
A Case of Bovine Eosinophilic Myositis (BEM) Associated with Co-Infection by Sarcocystis hominis and Toxoplasma gondii

 , , , , and
, , , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Source Material
2.2. Gross Pathology, Cytology and Histology Investigation
2.3. Meat Juice Extraction and Serology
2.4. Molecular Investigations

{kind=link}
{kind=link}
{kind=link}
| Primers | Gene | Primer Sequences | Product Length | Reference | |
|---|---|---|---|---|---|
| 18S Apicomplexa | COC-1 | 18S | AAGTATAAGCTTTTATACGGCT | 300 bp | [31] |
| COC-2 | CACTGCCACGGTAGTCCAATA | ||||
| Sarcocystis spp. Multiplex PCR | Sarco_Rev | 18S | AACCCTAATTCCCCGTTA | [15] | |
| SarF | TGGCTAATACATGCGCAAATA | 200–250 bp | [35] | ||
| Hirsuta | CATTTCGGTGATTATTGG | 108 bp | [15] | ||
| Cruzi | ATCAGATGAAAATCTACTACATGG | 300 bp | |||
| COI_HB | COI | AATGTGGTGCGGTATGAACT | [19] | ||
| COI_H | GGCACCAACGAACATGGTA | 420 bp | |||
| COI_B | TCAAAAACCTGCTTTGCTG | 700 bp | |||
| B1 Toxoplasma nested PCR | I Round for | B1 | GGAACTGCATCCGTTCATGAG | [32] | |
| I Round rev | TCTTTAAAGCGTTCGTGGTC- | ||||
| II Round for | TGCATAGGTTGCAGTCACTG | 96bp | |||
| II Round rev | GGCGACCAATCTGCGAATACACC |
3. Results
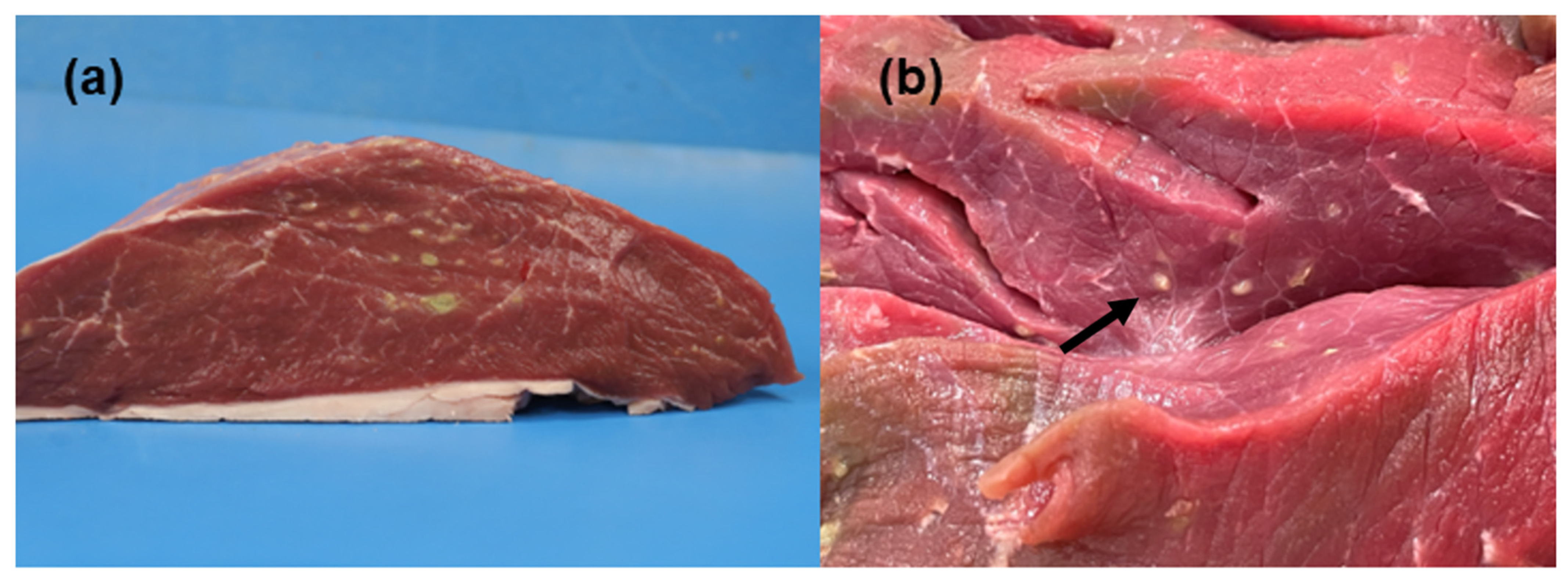
3.1. Gross-Pathological Findings
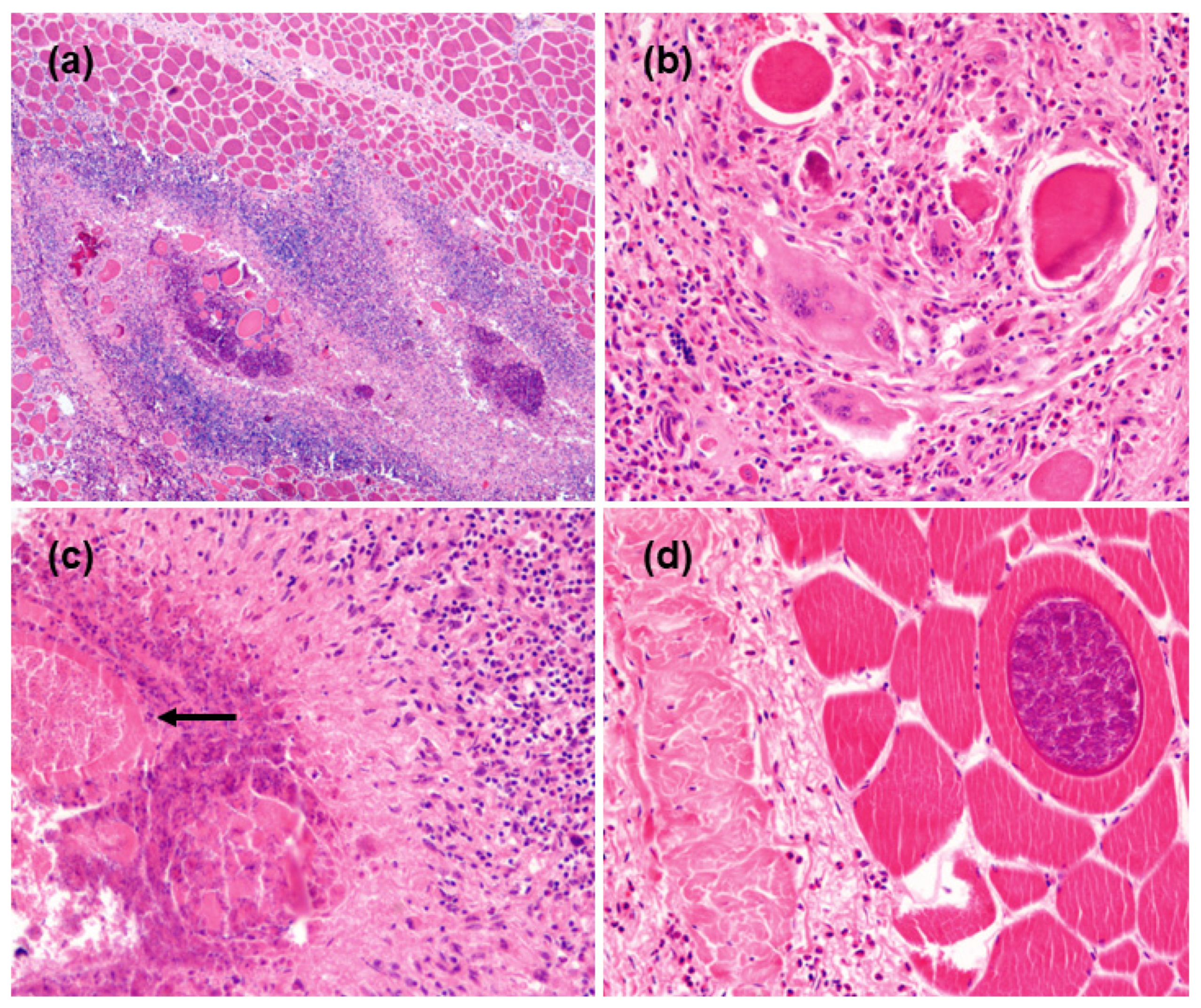
3.2. Cytological and Histological Findings
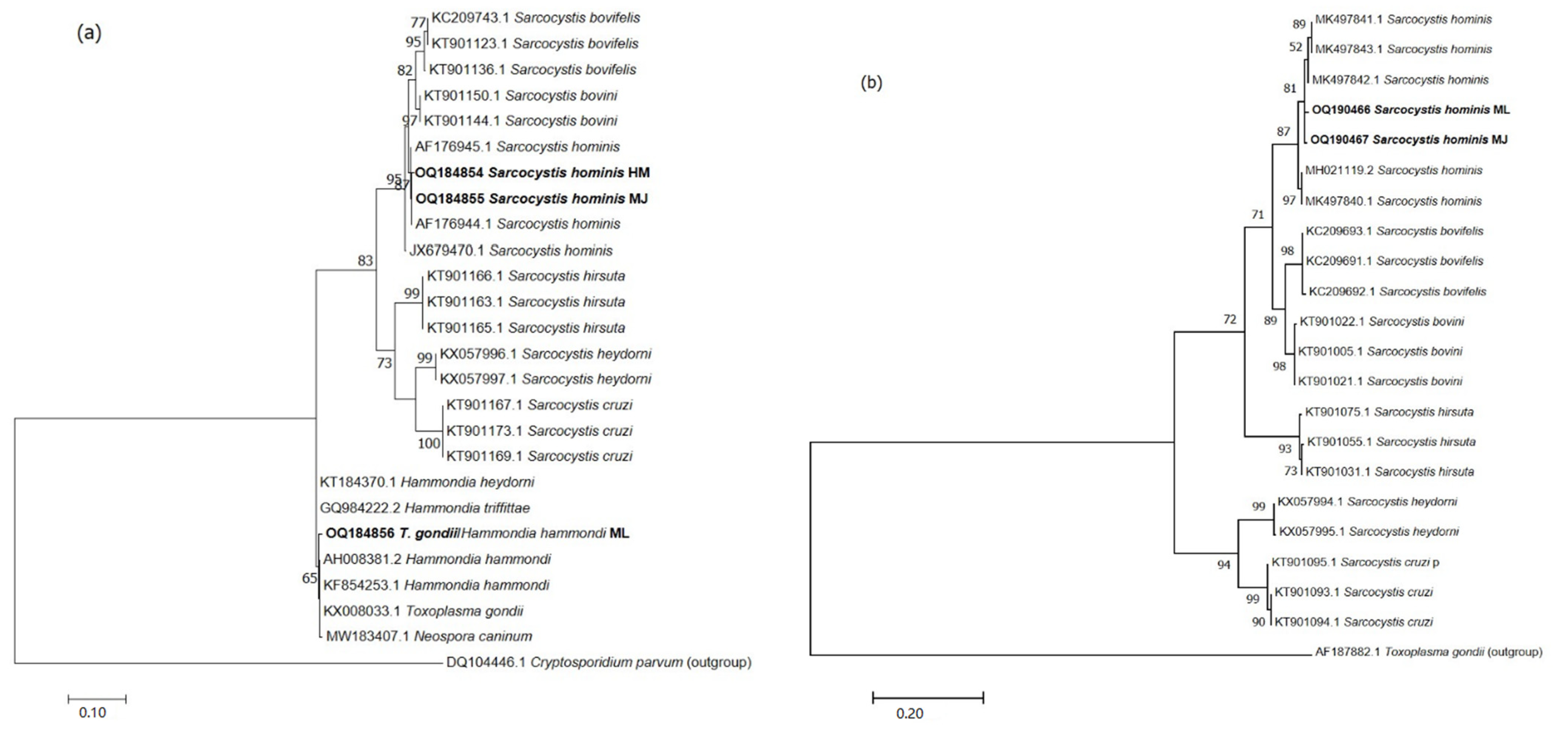
3.3. Serological and Molecular Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rubiola, S.; Civera, T.; Panebianco, F.; Vercellino, D.; Chiesa, F. Molecular Detection of Cattle Sarcocystis spp. in North-West Italy highlights their association with bovine eosinophilic myositis. Parasites Vectors 2021, 14, 1–8. [Google Scholar] [CrossRef]
- Gjerde, B. Molecular Characterisation of Sarcocystis bovifelis, Sarcocystis bovini sp., Sarcocystis hirsuta and Sarcocystis cruzi from Cattle (Bos taurus) and Sarcocystis sinensis from Water Buffaloes (Bubalus bubalis). Parasitol. Res. 2016, 115, 1473–1492. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P.; van Wilpe, E.; Calero-Bernal, R.; Verma, S.K.; Fayer, R. Sarcocystis heydorni, n. sp. (Apicomplexa: Sarcocystidae) with cattle (Bos taurus) and human (Homo sapiens) cycle. Parasitol. Res. 2015, 114, 4143–4147. [Google Scholar] [CrossRef] [PubMed]
