


A Functional Variant Alters the Binding of Bone morphogenetic protein 2 to the Transcription Factor NF-κB to Regulate Bone morphogenetic protein 2 Gene Expression and Chicken Abdominal Fat Deposition

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Statement
2.2. Experimental Animals and Sample Collection
2.3. Construction of Luciferase Reporter Vector
2.4. Bioinformatics Analysis
2.5. Designing of Primers
2.6. Cell Culture
2.7. Construction and Validation of Eukaryotic Expression Vector
2.8. Luciferase Reporter Assay
2.9. Electrophoretic Mobility Shift Analysis
2.10. Statistical Analysis
3. Results
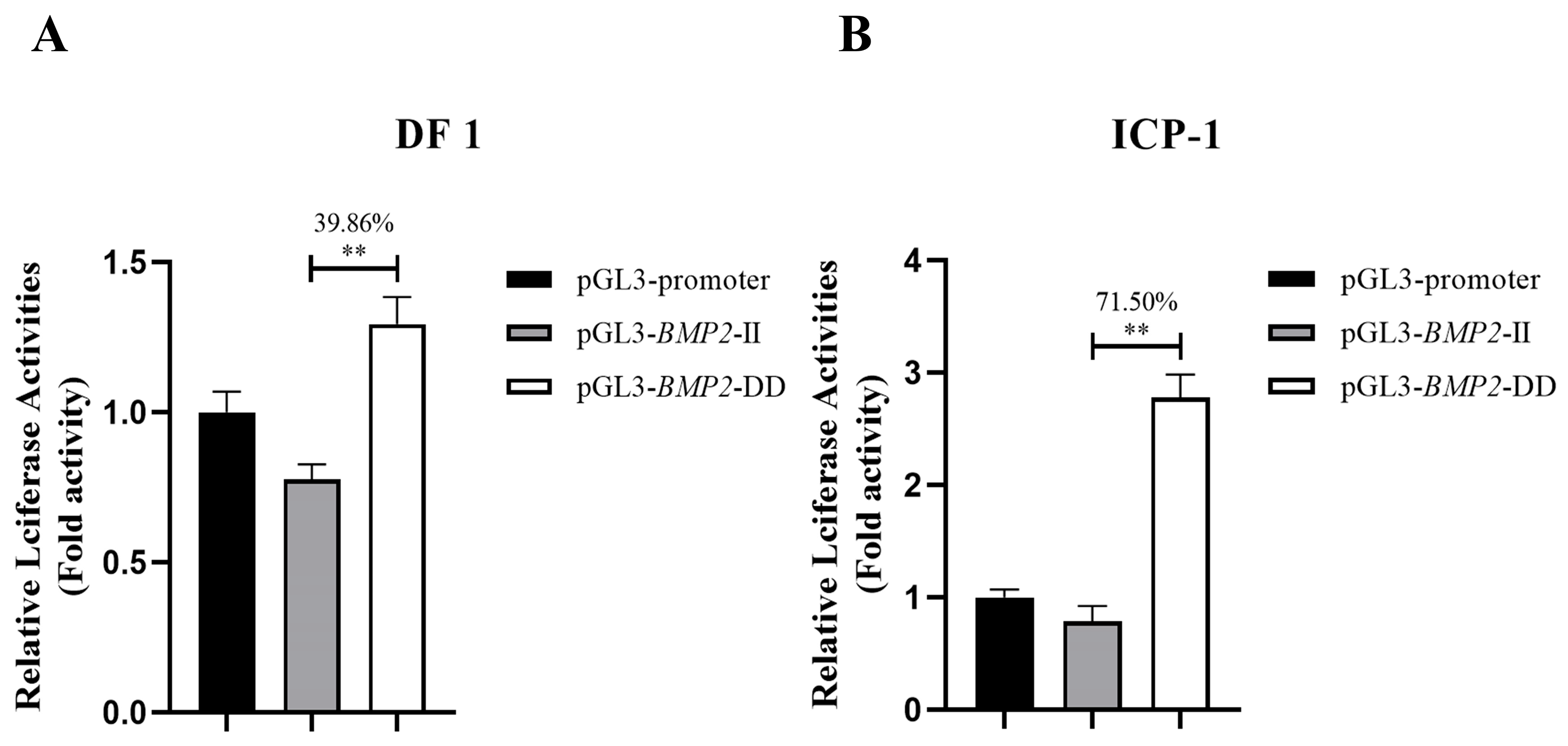
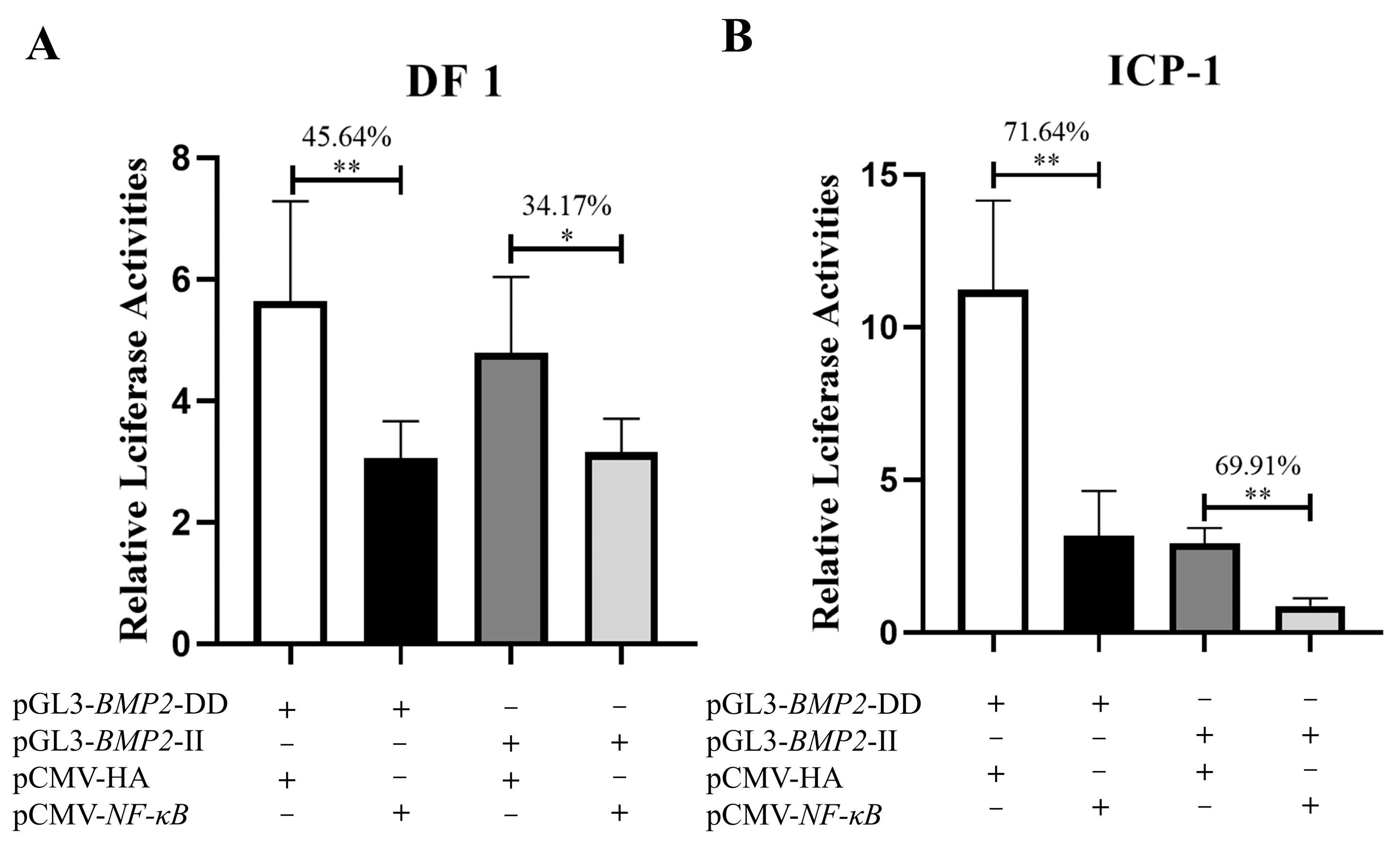
3.1. Luciferase Reporter Assay
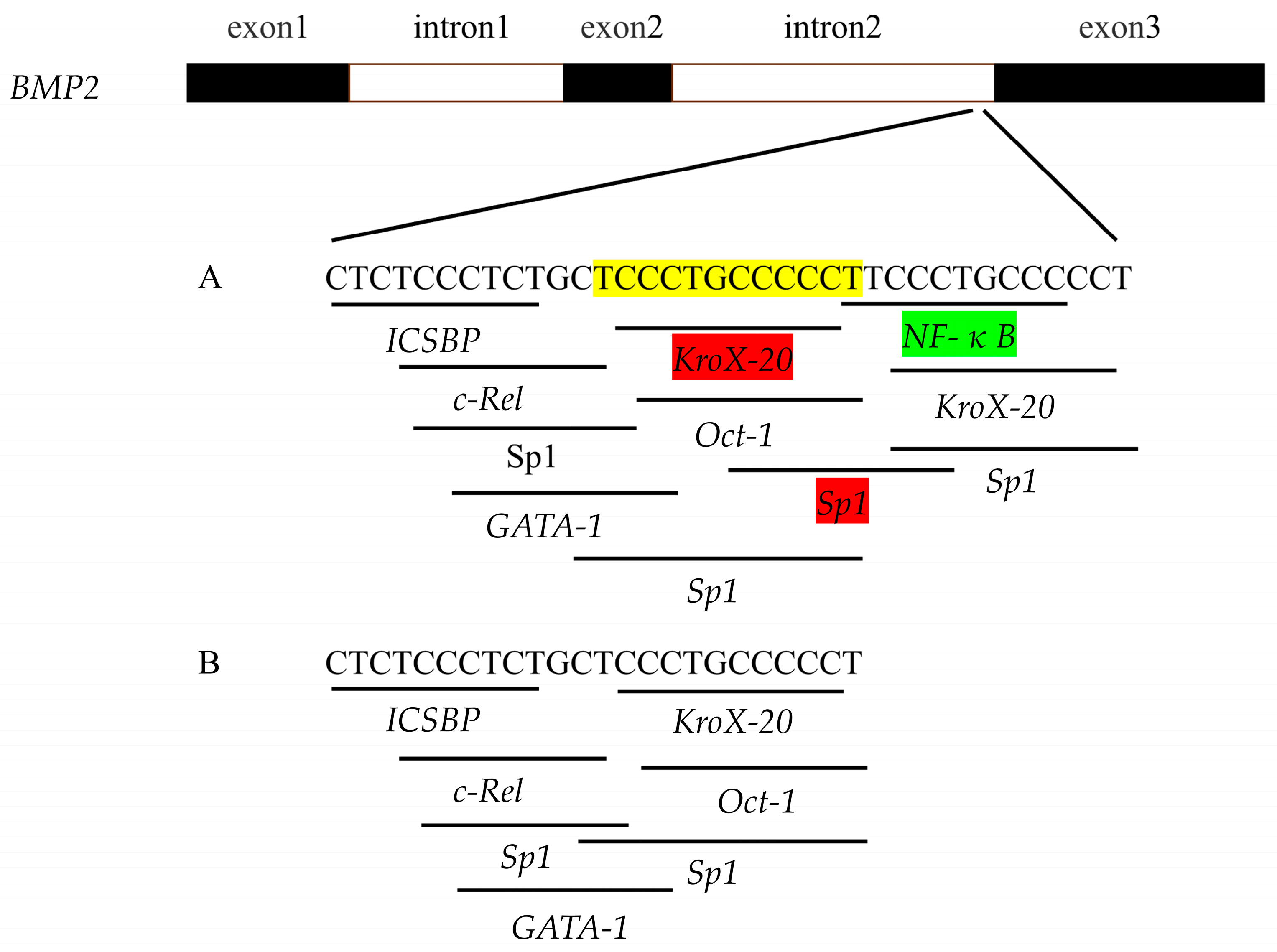
3.2. Bioinformatics Analysis
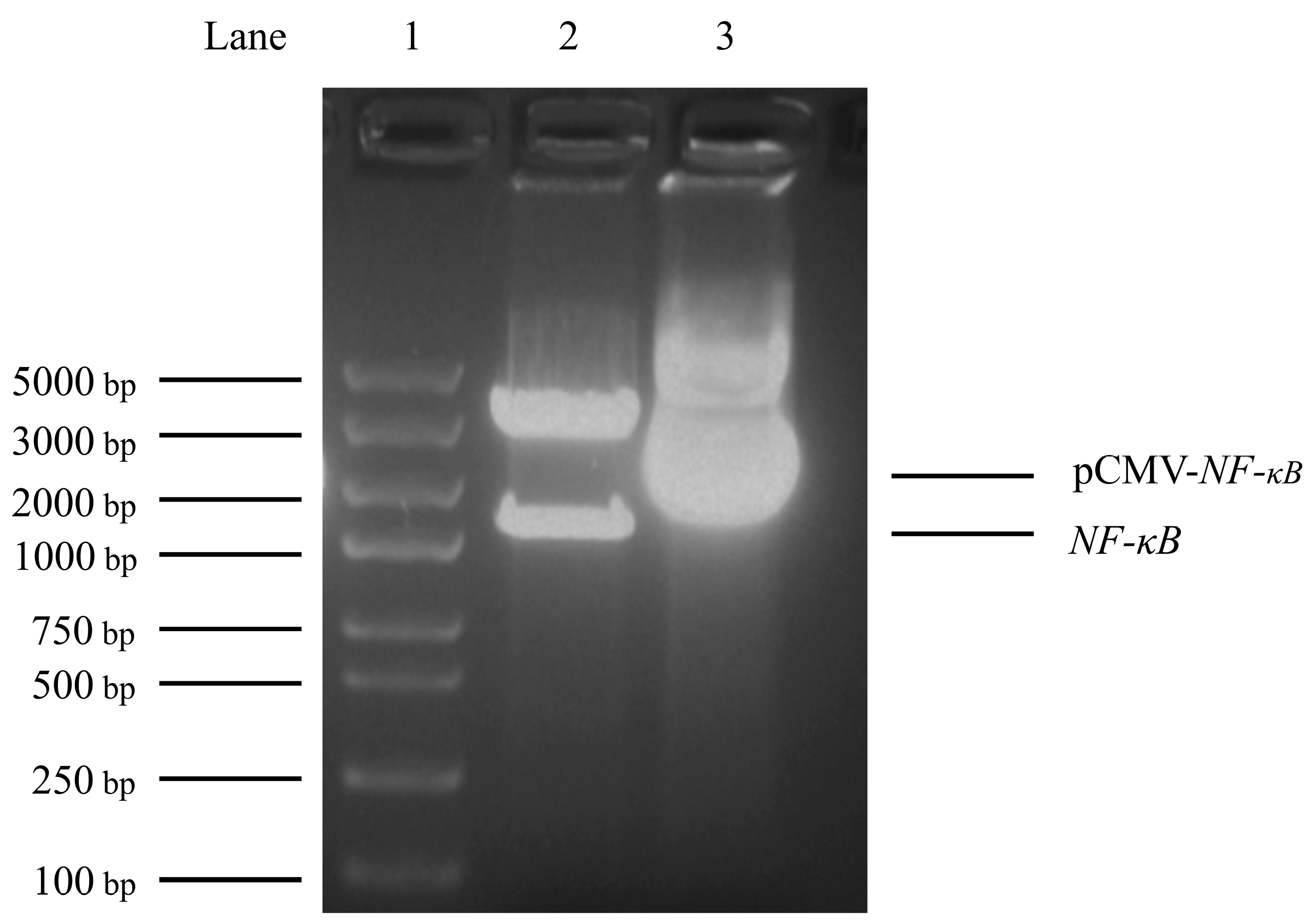
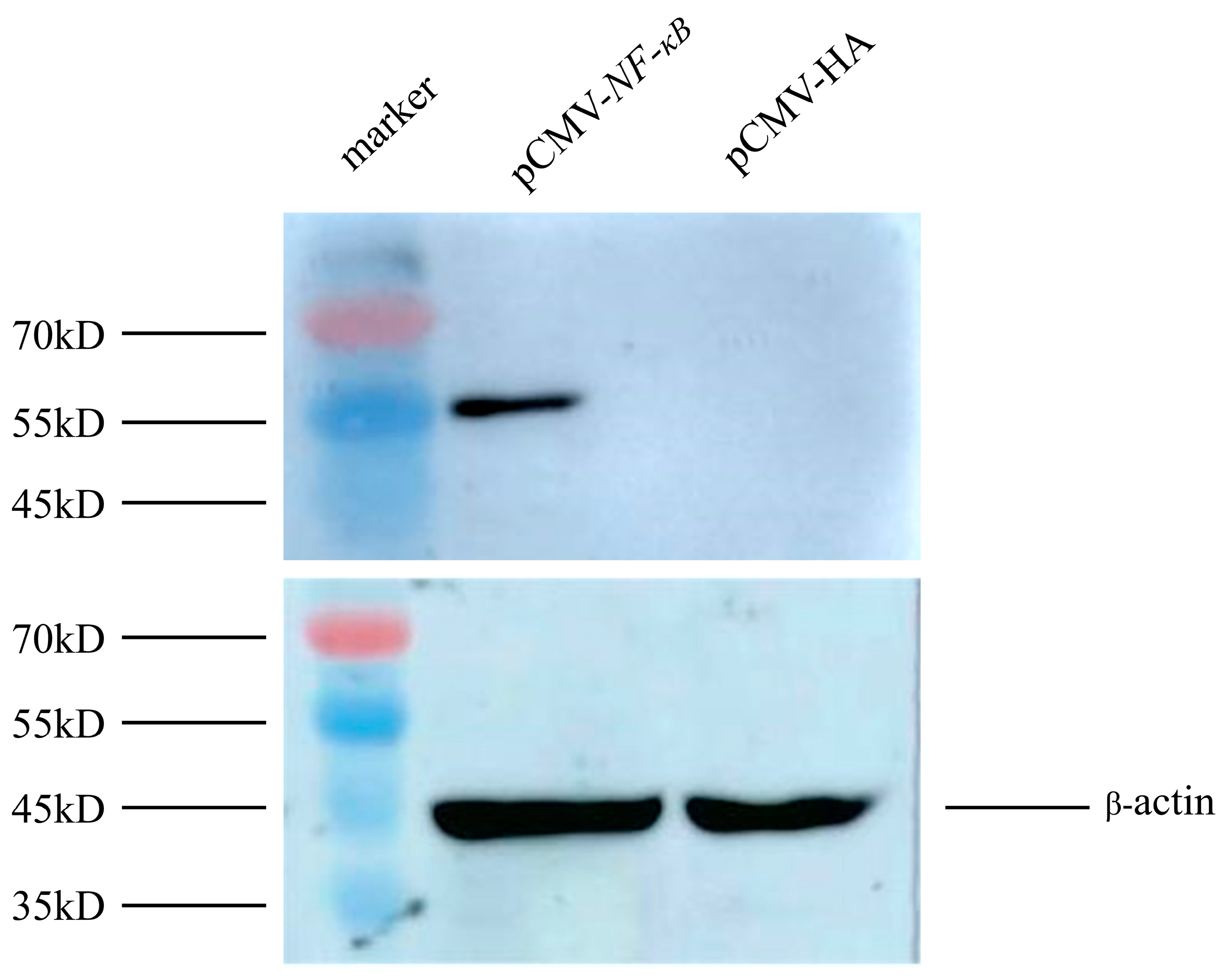
3.3. Construction of Transcription factor NF-κB Eukaryotic Expression Vector
3.4. Transcription Factor NF-κB Regulates the Expression of BMP2 Gene via the 12-bp InDel Variation
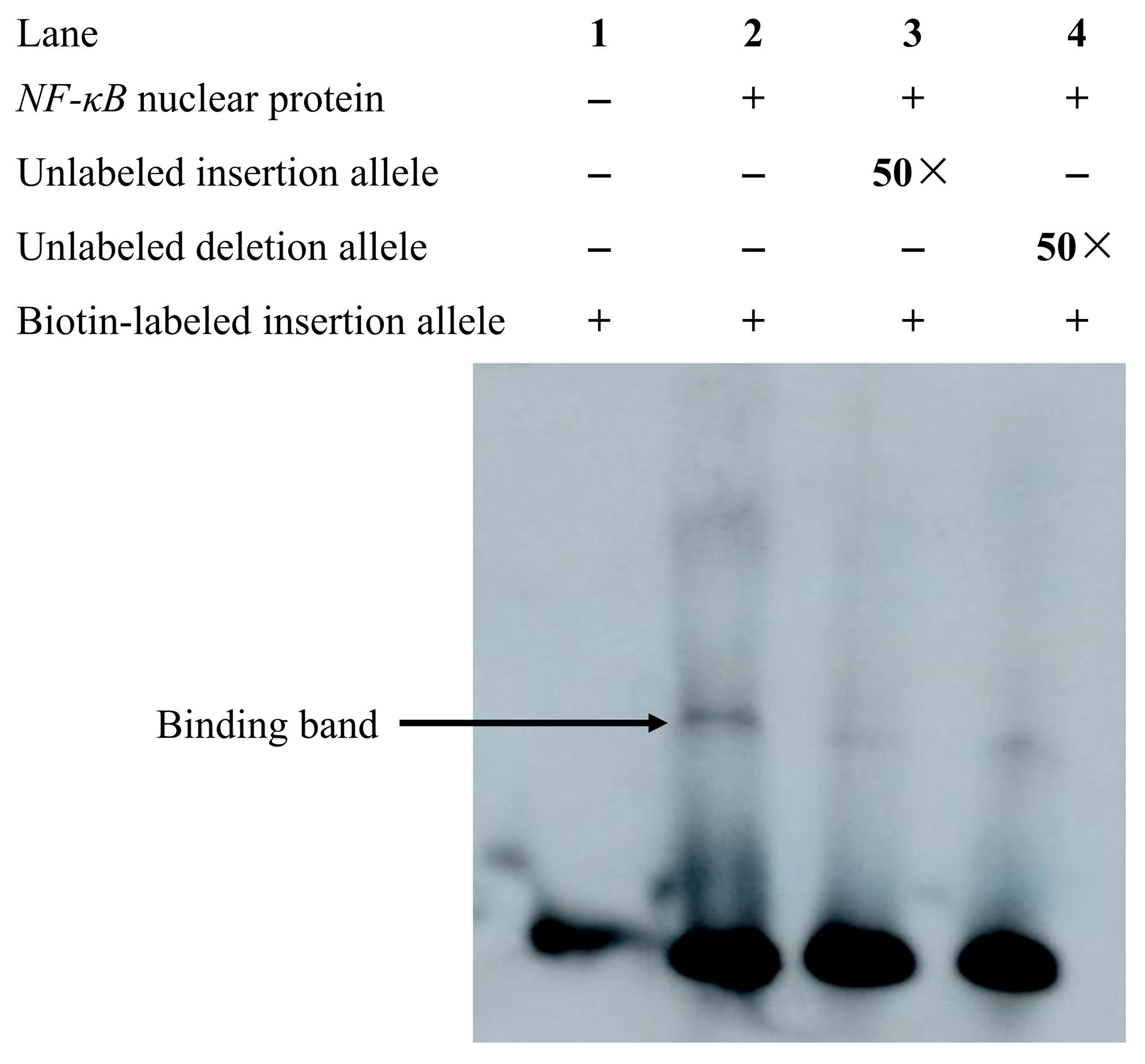
3.5. Electrophoretic Mobility Shift Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Chen, L.; Zhang, T.; Zhang, S.; Huang, J.; Zhang, G.; Xie, K.; Wang, J.; Wu, H.; Dai, G. Identification of long non-coding RNA-associated competing endogenous RNA network in the differentiation of chicken preadipocytes. Genes 2019, 10, 795. [Google Scholar] [CrossRef]
- Moreira, G.C.M.; Boschiero, C.; Cesar, A.S.M.; Reecy, J.M.; Godoy, T.F.; Pértille, F.; Ledur, M.C.; Moura, A.S.A.M.T.; Garrick, D.J.; Coutinho, L.L. Integration of genome wide association studies and whole genome sequencing provides novel insights into fat deposition in chicken. Sci. Rep. 2018, 8, 16222. [Google Scholar] [CrossRef]
- Moreira, G.C.M.; Boschiero, C.; Cesar, A.S.M.; Reecy, J.M.; Godoy, T.F.; Trevisoli, P.A.; Cantão, M.E.; Ledur, M.C.; Ibelli, A.M.G.; Peixoto, J.D.O.; et al. A genome-wide association study reveals novel genomic regions and positional candidate cenes for fat deposition in broiler chickens. BMC Genom. 2018, 19, 374. [Google Scholar]
- Zhang, X.Y.; Wu, M.Q.; Wang, S.Z.; Zhang, H.; Du, Z.Q.; Li, Y.M.; Cao, Z.P.; Luan, P.; Leng, L.; Li, H. Genetic selection on abdominal fat content alters the reproductive performance of broilers. Animal 2018, 12, 1232–1241. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, Y.; Liu, Z.; Guo, X.; Deng, Y.; Ouyang, Q.; Liu, H.; Hu, S.; Hu, B.; Li, L.; et al. Effects of rearing systems on production performance, antioxidant capacity and immune status of meat ducks at different ages. Animal 2021, 15, 100199. [Google Scholar] [CrossRef]
