

Testing the Effectiveness of the “Smelly” Elephant Repellent in Controlled Experiments in Semi-Captive Asian and African Savanna Elephants
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites
2.1.1. Thailand
2.1.2. Zambia
2.2. Smelly Elephant Repellent
2.3. Trials and Data Collection
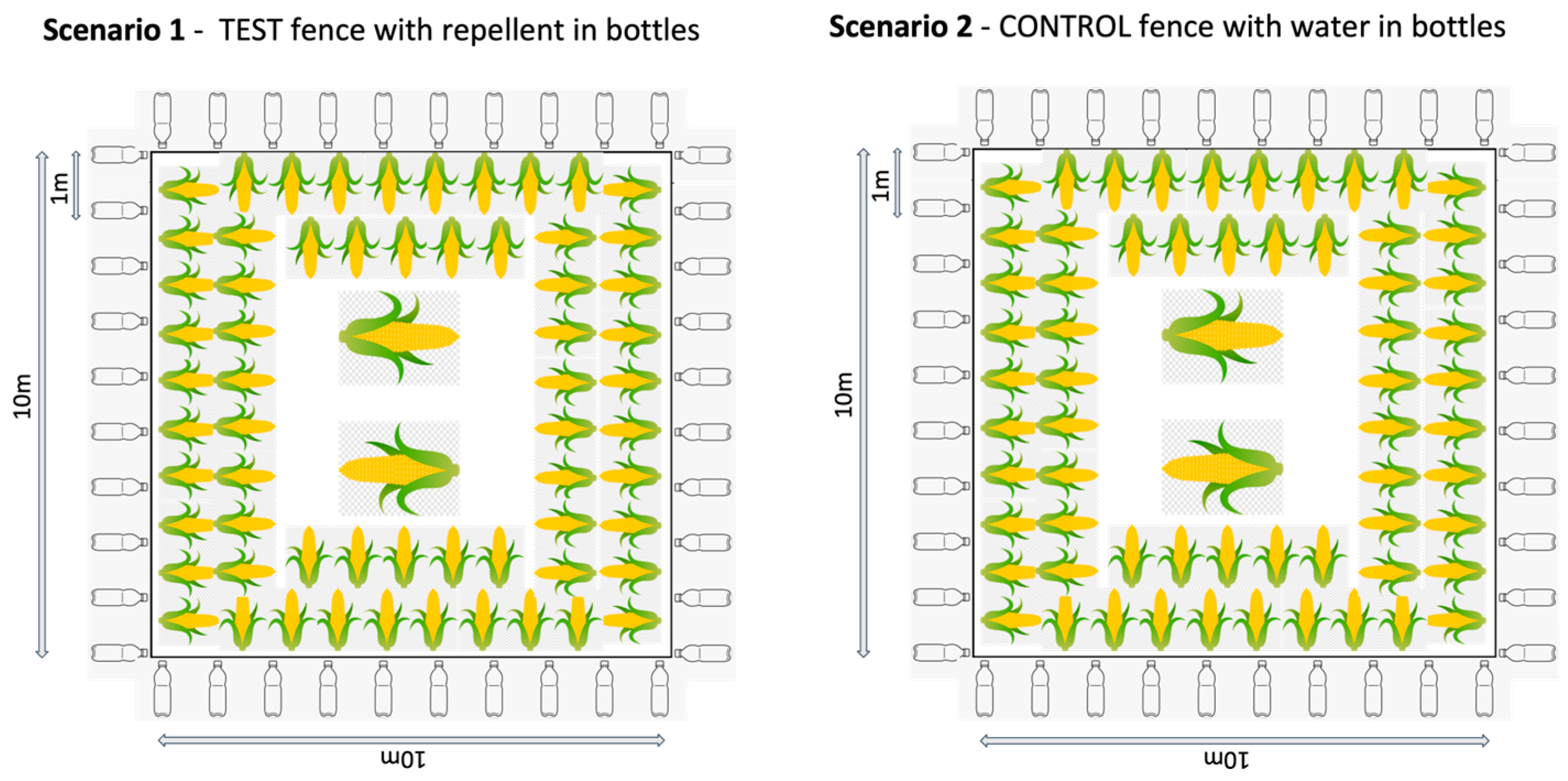
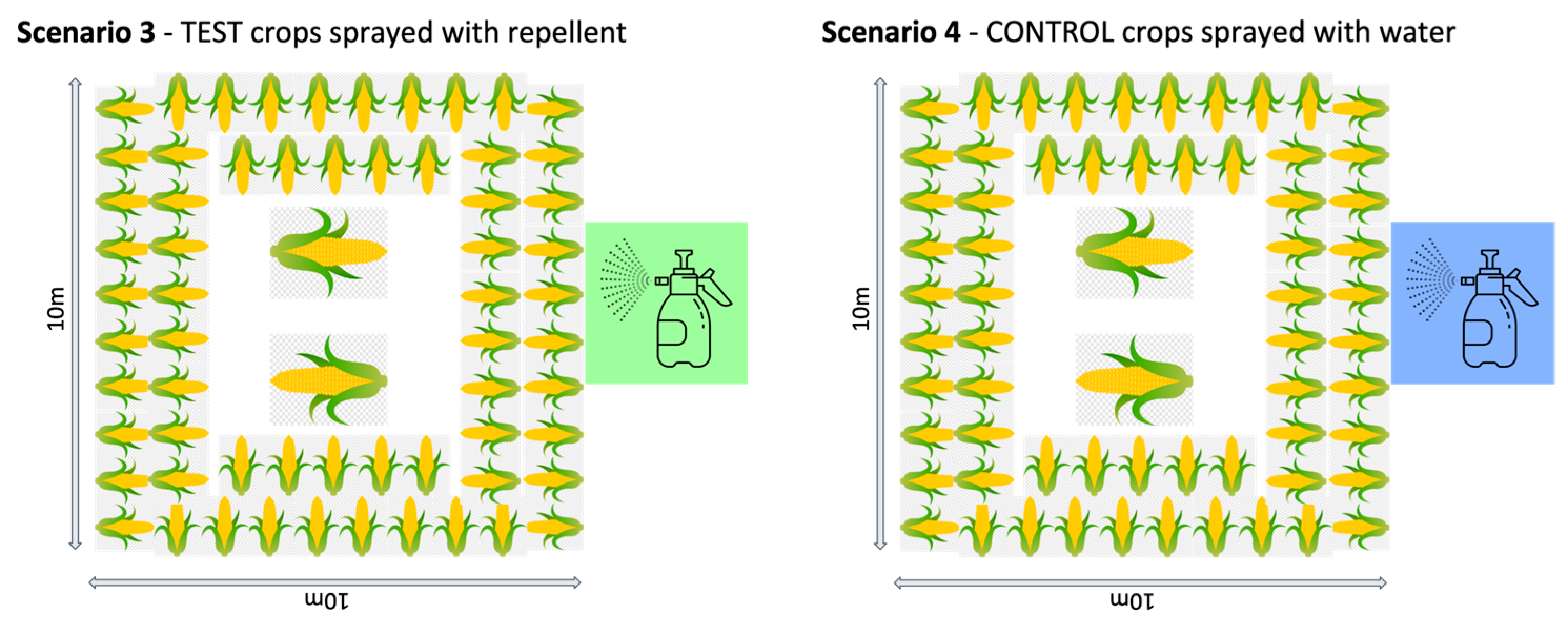
2.3.1. Thailand
- Repellent fence: The test fence with the repellent in plastic bottles. A fence line using rope was erected using 4–8 poles. Plastic bottles were hung onto the fence line at a distance of 1 m apart. The bottles were filled to roughly a quarter full with repellent and had holes punctured in them to allow the smell of the repellent to diffuse.
