Google Scholar] [CrossRef] [Green Version]
- Boivin, D.B.; James, F.O.; Wu, A.; Cho-Park, P.F.; Xiong, H.; Sun, Z.S. Circadian clock genes oscillate in human peripheral blood mononuclear cells. Blood 2003, 102, 4143–4145. Available online: https://ashpublications.org/blood/article/102/12/4143/16705/Circadian-clock-genes-oscillate-in-human (accessed on 6 May 2023). [CrossRef] [Green Version]
- Burgess, H.J.; Wyatt, J.K.; Park, M.; Fogg, L.F. Home Circadian Phase Assessments with Measures of Compliance Yield Accurate Dim Light Melatonin Onsets. Sleep 2015, 38, 889–897. [Google Scholar] [CrossRef] [Green Version]
- Akashi, M.; Soma, H.; Yamamoto, T.; Tsugitomi, A.; Yamashita, S.; Yamamoto, T.; Nishida, E.; Yasuda, A.; Liao, J.K.; Node, K. Noninvasive method for assessing the human circadian clock using hair follicle cells. Proc. Natl. Acad. Sci. USA 2010, 107, 15643–15648. [Google Scholar] [CrossRef]
- Liu, L.-P.; Li, M.-H.; Zheng, Y.-W. Hair Follicles as a Critical Model for Monitoring the Circadian Clock. Int. J. Mol. Sci. 2023, 24, 2407. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Mitsui, S.; Yan, L.; Yagita, K.; Miyake, S.; Okamura, H. Role of DBP in the Circadian Oscillatory Mechanism. Mol. Cell. Biol. 2000, 20, 4773–4781. [Google Scholar] [CrossRef] [Green Version]
- Yoshitane, H.; Asano, Y.; Sagami, A.; Sakai, S.; Suzuki, Y.; Okamura, H.; Iwasaki, W.; Ozaki, H.; Fukada, Y. Functional D-box sequences reset the circadian clock and drive mRNA rhythms. Commun. Biol. 2019, 2, 300. [Google Scholar] [CrossRef] [Green Version]
- Refinetti, R.; Cornélissen, G.; Halberg, F. Procedures for numerical analysis of circadian rhythms. Biol. Rhythm. Res. 2007, 38, 275–325. [Google Scholar] [CrossRef] [PubMed]
- Halberg, F.; Tong, Y.L.; Johnson, E.A. Circadian System Phase—An Aspect of Temporal Morphology; Procedures and Illustrative Examples; The Cellular Aspects of Biorhythms; Springer: Berlin/Heidelberg, Germany, 1967; pp. 20–48. [Google Scholar] [CrossRef]