- Wouda, W.; Snoep, J.J.; Dubey, J.P. Eosinophilic myositis due to Sarcocystis hominis in a beef cow. J. Comp. Pathol. 2006, 135, 249–253. [Google Scholar] [CrossRef] [PubMed]
- Vangeel, L.; Houf, K.; Geldhof, P.; De Preter, K.; Vercruysse, J.; Ducatelle, R.; Chiers, K. Different Sarcocystis spp. Are present in bovine eosinophilic myositis. Vet. Parasitol. 2013, 197, 543–548. [Google Scholar] [CrossRef]
- Hoeve-Bakker, B.J.A.; van der Giessen, J.W.B.; Franssen, F.F.J. Molecular identification targeting Cox1 and 18s genes confirms the high prevalence of Sarcocystis spp. in cattle in the Netherlands. Int. J. Parasitol. 2019, 49, 859–866. [Google Scholar] [CrossRef]
- Gajadhar, A.A.; Marquardt, W.C. Ultrastructural and transmission evidence of Sarcocysis cruzi associated with eosinophilic myositis in cattle. Can. J. Vet. Res. 1992, 56, 41. [Google Scholar] [PubMed]
- Waine, K.; Bartley, P.M.; Cox, A.; Newsome, R.; Strugnell, B.; Katzer, F. Molecular detection of Sarcocystis cruzi in three beef carcases with eosinophilic myositis lesions and in unaffected beef from animals in the same herd. Vet. Parasitol. Reg. Stud. Rep. 2022, 33, 100751. [Google Scholar] [CrossRef]
- Dubey, J.P.; Rosenthal, B.M. Bovine sarcocystosis: Sarcocystis species, diagnosis, prevalence, economic and public health considerations, and association of Sarcocystis species with eosinophilic myositis in cattle. Int. J. Parasitol. 2022; in press. [Google Scholar] [CrossRef]
- Lindsay, D.S.; Dubey, J.P. Neosporosis, Toxoplasmosis, and Sarcocystosis in ruminants: An update. Vet. Clin. N. Am. Food Anim. Pract. 2020, 36, 205–222. [Google Scholar] [CrossRef]
- Dubey, J.P.; Calero-Bernal, R.; Rosenthal, B.M.; Speer, C.A.; Fayer, R. Sarcocystosis of Animals and Humans, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015; ISBN 9781498710152. [Google Scholar]
