

First Detection of Theileria sinensis-like and Anaplasma capra in Ixodes kashmiricus: With Notes on cox1-Based Phylogenetic Position and New Locality Records

 ,
,  , ,
, ,  , , , ,
, , , ,  and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
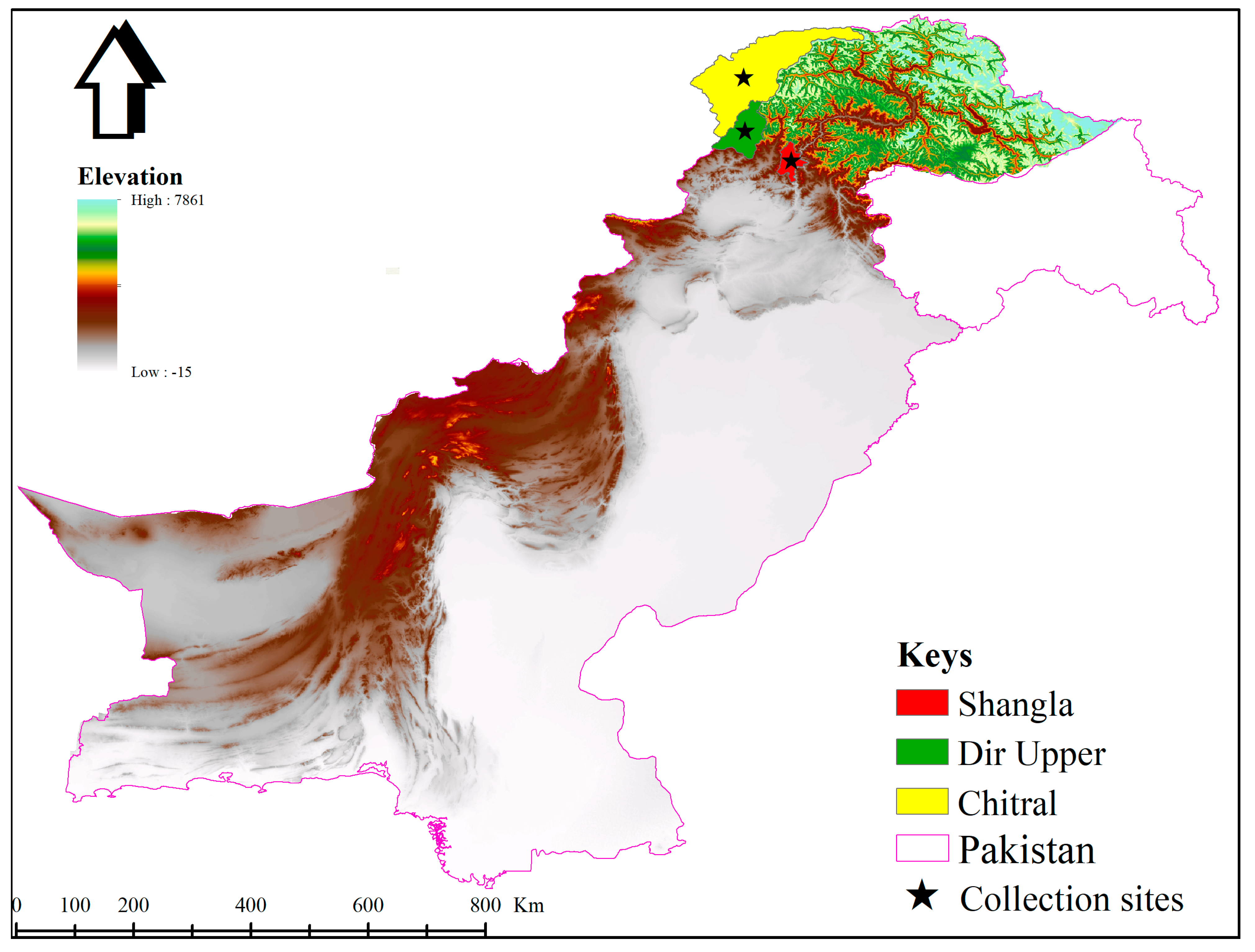
2.2. Study Area and Tick Collection
2.3. Morphological Identification of Ticks
2.4. DNA Isolation and PCR
2.5. DNA Sequencing and Phylogenetic Analysis
2.6. Statistical Analyses
3. Results
3.1. Morphological Identification and Description of Ixodes kashmiricus
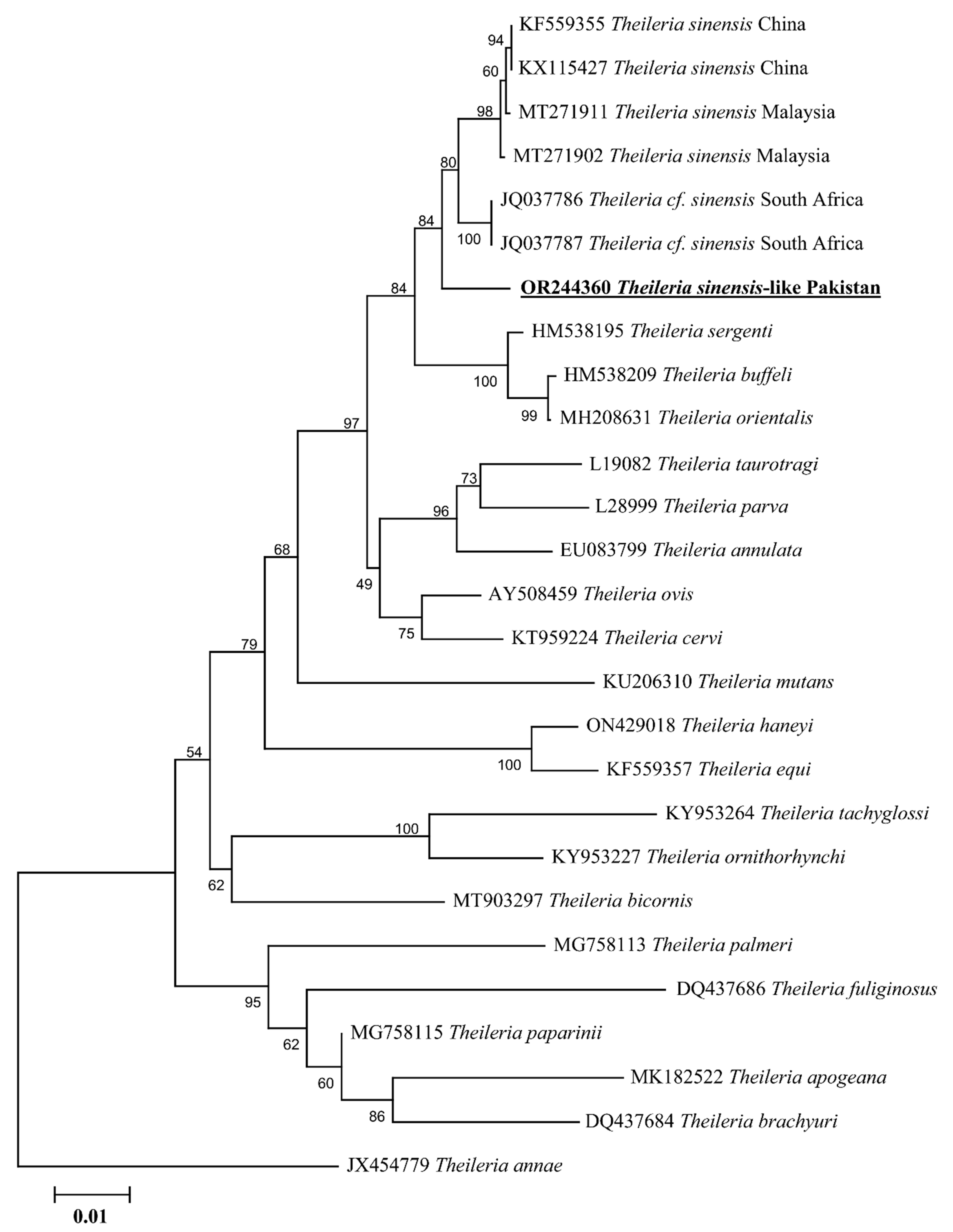
3.2. Sequences and Phylogenetic Relationship of Ticks
3.3. Sequences and Phylogenetic Relationship of Theileria spp. and Anaplasma spp.
4. Discussions
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Clifford, C.M.; Sonenshine, D.E.; Keirans, J.E.; Kohls, G.M. Systematics of the subfamily Ixodinae (Acarina: Ixodidae). 1. the subgenera of Ixodes. Ann. Entomol. Soc. Am. 1973, 66, 489–500. [Google Scholar] [CrossRef]
- Filippova, N.A. Ixodid ticks of the subfamily Ixodinae. Fauna SSSR, Paukoobraznye, Izdatielstwo Nauka, Moskwa–Leningrad. Arachnida 1977, 4, 272–283. (In Russian) [Google Scholar]
- Guglielmone, A.A.; Petney, T.N.; Robbins, R.G. Ixodidae (Acari: Ixodoidea): Descriptions and redescriptions of all known species from 1758 to December 31, 2019. Zootaxa 2020, 4871, 1–322. [Google Scholar]
- Rar, V.; Yakimenko, V.; Tikunov, A.; Vinarskaya, N.; Tancev, A.; Babkin, I.; Epikhina, T.; Tikunova, N. Genetic and morphological characterization of Ixodes apronophorus from Western Siberia, Russia. Ticks Tick Borne Dis. 2020, 11, 101284. [Google Scholar] [CrossRef] [PubMed]
- Pietzsch, M.E.; Medlock, J.M.; Jones, L.; Avenell, D.; Abbott, J.; Harding, P.; Leach, S. Distribution of Ixodes ricinus in the British Isles: Investigation of historical records. Med. Vet. Entomol. 2005, 19, 306–314. [Google Scholar] [PubMed]
- Lommano, E.; Bertaiola, L.; Dupasquier, C.; Gern, L. Infections and coinfections of questing Ixodes ricinus ticks by emerging zoonotic pathogens in Western Switzerland. Appl. Environ. Microbiol. 2012, 78, 4606–4612. [Google Scholar] [CrossRef]