- Bartness, T.J.; Song, C.K.; Demas, G.E. SCN Efferents to Peripheral Tissues: Implications for Biological Rhythms. J. Biol. Rhythm. 2001, 16, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Brown, S.A.; Zumbrunn, G.; Fleury-Olela, F.; Preitner, N.; Schibler, U. Rhythms of Mammalian Body Temperature Can Sustain Peripheral Circadian Clocks. Curr. Biol. 2002, 12, 1574–1583. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiola, F.; Minh, N.L.; Preitner, N.; Kornmann, B.; Fleury-Olela, F.; Schibler, U. Restricted feeding uncouples circadian oscillators in peripheral tissues from the central pacemaker in the suprachiasmatic nucleus. Genes Dev. 2000, 14, 2950–2961. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- A Ripperger, J.; Shearman, L.P.; Reppert, S.M.; Schibler, U. CLOCK, an essential pacemaker component, controls expression of the circadian transcription factor DBP. Genes Dev. 2000, 14, 679–689. Available online: http://www.ncbi.nlm.nih.gov/pubmed/10733528 (accessed on 6 May 2023). [CrossRef]
- Bozek, K.; Relógio, A.; Kielbasa, S.M.; Heine, M.; Dame, C.; Kramer, A.; Herzel, H. Regulation of Clock-Controlled Genes in Mammals. PLoS ONE 2009, 4, e4882. [Google Scholar] [CrossRef] [Green Version]
- Storch, K.-F.; Lipan, O.; Leykin, I.; Viswanathan, N.; Davis, F.C.; Wong, W.H.; Weitz, C.J. Extensive and divergent circadian gene expression in liver and heart. Nature 2002, 417, 78–83. [Google Scholar] [CrossRef]
- Welsh, D.K.; Logothetis, D.E.; Meister, M.; Reppert, S.M. Individual neurons dissociated from rat suprachiasmatic nucleus express independently phased circadian firing rhythms. Neuron 1995, 14, 697–706. [Google Scholar] [CrossRef] [Green Version]