- Water fence: The control fence with water in plastic bottles. The same process as above was applied, however, the bottles were quarter filled with water instead of repellent.
- Repellent spray: The test crops were sprayed with repellent. The fence line around the plot was removed and the crops in the plot were dipped in the repellent. The ground was not sprayed so that the same plot could be used for each scenario.
- Water spray: The control crops were sprayed with water. The crops in the unfenced plot were dipped in water.


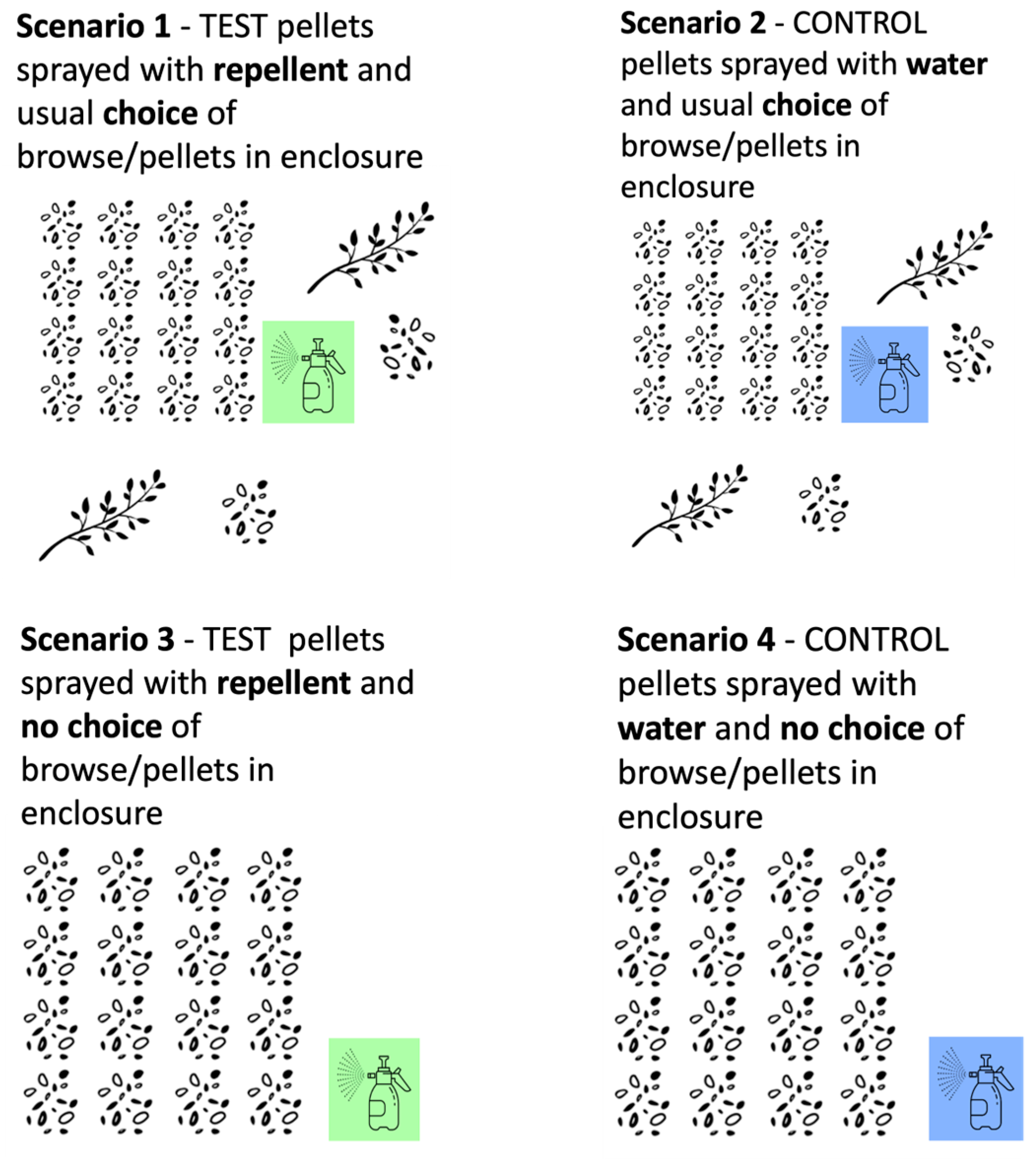
2.3.2. Zambia
- Repellent choice: The test pellets were sprayed with repellent and there was the usual choice of browse/pellets in the boma.
- Water choice: The control pellets were sprayed with water and there was the usual choice of browse/pellets in the boma.
- Repellent no-choice: The test pellets were sprayed with repellent and there was no choice of browse/pellets in the boma.
- Water no-choice: The control pellets were sprayed with water and there was no choice of browse/pellets in the boma.