- Pilloux, L.; Baumgartner, A.; Jaton, K.; Lienhard, R.; Ackermann-Gäumann, R.; Beuret, C.; Greub, G. Prevalence of Anaplasma phagocytophilum and Coxiella burnetii in Ixodes ricinus ticks in Switzerland: An underestimated epidemiologic risk. New Microbes New Infect. 2019, 27, 22–26. [Google Scholar] [CrossRef]
- Sands, B.; Lihou, K.; Lait, P.; Wall, R. Prevalence of Babesia spp. pathogens in the ticks Dermacentor reticulatus and Ixodes ricinus in the UK. Acta Trop. 2022, 236, 106692. [Google Scholar] [CrossRef]
- Guo, W.P.; Zhang, B.; Wang, Y.H.; Xu, G.; Wang, X.; Ni, X.; Zhou, E.M. Molecular identification and characterization of Anaplasma capra and Anaplasma platys-like in Rhipicephalus microplus in Ankang, Northwest China. BMC Infect. Dis. 2019, 19, 1–9. [Google Scholar]
- Altay, K.; Erol, U.; Sahin, O.F. The first molecular detection of Anaplasma capra in domestic ruminants in the central part of Turkey, with genetic diversity and genotyping of Anaplasma capra. Trop. Anim. Health Prod. 2022, 54, 129. [Google Scholar] [CrossRef]
- Aktas, M.; Vatansever, Z.; Altay, K.; Aydin, M.F.; Dumanli, N. Molecular evidence for Anaplasma phagocytophilum in Ixodes ricinus from Turkey. Trans. R. Soc. Trop. Med. Hyg. 2010, 104, 10–15. [Google Scholar] [PubMed]
- Bown, K.J.; Begon, M.; Bennett, M.; Woldehiwet, Z.; Ogden, N.H. Seasonal dynamics of Anaplasma phagocytophila in a rodent-tick (Ixodes trianguliceps) system, United Kingdom. Emerg. Infect. Dis. 2003, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Ogden, N.H.; Lindsay, L.R.; Hanincová, K.; Barker, I.K.; Bigras-Poulin, M.; Charron, D.F.; Heagy, A.; Francis, C.M.; O’Callaghan, C.J.; Schwartz, I.; et al. Role of migratory birds in introduction and range expansion of Ixodes scapularis ticks and of Borrelia burgdorferi and Anaplasma phagocytophilum in Canada. Appl. Environ. Microbiol. 2008, 74, 1780–1790. [Google Scholar]
- Remesar, S.; Diaz, P.; Prieto, A.; García-Dios, D.; Panadero, R.; Fernández, G.; Brianti, E.; Díez-Baños, P.; Morrondo, P.; López, C.M. Molecular detection and identification of piroplasms (Babesia spp.; Theileria spp.) and Anaplasma phagocytophilum in questing ticks from northwest Spain. Med. Vet. Entomol. 2021, 35, 51–58. [Google Scholar] [PubMed]