- Milićević, D.; Vranić, D.; Mašić, Z.; Parunović, N.; Trbović, D.; Nedeljković-Trailović, J.; Petrović, Z. The role of total fats, saturated/unsaturated fatty acids and cholesterol content in chicken meat as cardiovascular risk factors. Lipids Health Dis. 2014, 13, 42. [Google Scholar]
- Chen, C.; Su, Z.Y.; Li, Y.M.; Luan, P.; Wang, S.Z.; Zhang, H.; Xiao, F.; Guo, H.S.; Cao, Z.P.; Li, H.; et al. Estimation of the genetic parameters of traits relevant to feed efficiency: Result from broiler lines divergent for high or low abdominal fat content. Poult. Sci. 2021, 100, 461–466. [Google Scholar] [CrossRef]
- Willems, O.W.; Miller, S.P.; Wood, B.J. Aspects of selection for feed efficiency in meat producing poultry. Worlds Poult. Sci. J. 2013, 69, 77–88. [Google Scholar] [CrossRef]
- Fu, Q.H.; Wang, P.; Zhang, Y.R.; Wu, T.; Huang, J.P.; Song, Z.Y. Effects of dietary inclusion of asiaticoside on growth performance, lipid metabolism, and gut microbiota in Yellow-Feathered Chickens. Animals 2023, 13, 2653. [Google Scholar]
- Chen, Y.; Akhtar, M.; Ma, Z.Y.; Hu, T.W.; Liu, Q.Y.; Pan, H.; Zhang, X.L.; Nafady, A.A.; Ansari, A.R.; Abdel-Kafy, E.-S.M.; et al. Chicken cecal microbiota reduces abdominal fat deposition by regulating fat metabolism. NPJ Biofilms Microbiomes 2023, 9, 28. [Google Scholar]
- Zerehdaran, S.; Vereijken, A.L.J.; Van Arendonk, J.A.M.; Van Der Waaijt, E.H. Estimation of genetic parameters for fat deposition and carcass traits in broilers. Poult. Sci. 2004, 83, 521–525. [Google Scholar] [CrossRef] [PubMed]
- Urist, M.R. Bone: Formation by autoinduction. Science 1965, 150, 893–899. [Google Scholar] [CrossRef] [PubMed]
- Ahrens, M.; Ankenbauer, T.; Schröder, D.; Hollnagel, A.; Mayer, H.; Gross, G. Expression of human Bone morphogenetic proteins-2 or -4 in murine mesenchymal progenitor C3H10T½ cells induces differentiation into distinct mesenchymal cell lineages. DNA Cell Biol. 1993, 12, 871–880. [Google Scholar] [PubMed]
- Devaney, J.M.; Tosi, L.L.; Fritz, D.T.; Gordish-Dressman, H.A.; Jiang, S.; Orkunoglu-Suer, F.E.; Gordon, A.H.; Harmon, B.T.; Thompson, P.D.; Clarkson, P.M.; et al. Differences in fat and muscle mass associated with a functional human polymorphism in a post-transcriptional BMP2 gene regulatory element. J. Cell. Biochem. 2009, 107, 1073–1082. [Google Scholar]