- Yamaguchi, S.; Isejima, H.; Matsuo, T.; Okura, R.; Yagita, K.; Kobayashi, M.; Okamura, H. Synchronization of Cellular Clocks in the Suprachiasmatic Nucleus. Science 2003, 302, 1408–1412. [Google Scholar] [CrossRef]
- Welsh, D.K.; Yoo, S.-H.; Liu, A.C.; Takahashi, J.S.; Kay, S.A. Bioluminescence Imaging of Individual Fibroblasts Reveals Persis-tent, Independently Phased Circadian Rhythms of Clock Gene Expression. Curr. Biol. 2004, 14, 2289–2295. Available online: http://linkinghub.elsevier.com/retrieve/pii/S0960982204009157 (accessed on 6 May 2023). [CrossRef] [Green Version]
- Zambon, A.C.; McDearmon, E.L.; Salomonis, N.; Vranizan, K.M.; Johansen, K.L.; Adey, D.; Takahashi, J.S.; Schambelan, M.; Conklen, B.R. Time- and exercise-dependent gene regulation in human skeletal muscle. Genome Biol. 2003, 4, R61. Available online: http://www.ncbi.nlm.nih.gov/pubmed/14519196 (accessed on 6 May 2023). [CrossRef] [Green Version]
- McCarthy, J.J.; Andrews, J.L.; McDearmon, E.L.; Campbell, K.S.; Barber, B.K.; Miller, B.H.; Walker, J.R.; Hogenesch, J.B.; Takahashi, J.S.; Esser, K.A.; et al. Identification of the circadian transcriptome in adult mouse skeletal muscle. Physiol. Genom. 2007, 31, 86–95. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.; Wagner, A.; McGlynn, O.; Kharazyan, F.; Browne, J.; Elliott, J. Exercise influences circadian gene expression in equine skeletal muscle. Vet. J. 2014, 201, 39–45. [Google Scholar] [CrossRef]
- Panda, S. Circadian physiology of metabolism. Science 2016, 354, 1008–1015. [Google Scholar] [CrossRef]
- Schibler, U.; Gotic, I.; Saini, C.; Gos, P.; Curie, T.; Emmenegger, Y.; Sinturel, F.; Gosselin, P.; Gerber, A.; Fleury-Olela, F.; et al. Clock-Talk: Interactions between Central and Peripheral Circadian Oscillators in Mammals. In Cold Spring Harbor Symposia on Quantitative Biology; Cold Spring Harbor Laboratory Press: New York, NY, USA, 2015; Volume 80, pp. 223–232. [Google Scholar] [CrossRef] [Green Version]