- Li, H.; Zheng, Y.C.; Ma, L.; Jia, N.; Jiang, B.G.; Jiang, R.R.; Huo, Q.B.; Wang, Y.W.; Liu, H.B.; Chu, Y.L.; et al. Human infection with a novel tick-borne Anaplasma species in China: A surveillance study. Lancet Infect. Dis. 2015, 15, 663–670. [Google Scholar] [CrossRef] [PubMed]
- Ali, A.; Ullah, S.; Numan, M.; Almutairi, M.; Alouffi, A.; Tanaka, T. First report on tick-borne pathogens detected in ticks infesting stray dogs near butcher shops. Front. Vet. Sci. 2023, 10, 1246871. [Google Scholar]
- Camacho, A.T.; Pallas, E.; Gestal, J.J.; Guitián, F.J.; Olmeda, A.S.; Telford III, S.R.; Spielman, A. Ixodes hexagonus is the main candidate as vector of Theileria annae in northwest Spain. Vet. Parasitol. 2003, 112, 157–163. [Google Scholar] [CrossRef]
- Loh, S.M.; Paparini, A.; Ryan, U.; Irwin, P.; Oskam, C. Identification of Theileria fuliginosa-like species in Ixodes australiensis ticks from western grey kangaroos (Macropus fuliginosus) in Western Australia. Ticks Tick Borne Dis. 2018, 9, 632–637. [Google Scholar]
- Wang, Y.; Wang, B.; Zhang, Q.; Li, Y.; Yang, Z.; Han, S.; Yuan, G.; Wang, S.; He, H. The common occurrence of Theileria ovis in Tibetan Sheep and the first report of Theileria sinensis in yaks from southern Qinghai, China. Acta Parasitol. 2021, 66, 1177–1185. [Google Scholar]
- Sabitova, Y.; Rar, V.; Tikunov, A.; Yakimenko, V.; Korallo-Vinarskaya, N.; Livanova, N.; Tikunova, N. Detection and genetic characterization of a putative novel Borrelia genospecies in Ixodes apronophorus/Ixodes persulcatus/Ixodes trianguliceps sympatric areas in Western Siberia. Ticks Tick Borne Dis. 2023, 14, 102075. [Google Scholar] [CrossRef]
- Numan, M.; Islam, N.; Adnan, M.; Zaman Safi, S.; Chitimia-Dobler, L.; Labruna, M.B.; Ali, A. First genetic report of Ixodes kashmiricus and associated Rickettsia sp. Parasit. Vectors 2022, 15, 378. [Google Scholar] [CrossRef] [PubMed]
- Barker, S.C.; Walker, A.R.; Campelo, D. A list of the 70 species of Australian ticks; diagnostic guides to and species accounts of Ixodes holocyclus (paralysis tick), Ixodes cornuatus (southern paralysis tick) and Rhipicephalus australis (Australian cattle tick); and consideration of the place of Australia in the evolution of ticks with comments on four controversial ideas. Int. J. Parasitol. 2014, 44, 941–953. [Google Scholar] [PubMed]