- Vangeel, L.; Houf, K.; Geldhof, P.; Nollet, H.; Vercruysse, J.; Ducatelle, R.; Chiers, K. Intramuscular inoculation of cattle with Sarcocystis antigen results in focal eosinophilic myositis. Vet. Parasitol. 2012, 183, 224–230. [Google Scholar] [CrossRef]
- Bucca, M.; Brianti, E.; Giuffrida, A.; Ziino, G.; Cicciari, S.; Panebianco, A. Prevalence and distribution of Sarcocystis spp. cysts in several muscles of cattle slaughtered in Sicily, Southern Italy. Food Control 2011, 22, 105–108. [Google Scholar] [CrossRef]
- Domenis, L.; Peletto, S.; Sacchi, L.; Clementi, E.; Genchi, M.; Felisari, L.; Felisari, C.; Mo, P.; Modesto, P.; Zuccon, F.; et al. Detection of a morphogenetically novel Sarcocystis hominis-like in the context of a prevalence study in semi-intensively bred cattle in Italy. Parasitol. Res. 2011, 109, 1677–1687. [Google Scholar] [CrossRef] [PubMed]
- Chiesa, F.; Muratore, E.; Dalmasso, A.; Civera, T. A New Molecular approach to assess the occurrence of Sarcocystis spp. in cattle and products thereof: Preliminary data. Ital. J. Food Saf. 2013, 2, 148–151. [Google Scholar] [CrossRef]
- Meistro, S.; Peletto, S.; Pezzolato, M.; Varello, K.; Botta, M.; Richelmi, G.; Biglia, C.; Baioni, E.; Modesto, P.; Acutis, P.; et al. Sarcocystis spp. prevalence in bovine minced meat: A histological and molecular study. Ital. J. Food Saf. 2015, 4, 85–87. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubey, J.P. Foodborne and waterborne zoonotic sarcocystosis. Food Waterborne Parasitol. 2015, 1, 2–11. [Google Scholar] [CrossRef] [Green Version]
- Moré, G.; Schares, S.; Maksimov, A.; Conraths, F.J.; Venturini, M.C.; Schares, G. Development of a multiplex real time PCR to differentiate Sarcocystis spp. affecting cattle. Vet. Parasitol. 2013, 197, 85–94. [Google Scholar] [CrossRef]
- Rubiola, S.; Civera, T.; Ferroglio, E.; Zanet, S.; Zaccaria, T.; Brossa, S.; Cipriani, R.; Chiesa, F. Molecular Differentiation of Cattle Sarcocystis spp. by Multiplex PCR Targeting 18S and COI Genes Following Identification of Sarcocystis hominis in Human Stool Samples. Food Waterborne Parasitol. 2020, 18, e00074. [Google Scholar] [CrossRef]