- Kofuji, P.; Mure, L.S.; Massman, L.J.; Purrier, N.; Panda, S.; Engeland, W.C. Intrinsically Photosensitive Retinal Ganglion Cells (ipRGCs) Are Necessary for Light Entrainment of Peripheral Clocks. PLoS ONE 2016, 11, e0168651. [Google Scholar] [CrossRef] [Green Version]
- Fan, S.M.-Y.; Chang, Y.-T.; Chen, C.-L.; Wang, W.-H.; Pan, M.-K.; Chen, W.-P.; Huang, W.-Y.; Xu, Z.; Huang, H.-E.; Chen, T.; et al. External light activates hair follicle stem cells through eyes via an ipRGC–SCN–sympathetic neural pathway. Proc. Natl. Acad. Sci. USA 2018, 115, E6880–E6889. [Google Scholar] [CrossRef] [Green Version]
- Hanifin, J.; Lockley, S.; Cecil, K.; West, K.; Jablonski, M.; Warfield, B.; James, M.; Ayers, M.; Byrne, B.; Gerner, E.; et al. Randomized trial of polychromatic blue-enriched light for circadian phase shifting, melatonin suppression, and alerting responses. Physiol. Behav. 2018, 198, 57–66. [Google Scholar] [CrossRef] [Green Version]
- Najjar, R.P.; Wolf, L.; Taillard, J.; Schlangen, L.J.M.; Salam, A.; Cajochen, C.; Gronfier, C. Chronic Artificial Blue-Enriched White Light Is an Effective Countermeasure to Delayed Circadian Phase and Neurobehavioral Decrements. PLoS ONE 2014, 9, e102827. [Google Scholar] [CrossRef]
- MacIsaac, D.; Kanner, G.; Anderson, G. Basic physics of the incandescent lamp (lightbulb). Phys. Teach. 1999, 37, 520–525. [Google Scholar] [CrossRef] [Green Version]
- Paus, R.; Cotsarelis, G. The Biology of Hair Follicles. N. Engl. J. Med. 1999, 341, 491–497. [Google Scholar] [CrossRef] [Green Version]
- Lin, K.K.; Kumar, V.; Geyfman, M.; Chudova, D.; Ihler, A.T.; Smyth, P.; Paus, R.; Takahashi, J.S.; Andersen, B. Circadian Clock Genes Contribute to the Regulation of Hair Follicle Cycling. PLoS Genet. 