- Cheng, T.Y.; Chen, Z.; Li, Z.B.; Liu, G.H. First report of Ixodes nipponensis infection in goats in China. Vector-Borne Zoonotic Dis. 2018, 18, 575–578. [Google Scholar] [CrossRef] [PubMed]
- Lv, J.; Wu, S.; Zhang, Y.; Chen, Y.; Feng, C.; Yuan, X.; Jia, G.; Deng, J.; Wang, C.; Wang, Q.; et al. Assessment of four DNA fragments (COI, 16S rDNA, ITS2, 12S rDNA) for species identification of the Ixodida (Acari: Ixodida). Parasit. Vectors 2014, 7, 93. [Google Scholar]
- Ali, A.; Zahid, H.; Zeb, I.; Tufail, M.; Khan, S.; Haroon, M.; Bilal, M.; Hussain, M.; Alouffi, A.S.; Muñoz-Leal, S. Risk factors associated with tick infestations on equids in Khyber Pakhtunkhwa, Pakistan, with notes on Rickettsia massiliae detection. Parasit. Vectors 2021, 14, 363. [Google Scholar] [CrossRef]
- Ali, A.; Numan, M.; Khan, M.; Aiman, O.; Muñoz-Leal, S.; Chitimia-Dobler, L.; Labruna, M.B.; Nijhof, A.M. Ornithodoros (Pavlovskyella) ticks associated with a Rickettsia sp. in Pakistan. Parasit. Vectors 2022, 15, 138. [Google Scholar]
- Khan, Z.; Shehla, S.; Alouffi, A.; Kashif Obaid, M.; Zeb Khan, A.; Almutairi, M.M.; Numan, M.; Aiman, O.; Alam, S.; Ullah, S.; et al. Molecular survey and genetic characterization of Anaplasma marginale in ticks collected from livestock hosts in Pakistan. Animals 2022, 12, 1708. [Google Scholar] [CrossRef]
- Ahmad, I.; Ullah, S.; Alouffi, A.; Almutairi, M.M.; Khan, M.; Numan, M.; Safi, S.Z.; Chitimia-Dobler, L.; Tanaka, T.; Ali, A. Description of Male, Redescription of Female, Host Record, and Phylogenetic Position of Haemaphysalis danieli. Pathogens 2022, 11, 1495. [Google Scholar] [CrossRef]
- Mangold, A.J.; Bargues, M.D.; Mas-Coma, S. Mitochondrial 16S rDNA sequences and phylogenetic relationships of species of Rhipicephalus and other tick genera among Metastriata (Acari: Ixodidae). Parasitol. Res. 1998, 84, 478–484. [Google Scholar] [CrossRef]
- Kovalev, S.Y.; Golovljova, I.V.; Mukhacheva, T.A. Natural hybridization between Ixodes ricinus and Ixodes persulcatus ticks evidenced by molecular genetics methods. Ticks Tick Borne Dis. 2016, 7, 113–118. [Google Scholar] [CrossRef]
- Karim, S.; Budachetri, K.; Mukherjee, N.; Williams, J.; Kausar, A.; Hassan, M.J.; Adamson, S.; Dowd, S.E.; Apanskevich, D.; Arijo, A.; et al. A study of ticks and tick-borne livestock pathogens in Pakistan. PLoS Negl. Trop. Dis. 2017, 11, e0005681. [Google Scholar] [CrossRef] [PubMed]