- Prakas, P.; Strazdaitė-Žielienė, Ž.; Januškevičius, V.; Chiesa, F.; Baranauskaitė, A.; Rudaitytė-Lukošienė, E.; Servienė, E.; Petkevičius, S.; Butkauskas, D. Molecular identification of four Sarcocystis species in cattle from Lithuania, including S. hominis, and development of a rapid molecular detection method. Parasit. Vectors. 2020, 13, 610. [Google Scholar] [CrossRef]
- Gjerde, B. Phylogenetic Relationships among Sarcocystis Species in Cervids, Cattle and Sheep inferred from the mitochondrial cytochrome c oxidase subunit i gene. Int. J. Parasitol. 2013, 43, 579–591. [Google Scholar] [CrossRef]
- Zeng, H.; Van Damme, I.; Kabi, T.W.; Šoba, B.; Gabriël, S. Sarcocystis species in bovine carcasses from a Belgian abattoir: A cross-sectional study. Parasit. Vectors. 2021, 14, 271. [Google Scholar] [CrossRef]
- Rubiola, S.; Chiesa, F.; Zanet, S.; Civera, T. Molecular Identification of Sarcocystis spp. in Cattle: Partial Sequencing of Cytochrome C Oxidase Subunit 1 (COI). Ital. J. Food Saf. 2018, 7, 204–208. [Google Scholar] [CrossRef]
- Moré, G.; Basso, W.; Bacigalupe, D.; Venturini, M.C.; Venturini, L. Diagnosis of Sarcocystis cruzi, Neospora caninum, and Toxoplasma gondii infections in cattle. Parasitol. Res. 2008, 102, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Amairia, S.; Rouatbi, M.; Rjeibi, M.R.; Gomes, J.; Rekik, M.; Darghouth, M.A.; Gharbi, M. Molecular detection of Toxoplasma gondii and Sarcocystis spp. co-infection in tunisian merguez, a traditional processed sausage beef meat. Food Control 2021, 121, 107618. [Google Scholar] [CrossRef]
- Langoni, H.; Generoso, D.; Hayasaka, Ê.Y.; Mantovan, K.B.; Menozzi, B.D.; Richini-Pereira, V.B.; da Silva, R.C. Molecular characterization of Toxoplasma gondii and Sarcocystis spp. in raw kibbeh and other meat samples commercialized in Botucatu, Southeastern Brazil. Rev. Bras. Parasitol. Vet. 2021, 30, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Hooshyar, H.; Chehrazi, F.; Arbabi, M. Molecular identification and frequency of cyst-forming coccidia (Sarcocystis, Toxoplasma gondii, and Neospora caninum) in native slaughtered cattle in Kashan, Central Iran. Int. Arch. Health Sci. 2021, 8, 301. [Google Scholar] [CrossRef]