2.3.3. Personality Assessment
2.3.4. Data Coding and Statistical Analysis
3. Results
3.1. Results
3.1.1. Thailand
3.1.2. Zambia
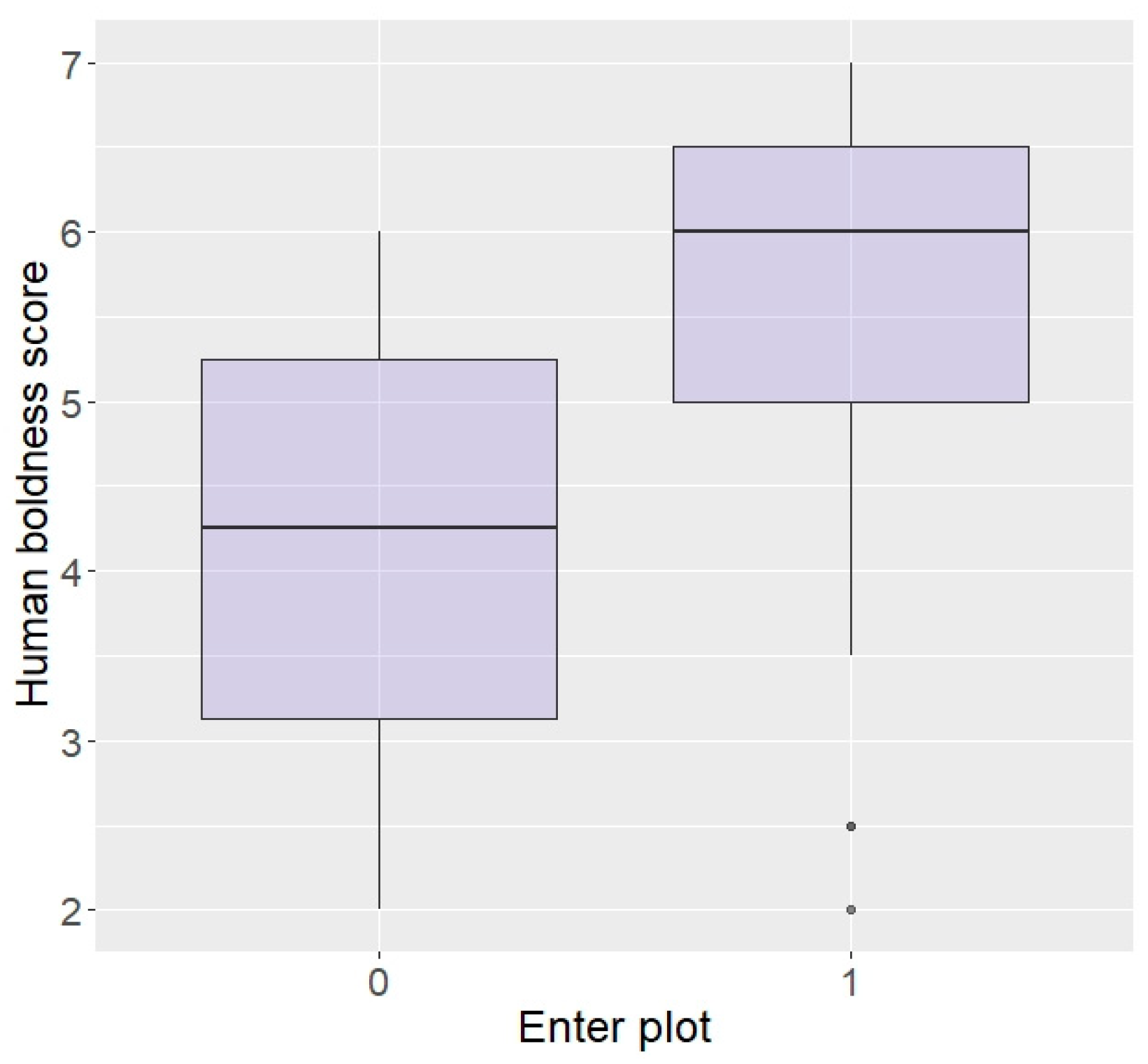
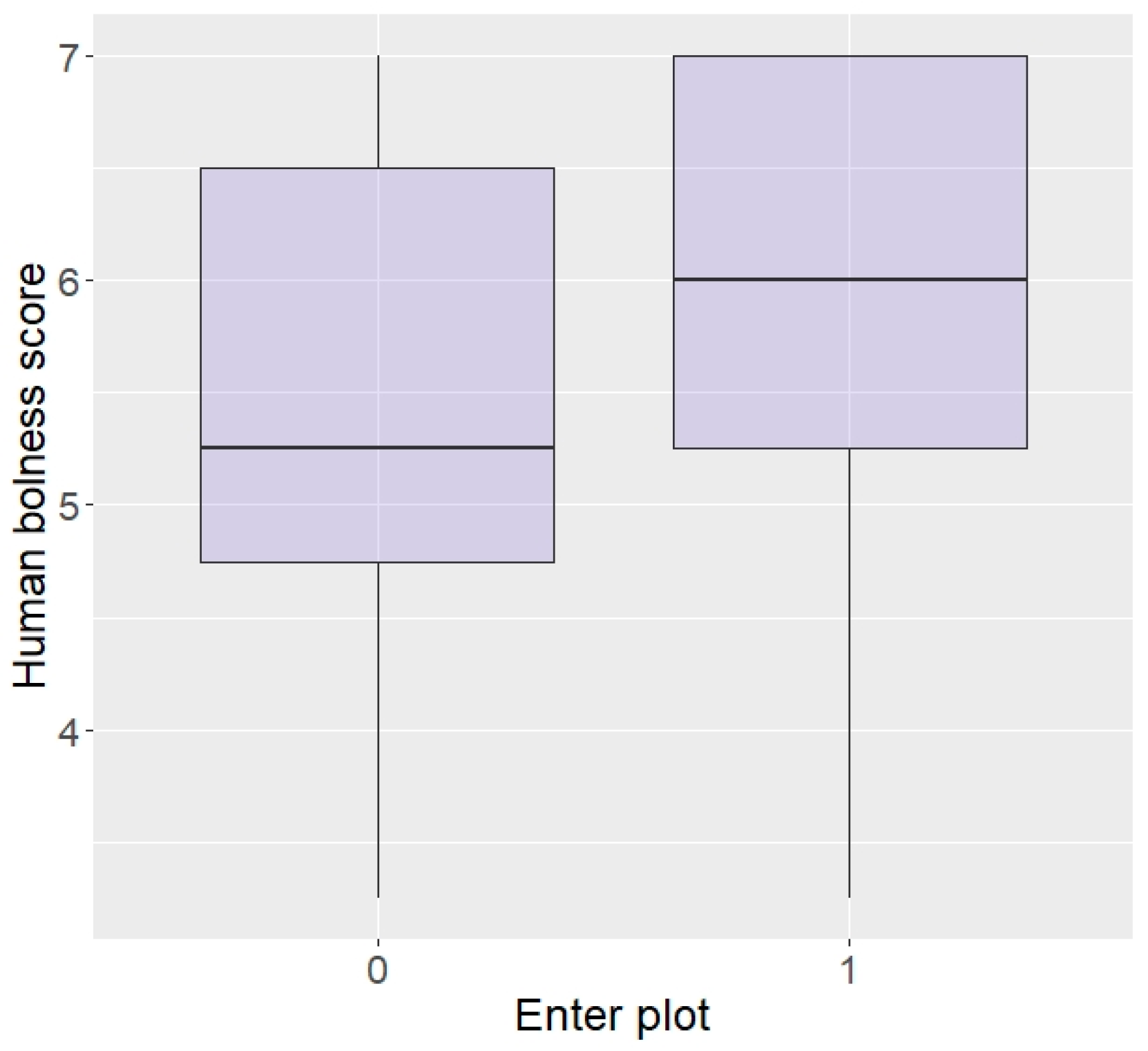
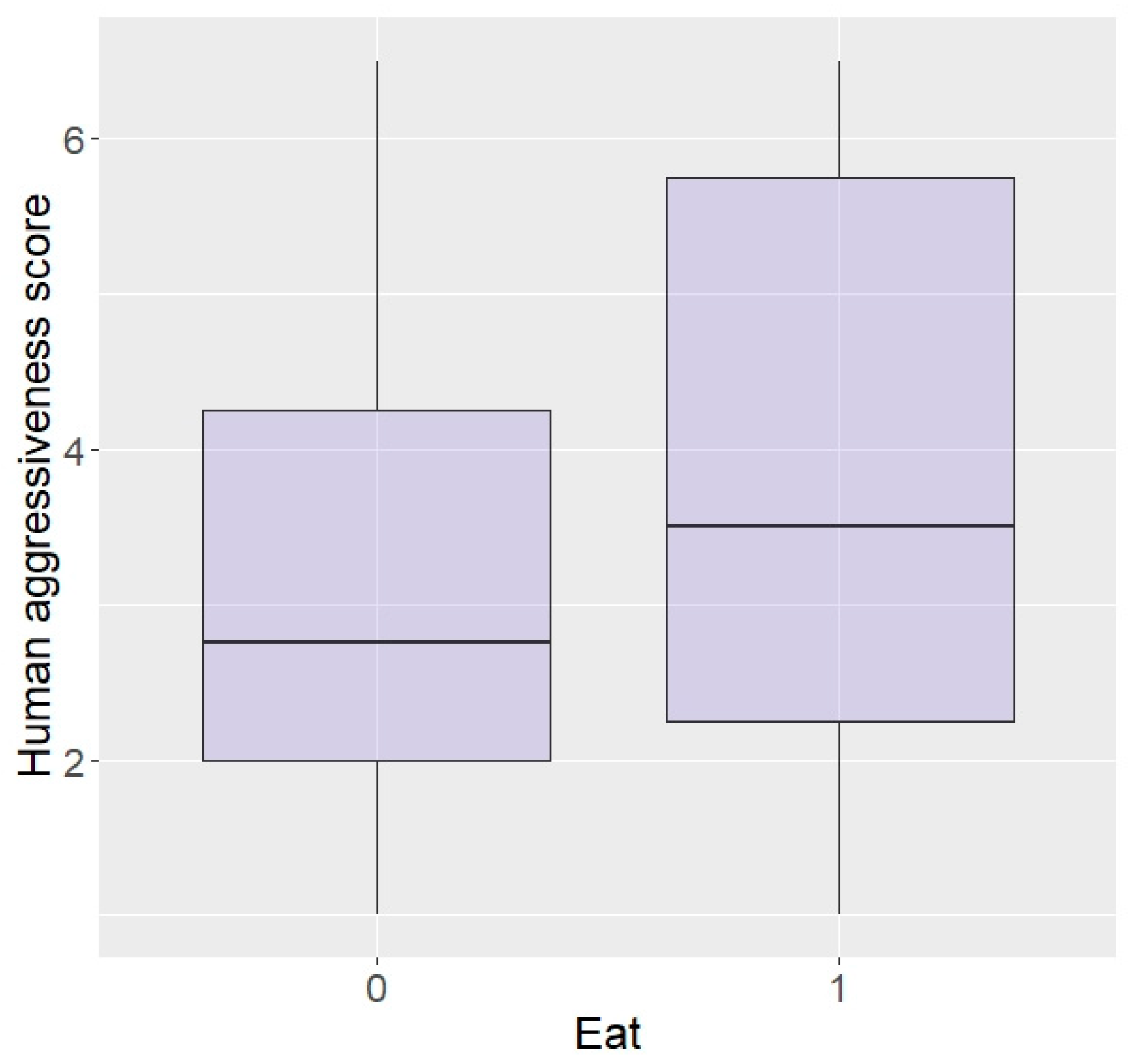
3.2. Effect of Personality
3.2.1. Thailand
3.2.2. Zambia
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Elephant | Age | Group | #RF Trials | #RS Trials | #WF Trials | #WS Trials |
|---|---|---|---|---|---|---|---|
| 1 | Yuki | 35 | 1 | 3 | 3 | 3 | 3 |
| 2 | Poonlarb | 35 | 1 | 3 | 3 | 3 | 3 |
| 3 | Beau | 42 | 2 | 3 | 3 | 3 | 3 |
| 4 | Yui | 25 | 2 | 3 | 3 | 3 | 3 |
| 5 | Pleum | 23 | 2 | 3 | 3 | 3 | 3 |
| 6 | Dah | 19 | 3 | 3 | 3 | 3 | 3 |
| 7 | Pumpui | 44 | 3 | 3 | 3 | 3 | 3 |
| 8 | Jathong | 30 | 4 | 3 | 3 | 3 | 2 |
| 9 | Kamoon | 50 | 4 | 3 | 3 | 3 | 2 |
| 10 | Maemoo | 47 | 5 | 3 | 3 | 3 | 2 |
| 11 | Kamtoon | 17 | 5 | 3 | 2 | 2 | 1 |
| 12 | Maenoi | 19 | 5 | 3 | 3 | 3 | 2 |
| 13 | Lanna | 33 | 6 | 3 | 3 | 3 | 2 |
| No. | Elephant | Sex | Age | #RC Trials | #RN Trials | #WC Trials | #WN Trials |
|---|---|---|---|---|---|---|---|
| 1 | Chamilandu | F | 15 | 3 | 4 | 4 | 4 |
| 2 | Kasewe | F | 5 | 4 | 2 | 3 | 3 |
| 3 | Kavalamanja | F | 10 | 4 | 4 | 4 | 3 |
| 4 | Ludaka | M | 4 | 2 | 2 | 3 | 3 |
| 5 | Maramba | M | 11 | 4 | 4 | 4 | 3 |
| 6 | Mkaliva | F | 5 | 4 | 4 | 4 | 3 |
| 7 | Mosi oa tunya | M | 10 | 4 | 4 | 4 | 4 |
| 8 | Mphamvu | M | 8 | 4 | 4 | 4 | 4 |
| 9 | Muchichili | M | 6 | 3 | 3 | 4 | 3 |
| 10 | Mulisani | M | 6 | 4 | 4 | 3 | 4 |
| 11 | Musolole | M | 10 | 4 | 4 | 4 | 4 |
| 12 | Mutaanzi David | M | 1 | 3 | 4 | 4 | 3 |
| 13 | Nkala | M | 8 | 4 | 4 | 3 | 3 |
| 14 | Rufunsa | M | 10 | 3 | 4 | 4 | 3 |
| 15 | Tuko | F | 3 | 2 | 2 | 3 | 3 |
Appendix B
| Behaviour | Description |
|---|---|
| Sniffing behaviour | One or more of any of the following: A Sniff-Toward: The trunk is held relatively straight and pointed in the direction of interest. Periscope-sniff: The trunk lifted up in an s shape. Hovering- Sniff: The trunk is held suspended over a particular scent on the ground or object of interest. |
| Alert behaviour | One or more of any of the following: Head shake: An abrupt shaking of the head Ears out: Fully spreads the ears (perpendicular, or almost, to its body). Tail raised: The tail is raised so that it is no longer flat against the back legs. |
| Vocalising | Any vocalisation (e.g., rumble, trumpet, or chirping). |
| Approached the plot straight away | Went within 5 m of the plot but then moved to be more than 5 m from the plot within 5 min. |
| Turned away from the plot immediately | Upon release by the mahout, the elephant moved away from the plot within 5 min |
| Strong reaction | The elephant showed sniffing and/or alert and/or vocalisation, as well as turning away immediately. |
| Did nothing | The elephant did not move towards or away from the plot. They did not show any interest in the plot. They may have grazed in the area (not in the plot) or stood still. |
| Other | It was noted if the elephant did any behaviour not included in this list. |
References
- De Silva, S.; Wu, T.; Nyhus, P.; Weaver, A.; Thieme, A.; Johnson, J.; Wadey, J.; Mossbrucker, A.; Vu, T.; Neang, T.; et al. Land-use change is associated with multi-century loss of elephant ecosystems in Asia. Sci. Rep. 2023, 13, 5996. [Google Scholar] [CrossRef] [PubMed]