2009, 5, e1000573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Brien, C.; Darcy-Dunne, M.R.; Murphy, B.A. The effects of extended photoperiod and warmth on hair growth in ponies and horses at different times of year. PLoS ONE 2020, 15, e0227115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buhr, E.D.; Takahashi, J.S. Molecular Components of the Mammalian Circadian Clock. Handb. Exp. Pharmacol. 2013, 217, 3–27. [Google Scholar] [CrossRef] [Green Version]
- Bonmati-Carrion, M.A.; Arguelles-Prieto, R.; Martinez-Madrid, M.J.; Reiter, R.; Hardeland, R.; Rol, M.A.; Madrid, J.A. Protecting the Melatonin Rhythm through Circadian Healthy Light Exposure. Int. J. Mol. Sci. 2014, 15, 23448–23500. [Google Scholar] [CrossRef] [Green Version]
- Pauley, S.M. Lighting for the human circadian clock: Recent research indicates that lighting has become a public health issue. Med. Hypotheses 2004, 63, 588–596. [Google Scholar] [CrossRef] [Green Version]
- Murphy, B.A. Circadian and Circannual Regulation in the Horse: Internal Timing in an Elite Athlete. J. Equine Vet. Sci. 2019, 76, 14–24. [Google Scholar] [CrossRef]




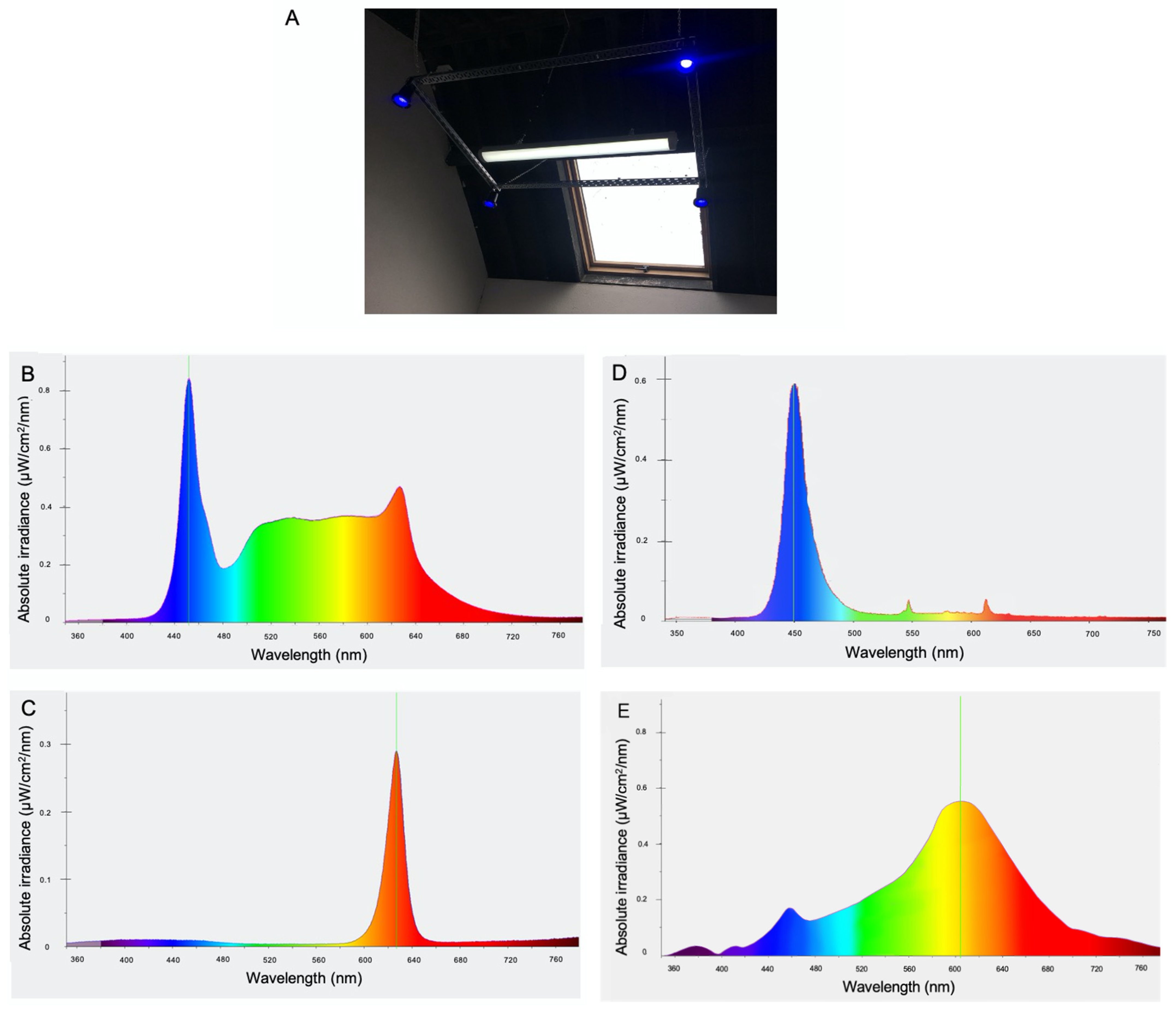{kind=link}
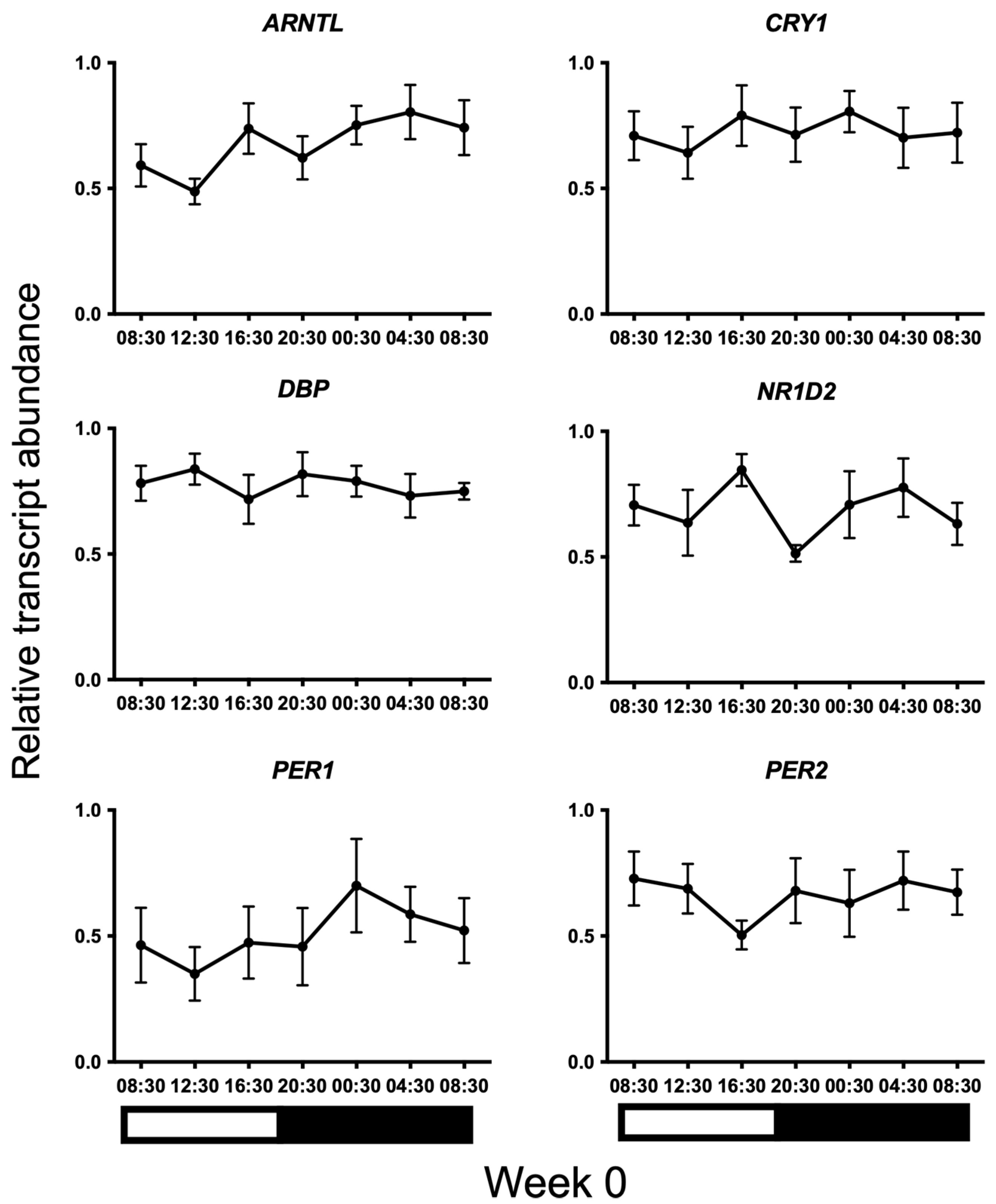{kind=link}
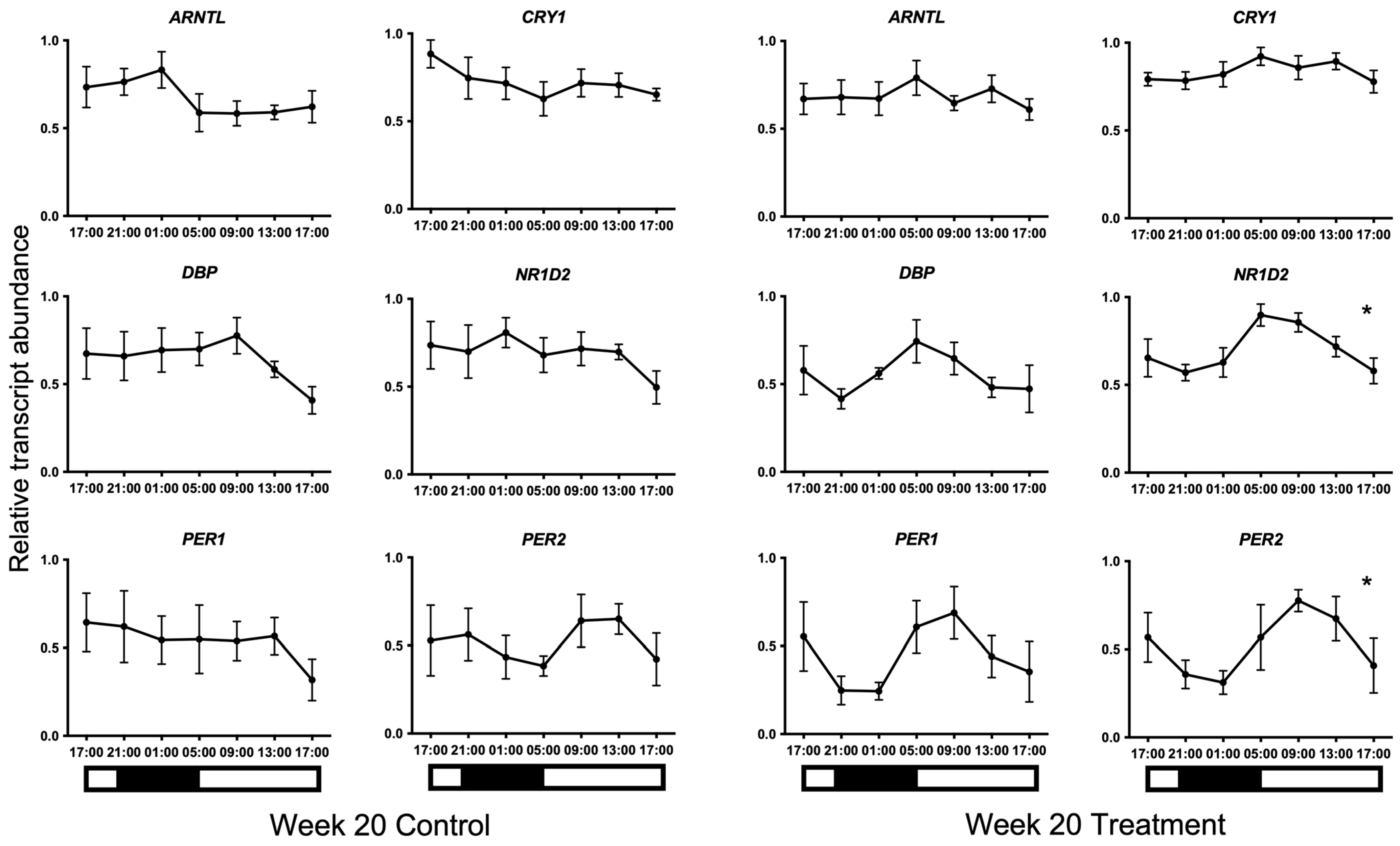{kind=link}
| Gene Symbol | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| ACTB | CACCTTCTACAACGAGCTGC | CGGGGTGTTGAAGGTCTCA |
| GAPDH | GGAGTCCACTGGTGTCTTCA | GTTCACGCCCATCACAAACA |
| H3F3A | CAAACTTCCCTTCCAGCGTC | TGGATAGCACACAGGTTGGT |
| PPIA | GCATCTTGTCCATGGCGAAT | CAAAGACCACATGCTTGCCA |
| RPL19 | CTGATCATCCGGAAGCCTGT | GGCAGTACCCTTTCGCTTAC |
| SDHA | TGTTGTGTCTCGGTCCATGA | AGATCATGGCCGTCTCTGAA |
| YWHAZ | AGACGGAAGGTGCTGAGAAA | CTTGTGAAGCATTGGGGATCA |
| RNF11 | AGGATAGCTCAAAGAATAGGC | CGGCAGAAATCGAATTGGGT |
| Gene Symbol | Forward Primer Sequence (5′-3′) | Reverse Primer Sequence (5′-3′) |
|---|---|---|
| ARNTL | CACCTTCTACAACGAGCTGC | CGGGGTGTTGAAGGTCTCA |
| CRY1 | GGAGTCCACTGGTGTCTTCA | GTTCACGCCCATCACAAACA |
| DBP | CAAACTTCCCTTCCAGCGTC | TGGATAGCACACAGGTTGGT |