- Hata, K.; Nishimura, R.; Ikeda, F.; Yamashita, K.; Matsubara, T.; Nokubi, T.; Yoneda, T. Differential roles of Smad1 and P38 kinase in regulation of peroxisome proliferator-activating receptor γ during bone morphogenetic protein 2-induced adipogenesis. Mol. Biol. Cell. 2003, 14, 545–555. [Google Scholar]
- Denton, N.F.; Eghleilib, M.; Al-Sharifi, S.; Todorčević, M.; Neville, M.J.; Loh, N.; Drakesmith, A.; Karpe, F.; Pinnick, K.E. Bone morphogenetic protein 2 is a depot-specific regulator of human adipogenesis. Int. J. Obes. 2019, 43, 2458–2468. [Google Scholar]
- Wang, S.B.; Zhou, G.X.; Shu, G.; Wang, L.N.; Zhu, X.T.; Gao, P.; Xi, Q.Y.; Zhang, Y.L.; Yuan, L.; Jiang, Q.Y. Glucose utilization, lipid metabolism and BMP-Smad signaling pathway of porcine intramuscular preadipocytes compared with subcutaneous preadipocytes. Cell. Physiol. Biochem. 2013, 31, 981–996. [Google Scholar] [CrossRef]
- Li, S.; Chen, C.Z.; Chai, M.L.; Wang, J.W.; Yuan, B.; Gao, Y.; Jiang, H.; Zhang, J.B. Identification and analysis of lncRNAs by whole-transcriptome sequencing in porcine preadipocytes induced by BMP2. Cytogenet. Genome Res. 2019, 158, 133–144. [Google Scholar] [CrossRef]
- Yuan, Z.; Liu, E.; Liu, Z.; Kijas, J.W.; Zhu, C.; Hu, S.; Ma, X.; Zhang, L.; Du, L.; Wang, H.; et al. Selection signature analysis reveals genes associated with tail type in Chinese indigenous sheep. Anim. Genet. 2017, 48, 55–66. [Google Scholar]
- Wang, X.L.; Zhou, G.X.; Xu, X.C.; Geng, R.Q.; Zhou, J.P.; Yang, Y.X.; Yang, Z.X.; Chen, Y.L. Transcriptome profile analysis of adipose tissues from fat and short-tailed sheep. Gene 2014, 549, 252–257. [Google Scholar]
- Lu, Z.K.; Liu, J.B.; Han, J.L.; Yang, B.H. Association between BMP2 functional polymorphisms and sheep tail type. Animals 2020, 10, 739. [Google Scholar] [PubMed]
- Pan, Z.Y.; Li, S.D.; Liu, Q.Y.; Wang, Z.; Zhou, Z.K.; Di, R.; An, X.J.; Miao, B.P.; Wang, X.Y.; Hu, W.P.; et al. Rapid evolution of a retro-transposable hotspot of ovine genome underlies the alteration of BMP2 expression and development of fat tails. BMC Genom. 2019, 20, 261. [Google Scholar]
- Yang, L.; Hao, W.G.; Wang, H.Z.; Ren, W.P.; Yan, P.S.; Wei, S.J. BMP2 increases hyperplasia and hypertrophy of bovine subcutaneous preadipocytes via BMP/SMAD signaling. In Vitro Cell. Dev. Biol. Anim. 2022, 58, 210–219. [Google Scholar] [PubMed]
- Yang, C.Y.; Wang, Z.X.; Song, Q.Q.; Dong, B.Q.; Bi, Y.L.; Bai, H.; Jiang, Y.; Chang, G.B.; Chen, G.H. Transcriptome sequencing to identify important genes and lncRNAs regulating abdominal fat deposition in ducks. Animals 2022, 12, 1256. [Google Scholar] [PubMed]