- Wall, J.; Wittemyer, G.; Klinkenberg, B.; LeMay, V.; Blake, S.; Strindberg, S.; Henley, M.; Vollrath, F.; Maisels, F.; Ferwerda, J.; et al. Human footprint and protected areas shape elephant range across Africa. Curr. Biol. 2021, 31, 2437–2445. [Google Scholar] [CrossRef] [PubMed]
- IUCN. IUCN SSC Guidelines on Human-Wildlife Conflict and Coexistence, 1st ed.; IUCN: Gland, Switzerland, 2021. [Google Scholar]
- Balmford, A.; Moore, J.L.; Brooks, T.; Burgess, N.; Hansen, L.A.; Williams, P.; Rahbek, C. Conservation conflicts across Africa. Science 2001, 291, 2616–2619. [Google Scholar] [CrossRef] [PubMed]
- Madden, F. Creating Coexistence between Humans and Wildlife: Global Perspectives on Local Efforts to Address Human–Wildlife Conflict. Hum. Dimens. Wildl. 2004, 9, 247–257. [Google Scholar] [CrossRef]
- Nyhus, P.J. Human–Wildlife Conflict and Coexistence. Annu. Rev. Environ. Resour. 2016, 41, 143–171. [Google Scholar] [CrossRef]
- Woodroffe, R.; Thirgood, S.; Rabinowitz, A. (Eds.) People and Wildlife Conflict or Coexistence? Cambridge University Press: Cambridge, UK, 2005. [Google Scholar]
- Salerno, J.; Stevens, F.R.; Gaughan, A.E.; Hilton, T.; Bailey, K.; Bowles, T.; Cassidy, L.; Mupeta-Muyamwa, P.; Biggs, D.; Pricope, N.; et al. Wildlife impacts and changing climate pose compounding threats to human food security. Curr. Biol. 2021, 31, 5077–5085. [Google Scholar] [CrossRef]
- Barua, M.; Bhagwat, S.A.; Jadhav, S. The hidden dimensions of human-wildlife conflict: Health impacts, opportunity and transaction costs. Biol. Conserv. 2013, 157, 309–316. [Google Scholar] [CrossRef]
- Choudhury, A. Human–Elephant Conflicts in Northeast India. Hum. Dimens. Wildl. 2004, 9, 261–270. [Google Scholar] [CrossRef]
- Mijele, D.; Obanda, V.; Omondi, P.; Soriguer, R.C.; Gakuya, F.; Otiende, M.; Hongo, P.; Alasaad, S. Spatio-temporal distribution of injured elephants in masai mara and the putative negative and positive roles of the local community. PLoS ONE 2013, 8, e71179. [Google Scholar] [CrossRef]
- Dickman, A.J. Complexities of conflict: The importance of considering social factors for solving human-wildlife conflict. Anim. Conserv. 2010, 13, 458–466. [Google Scholar] [CrossRef]
- Pooley, S.; Barua, M.; Beinart, W.; Dickman, A.; Holmes, G.; Lorimer, J.; Loveridge, A.J.; Macdonald, D.W.; Marvin, G.; Redpath, S.; et al. An interdisciplinary review of current and future approaches to improving human–predator relations. Conserv. Biol. 2017, 31, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Shaffer, L.J.; Khadka, K.K.; Van Den Hoek, J.; Naithani, K.J. Human–elephant conflict: A review of current management strategies and future directions. Front. Ecol. Evol. 2019, 6, 235. [Google Scholar] [CrossRef]
- Wettasin, M.; Chaiyarat, R.; Youngpoy, N.; Jieychien, N.; Sukmasuang, R.; Tanhan, P. Environmental factors induced crop raiding by wild Asian elephant (Elephas maximus) in the Eastern Economic Corridor, Thailand. Sci. Rep. 2023, 13, 13388. [Google Scholar] [CrossRef] [PubMed]
- Branco, P.S.; Merkle, J.A.; Pringle, R.M.; Pansu, J.; Potter, A.B.; Reynolds, A.; Stalmans, M.; Long, R.A. Determinants of elephant foraging behaviour in a coupled human natural system: Is brown the new green? J. Anim. Ecol. 2019, 88, 780–792. [Google Scholar] [CrossRef]
- Rode, K.D.; Chiyo, P.I.; Chapman, C.A.; McDowell, L.R. Nutritional ecology of elephants in Kibale National Park, Uganda, and its relationship with crop raiding behaviour. J. Trop. Ecol. 2006, 22, 441–449. [Google Scholar] [CrossRef]