- Stelzer, S.; Basso, W.; Benavides Silván, J.; Ortega-Mora, L.M.; Maksimov, P.; Gethmann, J.; Conraths, F.J.; Schares, G. Toxoplasma gondii infection and toxoplasmosis in farm animals: Risk factors and economic impact. Food Waterborne Parasitol. 2019, 15, e00037. [Google Scholar] [CrossRef]
- Dubey, J.P. A Review of Toxoplasmosis in cattle. Vet. Parasitol. 1986, 22, 177–202. [Google Scholar] [CrossRef] [PubMed]
- Canada, N.; Meireles, C.S.; Rocha, A.; da Costa, J.M.C.; Erickson, M.W.; Dubey, J.P. Isolation of viable Toxoplasma gondii from naturally infected aborted bovine fetuses. J. Parasitol. 2002, 88, 1247–1248. [Google Scholar] [CrossRef]
- Hornok, S.; Mester, A.; Takács, N.; Baska, F.; Majoros, G.; Fok, É.; Biksi, I.; Német, Z.; Hornyák, Á.; Jánosi, S.; et al. Sarcocystis-infection of cattle in Hungary. Parasites Vectors 2015, 8, 4–9. [Google Scholar] [CrossRef]
- Jones, C.D.; Okhravi, N.; Adamson, P.; Tasker, S. Comparison of PCR detection methods for B1, P30, and 18S rDNA genes of T. gondii in aqueous humor. Investig. Ophthalmol. Vis. Sci. 2000, 41, 634–644. [Google Scholar]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis version 7 for bigger datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vangeel, L.; Houf, K.; Chiers, K.; Vercruysse, J.; D’Herde, K.; Ducatelle, R. Molecular-based identification of Sarcocystis hominis in Belgian minced beef. J. Food Prot. 2007, 70, 1523–1526. [Google Scholar] [CrossRef]
- Guo, M.; Dubey, J.P.; Hill, D.; Buchanan, R.L.; Ray Gamble, H.; Jones, J.L.; Pradhan, A.K. Prevalence and risk factors for Toxoplasma gondii infection in meat animals and meat products destined for human consumption. J. Food Prot. 2015, 78, 457–476. [Google Scholar] [CrossRef]