- He, L.Z.; Leng, L.; Li, H. Tissue expression characteristics of BMP2 gene and its difference in adipose tissue of different chicken strains, Safe and quality poultry production. In Proceedings of the 15th National Poultry Symposium, Guangzhou, China, 1 November 2011; pp. 162–165. [Google Scholar]
- Leng, L.; Wang, Q.G.; Wang, S.Z.; Li, H. Tissue expression of BMP-2 gene in chicken and its correlation with body fat and bone traits, Advances in animal genetics and breeding in China. In Proceedings of the 15th National Symposium on Animal Genetics and Breeding, Xianyang, China, 10 October 2009; p. 433. [Google Scholar]
- Guo, L.; Sun, B.; Shang, Z.; Leng, L.; Wang, Y.; Wang, N.; Li, H. Comparison of adipose tissue cellularity in chicken lines divergently selected for fatness. Poult. Sci. 2011, 90, 2024–2034. [Google Scholar] [PubMed]
- Leng, L.; Wang, S.; Li, Z.; Wang, Q.; Li, H. A polymorphism in the 3′-Flanking Region of insulin-like growth factor binding protein 2 gene associated with abdominal fat in chickens. Poult. Sci. 2009, 88, 938–942. [Google Scholar] [CrossRef]
- Mannstadt, M.; Bertrand, G.; Muresan, M.; Weryha, G.; Leheup, B.; Pulusani, S.R.; Grandchamp, B.; Jüppner, H.; Silve, C. Dominant-negative GCMB mutations cause an autosomal dominant form of hypoparathyroidism. J. Clin. Endocrinol. Metab. 2008, 93, 3568–3576. [Google Scholar]
- Wang, Y.S.; Ouyang, W.O.; Pan, Q.X.; Wang, X.L.; Xia, X.X.; Bi, Z.W.; Wang, Y.Q.; Wang, X.M. Overexpression of microRNA gga-miR-21 in chicken fibroblasts suppresses replication of infectious bursal disease virus through inhibiting VP1 translation. Antiviral Res. 2013, 100, 196–201. [Google Scholar] [CrossRef]
- Wang, W.; Zhang, T.M.; Wu, C.Y.; Wang, S.S.; Wang, Y.X.; Li, H.; Wang, N. Immortalization of chicken preadipocytes by retroviral transduction of chicken TERT and TR. PLoS ONE 2017, 12, e0177348. [Google Scholar]
- Sherf, B.A.; Navarro, S.L.; Hannah, R.R.; Wood, K.V. Dual-luciferase reporter assay: An advanced co-reporter technology integrating firefly and Renilla luciferase assays. Promega Notes 1996, 57, 2–8. [Google Scholar]
- Cheng, B.H.; Zhang, H.; Liu, C.; Chen, X.; Chen, Y.F.; Sun, Y.H.; Leng, L.; Li, Y.M.; Luan, P.; Li, H. Functional intronic variant in the retinoblastoma 1 gene underlies broiler chicken adiposity by altering nuclear factor-kB and SRY-related HMG box protein 2 binding sites. J. Agric. Food Chem. 2019, 67, 9727–9737. [Google Scholar] [CrossRef]