- Sukumar, R. Ecology of the Asian elephant in southern India II: Feeding habits and crop-raiding patterns. J. Trop. Ecol. 1990, 6, 33–53. [Google Scholar] [CrossRef]
- Sukumar, R.; Gadgil, M. Male-female differences in foraging on crops by Asian elephants. Anim. Behav. 1988, 36, 1233–1235. [Google Scholar] [CrossRef]
- Pokharel, S.S.; Singh, B.; Seshagiri, P.B.; Sukumar, R. Lower levels of glucocorticoids in crop-raiders: Diet quality as a potential ‘pacifier’ against stress in free-ranging Asian elephants in a human-production habitat. Anim. Conserv. 2019, 22, 177–188. [Google Scholar] [CrossRef]
- Chiyo, P.I.; Cochrane, E.P. Population structure and behaviour of crop-raiding elephants in Kibale National Park, Uganda. Afr. J. Ecol. 2005, 43, 233–241. [Google Scholar] [CrossRef]
- Osborn, F.V. Seasonal variation of feeding patterns and food selection by crop-raiding elephants in Zimbabwe. Afr. J. Ecol. 2004, 42, 322–327. [Google Scholar] [CrossRef]
- Chiyo, P.I.; Lee, P.C.; Moss, C.J.; Archie, E.A.; Hollister-Smith, J.A.; Alberts, S. No risk, no gain: Effects of crop raiding and genetic diversity on body size in male elephants. Behav. Ecol. 2011, 22, 552–558. [Google Scholar] [CrossRef]
- Douglas-Hamilton, I.; Krink, T.; Vollrath, F. Movements and corridors of African elephants in relation to protected areas. Naturwissenschaften 2005, 92, 158–163. [Google Scholar] [CrossRef] [PubMed]
- Graham, M.D.; Douglas-Hamilton, I.; Adams, W.M.; Lee, P.C. The movement of African elephants in a human-dominated land-use mosaic. Anim. Conserv. 2009, 12, 445–455. [Google Scholar] [CrossRef]
- Ihwagi, F.W.; Skidmore, A.K.; Wang, T.; Bastille-Rousseau, G.; Toxopeus, A.G.; Douglas-Hamilton, I. Poaching lowers elephant path tortuosity: Implications for conservation. J. Wildl. Manag. 2019, 83, 1022–1031. [Google Scholar] [CrossRef]
- Von Hagen, R.L.; Kasaine, S.; Githiru, M.; Amakobe, B.; Mutwiwa, U.N.; Schulte, B.A. Metal strip fences for preventing African elephant (Loxodonta africana) crop foraging in the Kasigau Wildlife Corridor, Kenya. Afr. J. Ecol. 2021, 59, 293–298. [Google Scholar] [CrossRef]
- Adams, T.S.; Mwezi, I.; Jordan, N.R. Panic at the disco: Solar-powered strobe light barriers reduce field incursion by African elephants Loxodonta africana in Chobe District, Botswana. Oryx 2021, 55, 739–746. [Google Scholar] [CrossRef]
- La Grange, M.; Matema, C.; Nyamukure, B.; Hoare, R. The Virtual Fence Dynamic: A Breakthrough for Low-Cost and Sustainable Mitigation of Human-Elephant Conflict in Subsistence Agriculture? Front. Conserv. Sci. 2022, 3, 863180. [Google Scholar] [CrossRef]
- Tiller, L.N.; Oniba, E.; Opira, G.; Brennan, E.J.; King, L.E.; Ndombi, V.; Wanjala, D.; Robertson, M.R. “Smelly” Elephant Repellent: Assessing the Efficacy of a Novel Olfactory Approach to Mitigating Elephant Crop Raiding in Uganda and Kenya. Diversity 2022, 14, 509. [Google Scholar] [CrossRef]
- Kiffner, C.; Schaal, I.; Cass, L.; Peirce, K.; Sussman, O.; Grueser, A.; Wachtel, E.; Adams, H.; Clark, K.; Konig, H.J.; et al. Perceptions and realities of elephant crop raiding and mitigation methods. Conserv. Sci. Pract. 2021, 3, 372. [Google Scholar] [CrossRef]
- Mumby, H.S.; Plotnik, J.M. Taking the elephants’ perspective: Remembering elephant behavior, cognition and ecology in human- elephant conflict mitigation. Front. Ecol. Evol. 2018, 6, 122. [Google Scholar] [CrossRef]