- Khan, S.M.; Khan, M.; Alouffi, A.; Almutairi, M.M.; Numan, M.; Ullah, S.; Obaid, M.K.; Islam, Z.U.; Ahmed, H.; Tanaka, T.; et al. Phylogenetic Position of Haemaphysalis kashmirensis and Haemaphysalis cornupunctata, with Notes on Rickettsia spp. Genes 2023, 14, 360. [Google Scholar] [CrossRef] [PubMed]
- Pomerantzev, B.I. New ticks of the family Ixodidae. Parazitol Sborn Zool Inst Akad Nauk. SSSR 1948, 10, 20–24. (In Russian) [Google Scholar]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Folmer, O.; Hoeh, W.R.; Black, M.B.; Vrijenhoek, R.C. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotech. 1994, 3, 294–299. [Google Scholar]
- Inokuma, H.; Raoult, D.; Brouqui, P. Detection of Ehrlichia platys DNA in brown dog ticks (Rhipicephalus sanguineus) in Okinawa Island, Japan. J. Clin. Microbiol. 2000, 38, 4219–4221. [Google Scholar] [CrossRef]
- Duron, O.; Noël, V.; McCoy, K.D.; Bonazzi, M.; Sidi-Boumedine, K.; Morel, O.; Vavre, F.; Zenner, L.; Jourdain, E.; Durand, P.; et al. The recent evolution of a maternally-inherited endosymbiont of ticks led to the emergence of the Q fever pathogen, Coxiella burnetii. PLoS Pathog. 2015, 11, e1004892. [Google Scholar] [CrossRef]
- Stromdahl, E.Y.; Williamson, P.C.; Kollars, T.M., Jr.; Evans, S.R.; Barry, R.K.; Vince, M.A.; Dobbs, N.A. Evidence of Borrelia lonestari DNA in Amblyomma americanum (Acari: Ixodidae) removed from humans. J. Clin. Microbiol. 2003, 41, 5557–5562. [Google Scholar] [CrossRef]
- Hall, T.; Biosciences, I.; Carlsbad, C.J.G.B.B. BioEdit: An important software for molecular biology. GERF Bull Biosci. 2011, 2, 60–61. [Google Scholar]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547. [Google Scholar] [CrossRef]
- Edgar, R.C. MUSCLE: Multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 2004, 32, 1792–1797. [Google Scholar] [CrossRef]
- Schouls, L.M.; Van De Pol, I.; Rijpkema, S.G.; Schot, C.S. Detection and identification of Ehrlichia, Borrelia burgdorferi sensu lato, and Bartonella species in Dutch Ixodes ricinus ticks. J. Clin. Microbiol. 1999, 37, 2215–2222. [Google Scholar] [CrossRef] [PubMed]