- Tang, T.Y.; Zhang, J.; Yin, J.; Staszkiewicz, J.; Gawronska-Kozak, B.; Jung, D.Y.; Ko, H.J.; Ong, H.; Kim, J.K.; Mynatt, R.; et al. Uncoupling of inflammation and insulin resistance by NF-kappaB in transgenic mice through elevated energy expenditure. J. Biol. Chem. 2010, 285, 4637–4644. [Google Scholar] [CrossRef]
- Ruan, H.; Pownall, H.J.; Lodish, H.F. Troglitazone antagonizes tumor necrosis factor-alpha-induced reprogramming of adipocyte gene expression by inhibiting the transcriptional regulatory functions of NF-kappaB. J. Biol. Chem. 2003, 278, 28181–28192. [Google Scholar] [CrossRef]
- Guilherme, A.; Virbasius, J.V.; Puri, V.; Czech, M.P. Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 367–377. [Google Scholar] [CrossRef]
- Chen, Z.; Torrens, J.I.; Anand, A.; Spiegelman, B.M.; Friedman, J.M. Krox20 stimulates adipogenesis via C/EBPβ-dependent and -independent mechanisms. Cell Metab. 2005, 1, 93–106. [Google Scholar] [CrossRef]
- Gonzalez, F.J. Getting fat: Two new players in molecular adipogenesis. Cell Metab. 2005, 1, 85–86. [Google Scholar] [CrossRef]
- Schreiber, R.; Xie, H.; Schweiger, M. Of mice and men: The physiological role of adipose triglyceride lipase (ATGL). Biochim. Biophys. Acta Mol. Cell Biol. Lipids 2019, 1864, 880–899. [Google Scholar]
- Chen, S.; Hu, Z.G.; He, H.; Liu, X.L. Fatty acid elongase7 is regulated via SP1 and is involved in lipid accumulation in bovine mammary epithelial cells. J. Cell. Physiol. 2018, 233, 4715–4725. [Google Scholar] [CrossRef]
- Zhu, J.J.; Sun, Y.T.; Luo, J.; Wu, M.; Li, J.H.; Cao, Y.H. Specificity protein 1 regulates gene expression related to fatty acid metabolism in goat mammary epithelial cells. Int. J. Mass. Spectrom. 2015, 16, 1806–1820. [Google Scholar] [CrossRef]
- Katagiri, T. Bone morphogenetic protein-2 converts the differentiation pathway of C2C12 myoblasts into the osteoblast lineage. J. Cell Biol. 1994, 127, 1755–1766. [Google Scholar] [CrossRef]
- Zhou, N.; Li, Q.; Lin, X.; Hu, N.; Liao, J.-Y.; Lin, L.B.; Zhao, C.; Hu, Z.M.; Liang, X.; Xu, W.; et al. BMP2 induces chondrogenic differentiation, osteogenic differentiation and endochondral ossification in stem cells. Cell Tissue Res. 2016, 366, 101–111. [Google Scholar] [CrossRef]
- Salazar, V.S.; Gamer, L.W.; Rosen, V. BMP signalling in skeletal development, disease and repair. Nat. Rev. Endocrinol. 2016, 12, 203–221. [Google Scholar]