- Barrett, L.P.; Stanton, L.; Benson-Amram, S. The cognition of ‘nuisance’ species. Anim. Behav. 2019, 147, 167–177. [Google Scholar] [CrossRef]
- Mutinda, M.; Chenge, G.; Gakuya, F.; Otiende, M.; Omondi, P.; Kasiki, S.; Soriguer, R.C.; Alasaad, S. Detusking fence-breaker elephants as an approach in human-elephant conflict mitigation. PLoS ONE 2014, 9, e91749. [Google Scholar] [CrossRef] [PubMed]
- Plotnik, J.M.; Jacobson, S.L. A “thinking animal” in conflict: Studying wild elephant cognition in the shadow of anthropogenic change. Curr. Opin. Behav. Sci. 2022, 46, 101148. [Google Scholar] [CrossRef]
- Niimura, Y.; Matsui, A.; Touhara, K. Extreme expansion of the olfactory receptor gene repertoire in African elephants and evolutionary dynamics of orthologous gene groups in 13 placental mammals. Genome Res. 2014, 24, 1485–1496. [Google Scholar] [CrossRef]
- Bates, L.A.; Sayialel, K.N.; Njiraini, N.W.; Poole, J.H.; Moss, C.J.; Byrne, R.W. African elephants have expectations about the locations of out-of-sight family members. Biol. Lett. 2008, 4, 34–36. [Google Scholar] [CrossRef]
- Plotnik, J.M.; Brubaker, D.L.; Dale, R.; Tiller, L.N.; Mumby, H.S.; Clayton, N.S. Elephants have a nose for quantity. Proc. Natl. Acad. Sci. USA 2019, 116, 12566–12571. [Google Scholar] [CrossRef]
- Schulte, B.A.; LaDue, C.A. The Chemical Ecology of Elephants: 21st Century Additions to Our Understanding and Future Outlooks. Animals 2021, 11, 2860. [Google Scholar] [CrossRef]
- Santiapillai, C.; Read, B. Would masking the smell of ripening paddy-fields help mitigate human–elephant conflict in Sri Lanka? Oryx 2010, 44, 509–511. [Google Scholar] [CrossRef]
- Osborn, F.V.; Rasmussen, L.E.L. Evidence for the effectiveness of an oleo-resin capsicum aerosol as a repellent against wild elephants in Zimbabwe. Pachyderm 1995, 20, 55–64. [Google Scholar]
- Osborn, F.V. Capsicum oleoresin as an elephant repellent: Field trials in the communal lands of Zimbabwe. J. Wildl. Manag. 2002, 66, 674–677. [Google Scholar] [CrossRef]
- Wright, M.G.; Spencer, C.; Cook, R.M.; Henley, M.D.; North, W.; Mafra-Neto, A. African bush elephants respond to a honeybee alarm pheromone blend. Curr. Biol. 2018, 28, 778–780. [Google Scholar] [CrossRef] [PubMed]
- Valenta, K.; Schmitt, M.H.; Ayasse, M.; Nevo, O. The sensory ecology of fear: African elephants show aversion to olfactory predator signals. Conserv. Sci. Pract. 2021, 3, 333. [Google Scholar] [CrossRef]
- Osborn, F.; Parker, G. Community-based methods to reduce crop loss to elephants: Experiments in the communal lands of Zimbabwe. Pachyderm 2002, 33, 32–38. [Google Scholar]
- Parker, G.E.; Osborn, F.V. Investigating the potential for chilli Capsicum spp. to reduce human-wildlife conflict in Zimbabwe. Oryx 2006, 40, 343–346. [Google Scholar] [CrossRef]
- Pozo, R.A.; Coulson, T.; McCulloch, G.; Stronza, A.; Songhurst, A. Chilli briquettes modify the temporal behaviour of elephants, but not their numbers. Oryx 2017, 53, 100–108. [Google Scholar] [CrossRef]
- Karidozo, M.; Osborn, F.V. Community based conflict mitigation trials: Results of field test of chilli as an elephant deterrent. Biodivers. Endanger. Species 2014, 3, 144. [Google Scholar]
- Hedges, S.; Gunaryadi, D. Chilli-based elephant deterrents. Oryx 2009, 44, 139–146. [Google Scholar] [CrossRef]
- Oniba, E.; Robertson, M.R. Trialling a new scent-based repellent to mitigate elephant crop-raiding around Murchison Falls National Park, Uganda. Pachyderm 2019, 60, 123–125. [Google Scholar]