| NR1D2 | GCATCTTGTCCATGGCGAAT | CAAAGACCACATGCTTGCCA |
| PER1 | CTGATCATCCGGAAGCCTGT | GGCAGTACCCTTTCGCTTAC |
| PER2 | TGTTGTGTCTCGGTCCATGA | AGATCATGGCCGTCTCTGAA |
| Week 0 | Week 20 (Control) | Week 20 (Treatment) | |||||||
|---|---|---|---|---|---|---|---|---|---|
| Gene Transcript | Robustness (%) | Acrophase (24 h) | p Value | Robustness (%) | Acrophase (24 h) | p Value | Robustness (%) | Acrophase (24 h) | p Value |
| ARNTL | 36.0 | n/a | 0.411 | 74.1 | 22:31 | 0.068 | 26.0 | n/a | 0.550 |
| CRY1 | 36.9 | n/a | 0.399 | 24.2 | n/a | 0.577 | 77.8 | 07:45 | 0.051 |
| DBP | 6.0 | n/a | 0.882 | 44.0 | n/a | 0.313 | 63.5 | n/a | 0.134 |
| NR1D2 | 1.6 | n/a | 0.970 | 2.5 | n/a | 0.603 | 89.5 * | 07:47 | 0.013 |
| PER1 | 68.2 | n/a | 0.101 | 2.6 | n/a | 0.949 | 65.3 | n/a | 0.121 |
| PER2 | 48.2 | n/a | 0.268 | 37.6 | n/a | 0.391 | 89.6 * | 10:23 | 0.013 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Collery, A.; Browne, J.A.; O’Brien, C.; Sheridan, J.T.; Murphy, B.A. Optimised Stable Lighting Strengthens Circadian Clock Gene Rhythmicity in Equine Hair Follicles. Animals 2023, 13, 2335. https://doi.org/10.3390/ani13142335
Collery A, Browne JA, O’Brien C, Sheridan JT, Murphy BA. Optimised Stable Lighting Strengthens Circadian Clock Gene Rhythmicity in Equine Hair Follicles. Animals. 2023; 13(14):2335. https://doi.org/10.3390/ani13142335
Chicago/Turabian StyleCollery, Aileen, John A. Browne, Christiane O’Brien, John T. Sheridan, and Barbara A. Murphy. 2023. "Optimised Stable Lighting Strengthens Circadian Clock Gene Rhythmicity in Equine Hair Follicles" Animals 13, no. 14: 2335. https://doi.org/10.3390/ani13142335
APA StyleCollery, A., Browne, J. A., O’Brien, C., Sheridan, J. T., & Murphy, B. A. (2023). Optimised Stable Lighting Strengthens Circadian Clock Gene Rhythmicity in Equine Hair Follicles. Animals, 13(14), 2335. https://doi.org/10.3390/ani13142335