- Baril, L.; Ancelle, T.; Goulet, V.; Thulliez, P.; Tirard-Fleury, V.; Carme, B. Risk Factors for Toxoplasma Infection in pregnancy: A case-control study in France. Scand. J. Infect. Dis. 1999, 31, 305–309. [Google Scholar] [CrossRef] [PubMed]
- Cook, A.J.; Gilbert, R.E.; Buffolano, W.; Zufferey, J.; Petersen, E.; Jenum, P.A.; Foulon, W.; Semprini, A.E.; Dunn, D.T. Sources of Toxoplasma infection in pregnant women: European multicentre case-control study. european research network on congenital toxoplasmosis. BMJ 2000, 321, 142–147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.L.; Dargelas, V.; Roberts, J.; Press, C.; Remington, J.S.; Montoya, J.G. Risk Factors for Toxoplasma gondii Infection in the United States. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc. Am. 2009, 49, 878–884. [Google Scholar] [CrossRef] [Green Version]
- García-Bocanegra, I.; Cabezón, O.; Hernández, E.; Martínez-Cruz, M.S.; Martínez-Moreno, Á.; Martínez-Moreno, J. Toxoplasma gondii in ruminant species (cattle, sheep, and goats) from Southern Spain. J. Parasitol. 2013, 99, 438–440. [Google Scholar] [CrossRef]
- Berger-Schoch, A.E.; Bernet, D.; Doherr, M.G.; Gottstein, B.; Frey, C.F. Toxoplasma gondii in Switzerland: A serosurvey based on meat juice analysis of slaughtered pigs, wild boar, sheep and cattle. Zoonoses Public Health 2011, 58, 472–478. [Google Scholar] [CrossRef]
- Gazzonis, A.L.; Marino, A.M.F.; Garippa, G.; Rossi, L.; Mignone, W.; Dini, V.; Giunta, R.P.; Luini, M.; Villa, L.; Zanzani, S.A.; et al. Toxoplasma gondii seroprevalence in beef cattle raised in Italy: A multicenter study. Parasitol. Res. 2020, 119, 3893–3898. [Google Scholar] [CrossRef]
- Opsteegh, M.; Teunis, P.; Züchner, L.; Koets, A.; Langelaar, M.; van der Giessen, J. Low predictive value of seroprevalence of Toxoplasma gondii in cattle for detection of parasite DNA. Int. J. Parasitol. 2011, 41, 343–354. [Google Scholar] [CrossRef] [PubMed]
- Dubey, J.P. Isolation of Toxoplasma gondii from a naturally infected beef cow. J. Parasitol. 1992, 78, 151–153. [Google Scholar] [CrossRef] [PubMed]