- Snyder, K.D.; Renstch, D. Rethinking assessment of success of mitigation strategies for elephant-induced crop damage. Conserv. Biol. 2020, 34, 829–842. [Google Scholar] [CrossRef]
- Sitati, N.W.; Walpole, M.J. Assessing farm-based measures for mitigating human-elephant conflict in Transmara District, Kenya. Oryx 2006, 40, 279–286. [Google Scholar] [CrossRef]
- Horback, K.M.; Miller, L.J.; Kuczaj II, S.A. Personality assessment in African elephants (Loxodonta africana): Comparing the temporal stability of ethological coding versus trait rating. Appl. Anim. Behav. Sci. 2013, 149, 55–62. [Google Scholar] [CrossRef]
- Quesada, J.; Chavez-Zichinelli, C.A.; Garcia-Arroyo, M.; Yeh, P.J.; Guevara, R.; Izquierdo-Palma, J.; MacGregor-Fors, I. Bold or shy? Examining the risk-taking behavior and neophobia of invasive and non-invasive house sparrows. Anim. Biodivers. Conserv. 2022, 45, 97–106. [Google Scholar] [CrossRef]
- Darrow, P.A.; Shivik, J.A. Bold, shy, and persistent: Variable coyote response to light and sound stimuli. Appl. Anim. Behav. Sci. 2009, 116, 82–87. [Google Scholar] [CrossRef]
- Bates, D.; Mächler, M.; Bolker, B.; Walker, S. Fitting Linear Mixed-Effects Models Using lme4. J. Stat. Softw. 2015, 67, 1–48. [Google Scholar] [CrossRef]
- King, L.E.; Lala, F.; Nzumu, H.; Mwambingu, E.; Douglas-Hamilton, I. Beehive fences as a multidimensional conflict-mitigation tool for farmers coexisting with elephants. Conserv. Biol. 2017, 31, 743–752. [Google Scholar] [CrossRef]
- Van de Water, A.; King, L.E.; Arkajak, R.; Arkajak, J.; van Doormaal, N.; Ceccarelli, V.; Sluiter, L.; Doornwaard, S.M.; Praet, V.; Owen, D.; et al. Beehive fences as a sustainable local solution to human-elephant conflict in Thailand. Conserv. Sci. Pract. 2020, 2, e260. [Google Scholar] [CrossRef]
- Dror, S.; Harich, F.; Duangphakdee, O.; Savini, T.; Pogány, Á.; Roberts, J.; Geheran, J.; Treydte, A.C. Are Asian elephants afraid of honeybees? Experimental studies in northern Thailand. Mamm. Biol. 2020, 100, 355–363. [Google Scholar] [CrossRef]
- Benson-Amram, S.; Weldele, M.L.; Holekamp, K.E. A comparison of innovative problem-solving abilities between wild and captive spotted hyaenas, Crocuta crocuta. Anim. Behav. 2013, 85, 349–356. [Google Scholar] [CrossRef]
- McCune, K.B.; Jablonski, P.; Lee, S.I.; Ha, R.R. Captive jays exhibit reduced problem-solving performance compared to wild conspecifics. R. Soc. Open Sci. 2019, 6, 181311. [Google Scholar] [CrossRef]
- Liehrmann, O.; Crawley, J.A.; Seltmann, M.W.; Feillet, S.; Nyein, U.K.; Aung, H.H.; Htut, W.; Lahdenperä, M.; Lansade, L.; Lummaa, V. Handler familiarity helps to improve working performance during novel situations in semi-captive Asian elephants. Sci. Rep. 2021, 11, 15480. [Google Scholar] [CrossRef]
- Huber, L.; Gajdon, G.K. Technical intelligence in animals: The kea model. Anim. Cogn. 2006, 9, 295–305. [Google Scholar] [CrossRef] [PubMed]
- Van de Waal, E.; Bshary, R. Contact with human facilities appears to enhance technical skills in wild vervet monkeys (Chlorocebus aethiops). Folia Primatol. 2019, 81, 282–291. [Google Scholar] [CrossRef]
- Biro, P.A.; Dingemanse, N.J. Sampling bias resulting from animal personality. Trends Ecol. Evol. 2008, 24, 66–67. [Google Scholar] [CrossRef] [PubMed]
- Sih, A.; Del Giudice, M. Linking behavioural syndromes and cognition: A behavioural ecology perspective. Philos. Trans. R. Soc. B 2012, 367, 2762–2772. [Google Scholar] [CrossRef] [PubMed]