- Guiu-Jurado, E.; Unthan, M.; Böhler, N.; Kern, M.; Landgraf, K.; Dietrich, A.; Schleinitz, D.; Ruschke, K.; Klöting, N.; Faßhauer, M.; et al. Bone morphogenetic protein 2 (BMP2) may contribute to partition of energy storage into visceral and subcutaneous fat depots. Obesity 2016, 24, 2092–2100. [Google Scholar]
- Sottile, V.; Seuwen, K. Bone morphogenetic protein-2 stimulates adipogenic differentiation of mesenchymal precursor cells in synergy with BRL 49653 (rosiglitazone). FEBS Lett. 2000, 475, 201–204. [Google Scholar]
- Katagiri, T.; Akiyama, S.; Namiki, M.; Komaki, M.; Yamaguchi, A.; Rosen, V.; Wozney, J.M.; Fujisawa-Sehara, A.; Suda, T. Bone morphogenetic protein-2 inhibits terminal differentiation of myogenic cells by suppressing the transcriptional activity of MyoD and myogenin. Exp. Cell Res. 1997, 230, 342–351. [Google Scholar]
- Lv, Y.; Gao, C.W.; Liu, B.; Wang, H.Y.; Wang, H.P. BMP-2 combined with salvianolic acid B promotes cardiomyocyte differentiation of rat bone marrow mesenchymal stem cells. Kaohsiung J. Med. Sci. 2017, 33, 477–485. [Google Scholar]
- Miao, Y.X.; Zhao, Y.X.; Wan, S.Q.; Mei, Q.S.; Wang, H.; Fu, C.K.; Li, X.Y.; Zhao, S.H.; Xu, X.W.; Xiang, T. Integrated analysis of genome-wide association studies and 3D epigenomic characteristics reveal the BMP2 gene regulating loin muscle depth in Yorkshire pigs. PLoS Genet. 2023, 19, e1010820. [Google Scholar]
- Li, Y.; Wang, L.; Zhou, J.W.; Li, F.G. Transcription factor organic cation transporter 1 (OCT-1) affects the expression of porcine Klotho (KL) gene. PeerJ 2016, 4, e2186. [Google Scholar]
- Fuxman Bass, J.I.; Tamburino, A.M.; Mori, A.; Beittel, N.; Weirauch, M.T.; Reece-Hoyes, J.S.; Walhout, A.J.M. Transcription factor binding to Caenorhabditis elegans first introns reveals lack of redundancy with gene promoters. Nucleic Acids Res. 2014, 42, 153–162. [Google Scholar]
- Hughes, T.R. Introduction to “a handbook of transcription factors”. Subcell. Biochem. 2011, 52, 1–6. [Google Scholar]
- Meyer, K.B.; Maia, A.-T.; O’Reilly, M.; Teschendorff, A.E.; Chin, S.-F.; Caldas, C.; Ponder, B.A.J. Allele-specific up-regulation of FGFR2 increases susceptibility to breast cancer. PLOS Biol. 2008, 6, e108. [Google Scholar] [CrossRef]
- Van Laere, A.-S.; Nguyen, M.; Braunschweig, M.; Nezer, C.; Collette, C.; Moreau, L.; Archibald, A.L.; Haley, C.S.; Buys, N.; Tally, M.; et al. A Regulatory mutation in IGF2 causes a major QTL effect on muscle growth in the pig. Nature 2003, 425, 832–836. [Google Scholar] [PubMed]