- Parola, P.; Paddock, C.D.; Socolovschi, C.; Labruna, M.B.; Mediannikov, O.; Kernif, T.; Abdad, M.Y.; Stenos, J.; Bitam, I.; Fournier, P.E.; et al. Update on tick-borne rickettsioses around the world: A geographic approach. Clin. Microbiol. Rev. 2013, 26, 657–702. [Google Scholar] [CrossRef] [PubMed]
- Clifford, C.M.; Hoogstraal, H.; Kohls, G.M. Ixodes hyatti, n. sp.; and I. shahi, n. sp. (Acarina: Ixodidae), parasites of pikas (Lagomorpha: Ochotonidae) in the Himalayas of Nepal and West Pakistan. J. Med. Entomol. 1971, 8, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Begum, F.; Wisseman, C.L., Jr.; Casals, J. Tick-borne viruses of west Pakistan: II. Hazara virus, a new agent isolated from Ixodes redikorzevi ticks from the Kaghan valley, w. Pakistan. Am. J. Epidemiol. 1970, 92, 192–194. [Google Scholar] [CrossRef]
- Filippova, N.A. Ixodes stromi, new species of tick and its systematic position in Ixodinae. Zool. Zhurnal. 1957, 36, 864–869. (In Russian) [Google Scholar]
- Begum, F.; Wisseman, C.L., Jr.; Traub, R. Tick-borne viruses of West Pakistan: I. Isolation and general characteristics. Am. J. Epidemiol. 1970, 92, 180–191. [Google Scholar] [CrossRef]
- Hoogstraal, H. The epidemiology of tick-borne Crimean-Congo hemorrhagic fever in Asia, Europe, and Africa. J. Med. Entomol. 1979, 15, 307–417. [Google Scholar] [CrossRef]
- Gilbert, L.; Aungier, J.; Tomkins, J.L. Climate of origin affects tick (Ixodes ricinus) host-seeking behavior in response to temperature: Implications for resilience to climate change? Ecol. Evol. 2014, 4, 1186–1198. [Google Scholar] [CrossRef]
- Filippova, N.A. Forms of sympatry and possible ways of microevolution of closely related species of the group Ixodes ricinus-persulcatus (Ixodidae). Acta Zool. Litu. 2002, 12, 215–227. [Google Scholar] [CrossRef]
- Volkova, V.V.; Howey, R.; Savill, N.J.; Woolhouse, M.E. Sheep movement networks and the transmission of infectious diseases. PLoS ONE 2010, 5, e11185. [Google Scholar] [CrossRef]
- Schnittger, L.; Yin, H.; Gubbels, M.J.; Beyer, D.; Niemann, S.; Jongejan, F.; Ahmed, J.S. Phylogeny of sheep and goat Theileria and Babesia parasites. Parasitol. Res. 2003, 91, 398–406. [Google Scholar] [PubMed]
- Schnittger, L.; Rodriguez, A.E.; Florin-Christensen, M.; Morrison, D.A. Babesia: A world emerging. Infect. Genet. Evol. 2012, 12, 1788–1809. [Google Scholar] [CrossRef] [PubMed]