- Day, R.L.; Coe, R.L.; Kendal, J.R.; Laland, K.N. Neophilia, innovation and social learning: A study of intergeneric differences in callitrichid monkeys. Anim. Behav. 2003, 65, 559–571. [Google Scholar] [CrossRef]
- Goldenberg, S.Z.; Douglas-Hamilton, I.; Daballen, D.; Wittemyer, G. Challenges of using behavior to monitor anthropogenic impacts on wildlife: A case study on illegal killing of African elephants. Anim. Conserv. 2017, 20, 215–224. [Google Scholar] [CrossRef]
- Poole, J.H.; Granli, P. Signals, Gestures, and Behavior of African Elephants. In The Amboseli Elephants; Moss, C.J., Ed.; University of Chicago Press: Chicago, IL, USA, 2011; pp. 109–124. [Google Scholar]




| Behaviour | Repellent Fence | Water Fence | Repellent Spray | Water Spray | Model p-Value |
|---|---|---|---|---|---|
| Sniffing | 74.36 | 60.53 | 97.37 | 37.50 | p < 0.0001 |
| Alert | 35.90 | 15.79 | 21.05 | 0.00 | p = 0.0001 |
| Vocalisation | 46.15 | 42.11 | 50.00 | 18.75 | p = 0.02 |
| Approach immediate | 66.67 | 81.58 | 100.00 | 100.00 | p < 0.0001 |
| Approach then turn | 58.97 | 34.21 | 63.16 | 31.25 | p = 0.007 |
| Turn away immediate | 5.13 | 2.63 | 7.89 | 0.00 | p > 0.05 |
| Strong reaction | 2.56 | 2.63 | 13.16 | 0.00 | p = 0.04 |
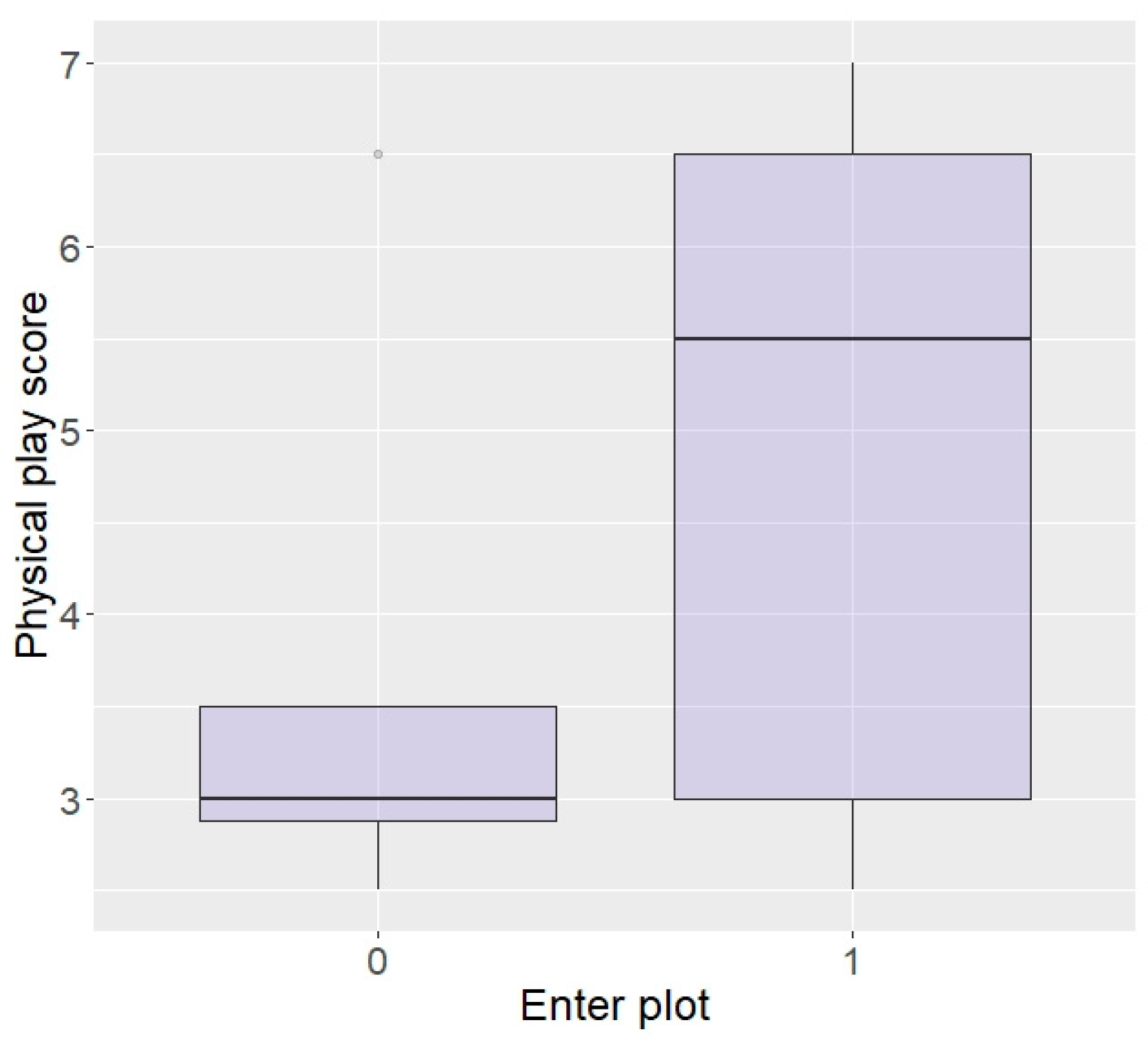
| Enter plot | 79.49 | 89.47 | 100.00 | 100.00 | p < 0.0001 |
| Eat | 82.05 | 82.86 | 74.29 | 100.00 | p = 0.0009 |
| Behaviour | Repellent Choice | Repellent No Choice | Water Choice | Water No Choice | Model p-Value |
|---|---|---|---|---|---|
| Sniffing | 53.85 | 83.02 | 65.45 | 58.00 | p = 0.005 |
| Alert | 34.62 | 26.42 | 29.09 | 32.00 | p > 0.05 |
| Vocalisation | 19.23 | 20.75 | 14.55 | 14.00 | p > 0.05 |
| Approach immediate | 36.54 | 26.42 | 18.18 | 36.00 | p = 0.03 |
| Approach then turn | 1.92 | 1.89 | 5.45 | 6.00 | p > 0.05 |
| Turn away immediate | 0.00 | 3.77 | 3.64 | 0.00 | - |
| Strong reaction | 0.00 | 1.89 | 0.00 | 0.00 | - |
| Enter plot | 29.41 | 66.04 | 23.64 | 48.00 | p < 0.0001 |
| Eat | 21.57 | 33.96 | 34.55 | 60.00 | p < 0.0001 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Robertson, M.R.; Olivier, L.J.; Roberts, J.; Yonthantham, L.; Banda, C.; N’gombwa, I.B.; Dale, R.; Tiller, L.N. Testing the Effectiveness of the “Smelly” Elephant Repellent in Controlled Experiments in Semi-Captive Asian and African Savanna Elephants. Animals 2023, 13, 3334. https://doi.org/10.3390/ani13213334
Robertson MR, Olivier LJ, Roberts J, Yonthantham L, Banda C, N’gombwa IB, Dale R, Tiller LN. Testing the Effectiveness of the “Smelly” Elephant Repellent in Controlled Experiments in Semi-Captive Asian and African Savanna Elephants. Animals. 2023; 13(21):3334. https://doi.org/10.3390/ani13213334
Chicago/Turabian StyleRobertson, Marion R., Lisa J. Olivier, John Roberts, Laddawan Yonthantham, Constance Banda, Innocent B. N’gombwa, Rachel Dale, and Lydia N. Tiller. 2023. "Testing the Effectiveness of the “Smelly” Elephant Repellent in Controlled Experiments in Semi-Captive Asian and African Savanna Elephants" Animals 13, no. 21: 3334. https://doi.org/10.3390/ani13213334
APA StyleRobertson, M. R., Olivier, L. J., Roberts, J., Yonthantham, L., Banda, C., N’gombwa, I. B., Dale, R., & Tiller, L. N. (2023). Testing the Effectiveness of the “Smelly” Elephant Repellent in Controlled Experiments in Semi-Captive Asian and African Savanna Elephants. Animals, 13(21), 3334. https://doi.org/10.3390/ani13213334