- Markljung, E.; Jiang, L.; Jaffe, J.D.; Mikkelsen, T.S.; Wallerman, O.; Larhammar, M.; Zhang, X.L.; Wang, L.; Saenz-Vash, V.; Gnirke, A.; et al. ZBED6, a novel transcription factor derived from a domesticated DNA transposon regulates IGF2 expression and muscle growth. PLOS Biol. 2009, 7, e1000256. [Google Scholar]
- Zou, H.Y.; Yu, D.W.; Yao, S.; Ding, F.R.; Li, J.L.; Li, L.; Li, X.; Zhao, S.J.; Pang, Y.W.; Hao, H.S.; et al. Efficient editing of the ZBED6-binding site in intron 3 of IGF2 in a bovine model using the CRISPR/Cas9 system. Genes 2022, 13, 1132. [Google Scholar] [PubMed]
- Shield, P.W.; Papadimos, D.J.; Walsh, M.D. GATA3: A promising marker for metastatic breast carcinoma in serous effusion specimens: GATA3 staining of serous effusions. Cancer Cytopathol. 2014, 122, 307–312. [Google Scholar]
- Zhou, W.B.; Nie, X.H. Afzelin attenuates asthma phenotypes by downregulation of GATA3 in a murine model of asthma. Mol. Med. Rep. 2015, 12, 71–76. [Google Scholar] [CrossRef]
- Sun, Z.W.; Andersson, R. NF- kappaB activation and inhibition: A review. Shock. 2002, 18, 99–106. [Google Scholar]
- Hayden, M.S.; Ghosh, S. Shared principles in NF-kappaB signaling. Cell 2008, 132, 344–362. [Google Scholar]
- Tisdale, M.J. Biology of cachexia. J. Natl. Cancer Inst. 1997, 89, 1763–1773. [Google Scholar]
- Larcombe, M.R.; Hsu, S.; Polo, J.M.; Knaupp, A.S. Indirect mechanisms of transcription factor-mediated gene regulation during cell fate changes. Adv. Genet. 2022, 3, 2200015. [Google Scholar]
- Bauer, S.; Eigenmann, J.; Zhao, Y.; Fleig, J.; Hawe, J.S.; Pan, C.; Bongiovanni, D.; Wengert, S.; Ma, A.; Lusis, A.J.; et al. Identification of the transcription factor ATF3 as a direct and indirect regulator of the LDLR. Metabolites 2022, 12, 840. [Google Scholar] [PubMed]
- Jaganathan, K.; Kyriazopoulou Panagiotopoulou, S.; McRae, J.F.; Darbandi, S.F.; Knowles, D.; Li, Y.I.; Kosmicki, J.A.; Arbelaez, J.; Cui, W.; Schwartz, G.B.; et al. Predicting splicing from primary sequence with deep learning. Cell 2019, 176, 535–548.e24. [Google Scholar] [PubMed]
- Pattison, J.M.; Posternak, V.; Cole, M.D. Transcription factor KLF5 binds a cyclin E1 polymorphic intronic enhancer to confer increased bladder cancer risk. Mol. Cancer Res. 2016, 14, 1078–1086. [Google Scholar] [PubMed]
- Fabo, T.; Khavari, P. Functional characterization of human genomic variation linked to polygenic diseases. Trends Genet. 2023, 39, 462–490. [Google Scholar] [PubMed]
- French, J.D.; Edwards, S.L. The role of noncoding variants in heritable disease. Trends Genet. 2020, 36, 880–891. [Google Scholar]