- Sivakumar, T.; Hayashida, K.; Sugimoto, C.; Yokoyama, N. Evolution and genetic diversity of Theileria. Infect. Genet. Evol. 2014, 27, 250–263. [Google Scholar] [CrossRef]
- Ullah, K.; Numan, M.; Alouffi, A.; Almutairi, M.M.; Zahid, H.; Khan, M.; Islam, Z.U.; Kamil, A.; Safi, S.Z.; Ahmed, H.; et al. Molecular Characterization and assessment of risk factors associated with Theileria annulata infection. Microorganisms 2022, 10, 1614. [Google Scholar] [CrossRef] [PubMed]
- Paparini, A.; Macgregor, J.; Ryan, U.M.; Irwin, P.J. First molecular characterization of Theileria ornithorhynchi Mackerras, 1959: Yet another challenge to the systematics of the piroplasms. Protist 2015, 166, 609–620. [Google Scholar] [CrossRef] [PubMed]
- Alam, S.; Khan, M.; Alouffi, A.; Almutairi, M.M.; Ullah, S.; Numan, M.; Islam, N.; Khan, Z.; Aiman, O.; Zaman Safi, S.; et al. Spatio-temporal patterns of ticks and molecular survey of Anaplasma marginale, with notes on their phylogeny. Microorganisms 2022, 10, 1663. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Organism/Marker | Primer Sequences 5′-3′ | Amplicons | Annealing Temperature | References |
|---|---|---|---|---|
| Ticks/cox1 | HC02198: TAAACTTCAGGGTGACCAAAAAATCA | 649 bp | 55 °C | [35] |
| LCO1490: GGTCAACAAATCATAAAGATATTGG | ||||
| Anaplasma spp./16S rDNA | EHR16SD: GGTACCYACAGAAGAAGTCC | 344 bp | 55 °C | [36] |
| EHR16SR: TAGCACTCATCGTTTACAGC | ||||
| Theileria spp./18S rRNA | 18S_F: GGTAATTCTAGAGCTAATACATGAGC | 897 bp | 56 °C | This study |
| 18S_R: ACAATAAAGTAAAAAACAYTTCAAAG | ||||
| Coxiella spp./groEL * | CoxGrF1: TTTGAAAAYATGGGCGCKCAAATGGT | 619 bp | 56 °C | [37] |
| CoxGrR2: CGRTCRCCAAARCCAGGTGC | ||||
| CoxGrF2: GAAGTGGCTTCGCRTACWTCAGACG | ||||
| CoxGrFR1: CCAAARCCAGGTGCTTTYAC | ||||
| Borrelia spp./flab | Fla SS: AAGAGCTGAAGAGCTTGGAATG | 354 bp | 55 °C | [38] |
| Fla RS: CTTTGATCACTTATCATTCTAATAGC |
| Location/ Districts | Host | Infested/Observed Hosts | Numbers of Ticks/Life Stages | Total Collected Ticks | p Value | Number of Ticks Subjected to DNA Isolation | Amplified cox1 for Ixodes kashmiricus | Amplified 18S rDNA for Theileria | Amplified 16S rDNA for Anaplasma | ||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Males | Adult Females | Nymphs | |||||||||