- Guneratne, R.; Mendis, D.; Bandara, T.; Fernando, S.D. Toxoplasma, Toxocara and tuberculosis co-infection in a four year old child. BMC Pediatr. 2011, 11, 44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yusuf, I.H.; Sahare, P.; Hildebrand, G.D. DRESS Syndrome in a child treated for toxoplasma retinochoroiditis. J. AAPOS Off. Publ. Am. Assoc. Pediatr. Ophthalmol. Strabismus 2013, 17, 521–523. [Google Scholar] [CrossRef] [PubMed]



| Sample | 18s PCR [31] | Multiplex Sarcocystis PCR [19] | B1 Toxoplasma Nested-PCR [32] |
|---|---|---|---|
| ML (4 samples) | 4 PCR positive * (3 S. hominis, 1 Toxoplasma/Hammondia) | 4 S. hominis PCR positive (1 S. hominis) | 4 PCR Positive (no sequence) |
| HM (4 samples) | 4 PCR Positive (1 S. hominis) | 4 S. hominis PCR positive | 4 PCR Positive (no sequence) |
| MJ (1 sample) | 1 PCR Positive (1 S. hominis) | 1 S. hominis PCR positive (1 S. hominis) | 1 PCR Positive (1 T. gondii) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Dini, F.M.; Caffara, M.; Jacinto, J.G.P.; Benazzi, C.; Gentile, A.; Galuppi, R. A Case of Bovine Eosinophilic Myositis (BEM) Associated with Co-Infection by Sarcocystis hominis and Toxoplasma gondii. Animals 2023, 13, 311. https://doi.org/10.3390/ani13020311
Dini FM, Caffara M, Jacinto JGP, Benazzi C, Gentile A, Galuppi R. A Case of Bovine Eosinophilic Myositis (BEM) Associated with Co-Infection by Sarcocystis hominis and Toxoplasma gondii. Animals. 2023; 13(2):311. https://doi.org/10.3390/ani13020311
Chicago/Turabian StyleDini, Filippo Maria, Monica Caffara, Joana G. P. Jacinto, Cinzia Benazzi, Arcangelo Gentile, and Roberta Galuppi. 2023. "A Case of Bovine Eosinophilic Myositis (BEM) Associated with Co-Infection by Sarcocystis hominis and Toxoplasma gondii" Animals 13, no. 2: 311. https://doi.org/10.3390/ani13020311
APA StyleDini, F. M., Caffara, M., Jacinto, J. G. P., Benazzi, C., Gentile, A., & Galuppi, R. (2023). A Case of Bovine Eosinophilic Myositis (BEM) Associated with Co-Infection by Sarcocystis hominis and Toxoplasma gondii. Animals, 13(2), 311. https://doi.org/10.3390/ani13020311