- Takata, A.; Matsumoto, N.; Kato, T. Genome-wide identification of splicing QTLs in the human brain and their enrichment among schizophrenia-associated loci. Nat. Commun. 2017, 8, 14519. [Google Scholar]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| GenBank Accession No. | Primer Name | Primer Sequence (5′-3′) | Purpose |
|---|---|---|---|
| NM_001396395.1 | NF-κB-F | 5′-CCGTATCTTCAAATCATTGAACAGCC-3′ | Construction of eukaryotic expression vector |
| NF-κB-R | 5′-ATAGCCTTCTCCAGGAACAGACCATC-3′ | ||
| NM_205518.1 | β-actin-F | 5′-TGGCCATGGAGGCCCGAATTCCCAAAATGCCAACCCT-3′ | Loading control |
| β-actin-R | 5′-CCGCGGCCGCGGTACCTCGAGACTGCCCAGAAAGTTGTG-3′ |
| Transcription Factor | Probes | Probe Sequence (5′-3′) |
|---|---|---|
| NF-κB | BMP2-DD-F | CTCCCTCTGCTCCCTGCCCCCT |
| BMP2-DD-R | AGGGGGCAGGGAGCAGAGGGAG | |
| BMP2-II-F | CTCCCTCTGCTCCCTGCCCCCTTCCCTGCCCCCT | |
| BMP2-II-R | AGGGGGCAGGGAAGGGGGCAGGGAGCAGAGGGAG | |
| BMP2-B-II-F | CTCCCTCTGCTCCCTGCCCCCTTCCCTGCCCCCT | |
| BMP2-B-II-R | AGGGGGCAGGGAAGGGGGCAGGGAGCAGAGGGAG |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yuan, M.; Liu, X.; Wang, M.; Li, Z.; Li, H.; Leng, L.; Wang, S. A Functional Variant Alters the Binding of Bone morphogenetic protein 2 to the Transcription Factor NF-κB to Regulate Bone morphogenetic protein 2 Gene Expression and Chicken Abdominal Fat Deposition. Animals 2023, 13, 3401. https://doi.org/10.3390/ani13213401
Yuan M, Liu X, Wang M, Li Z, Li H, Leng L, Wang S. A Functional Variant Alters the Binding of Bone morphogenetic protein 2 to the Transcription Factor NF-κB to Regulate Bone morphogenetic protein 2 Gene Expression and Chicken Abdominal Fat Deposition. Animals. 2023; 13(21):3401. https://doi.org/10.3390/ani13213401
Chicago/Turabian StyleYuan, Meng, Xin Liu, Mengdie Wang, Ziwei Li, Hui Li, Li Leng, and Shouzhi Wang. 2023. "A Functional Variant Alters the Binding of Bone morphogenetic protein 2 to the Transcription Factor NF-κB to Regulate Bone morphogenetic protein 2 Gene Expression and Chicken Abdominal Fat Deposition" Animals 13, no. 21: 3401. https://doi.org/10.3390/ani13213401
APA StyleYuan, M., Liu, X., Wang, M., Li, Z., Li, H., Leng, L., & Wang, S. (2023). A Functional Variant Alters the Binding of Bone morphogenetic protein 2 to the Transcription Factor NF-κB to Regulate Bone morphogenetic protein 2 Gene Expression and Chicken Abdominal Fat Deposition. Animals, 13(21), 3401. https://doi.org/10.3390/ani13213401