| Shangla | Sheep | 42/47 | 37 | 123 | 72 | 232 | <0.001 | 77 (17M, 38F, 22N) | 2 (1M, 1F) | 1 (1F) | 2 (1F, 1N) |
| Goats | 27/39 | 11 | 24 | 25 | 60 | 25 (13F, 12N) | 1 (1F) | 1 (1N) | 1 (1N) | ||
| Dir Upper | Sheep | 13/18 | 9 | 16 | 13 | 38 | 14 (3M, 6F, 5N) | 1 (1N) | 0 | 1 (1F) | |
| Goats | 11/16 | 5 | 11 | 5 | 21 | 5 (3F, 2N) | 1 (1N) | 0 | 0 | ||
| Chitral | Sheep | 1/5 | 0 | 1 | 0 | 1 | 0 | 0 | 0 | 0 | |
| Goats | 0/4 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | ||
| Total Sheep (%) | 56/70 (80) | 46 | 140 | 85 | 271 (77) | 91 (20M, 44F, 27N) | 3 (1M, 1F, 1N) | 1 (1F) | 3 (2F, 1N) | ||
| Total Goats (%) | 38/59 (64.4) | 16 | 35 | 30 | 81 (23) | 30 (16F, 14N) | 2 (1F, 1N) | 1 (1N) | 1 (1N) | ||
| Overall Total (%) | 94/129 (72.87) | 62 (17.6) | 175 (49.7) | 115 (32.7) | 352 (100) | 121 (20M, 60F, 41N) | 5 (1M, 2F, 2N) | 2 (1.65) (1F, 1N) | 4 (3.3) (2F, 2N) | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Numan, M.; Alouffi, A.; Almutairi, M.M.; Tanaka, T.; Ahmed, H.; Akbar, H.; Rashid, M.I.; Tsai, K.-H.; Ali, A. First Detection of Theileria sinensis-like and Anaplasma capra in Ixodes kashmiricus: With Notes on cox1-Based Phylogenetic Position and New Locality Records. Animals 2023, 13, 3232. https://doi.org/10.3390/ani13203232
Numan M, Alouffi A, Almutairi MM, Tanaka T, Ahmed H, Akbar H, Rashid MI, Tsai K-H, Ali A. First Detection of Theileria sinensis-like and Anaplasma capra in Ixodes kashmiricus: With Notes on cox1-Based Phylogenetic Position and New Locality Records. Animals. 2023; 13(20):3232. https://doi.org/10.3390/ani13203232
Chicago/Turabian StyleNuman, Muhammad, Abdulaziz Alouffi, Mashal M. Almutairi, Tetsuya Tanaka, Haroon Ahmed, Haroon Akbar, Muhammad Imran Rashid, Kun-Hsien Tsai, and Abid Ali. 2023. "First Detection of Theileria sinensis-like and Anaplasma capra in Ixodes kashmiricus: With Notes on cox1-Based Phylogenetic Position and New Locality Records" Animals 13, no. 20: 3232. https://doi.org/10.3390/ani13203232
APA StyleNuman, M., Alouffi, A., Almutairi, M. M., Tanaka, T., Ahmed, H., Akbar, H., Rashid, M. I., Tsai, K. -H., & Ali, A. (2023). First Detection of Theileria sinensis-like and Anaplasma capra in Ixodes kashmiricus: With Notes on cox1-Based Phylogenetic Position and New Locality Records. Animals, 13(20), 3232. https://doi.org/10.3390/ani13203232