
The Effect of a Synthetic Scent on Cheetah Behaviour



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Approval
2.2. Preparation of the Synthetic Scent
2.3. Sensory Analysis
2.4. Preliminary Tests
2.5. Experimental Design, Location and Animals
2.6. Oestrus Behaviour Observations
2.7. Statistical Analysis
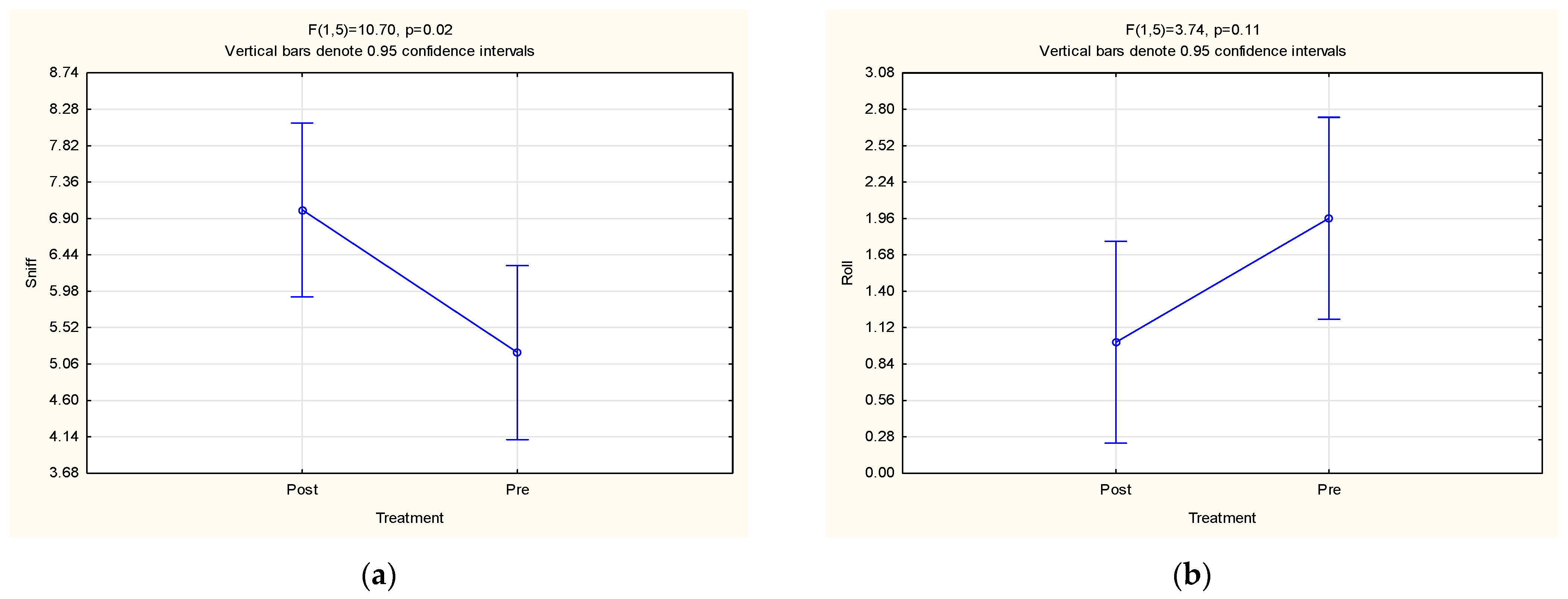
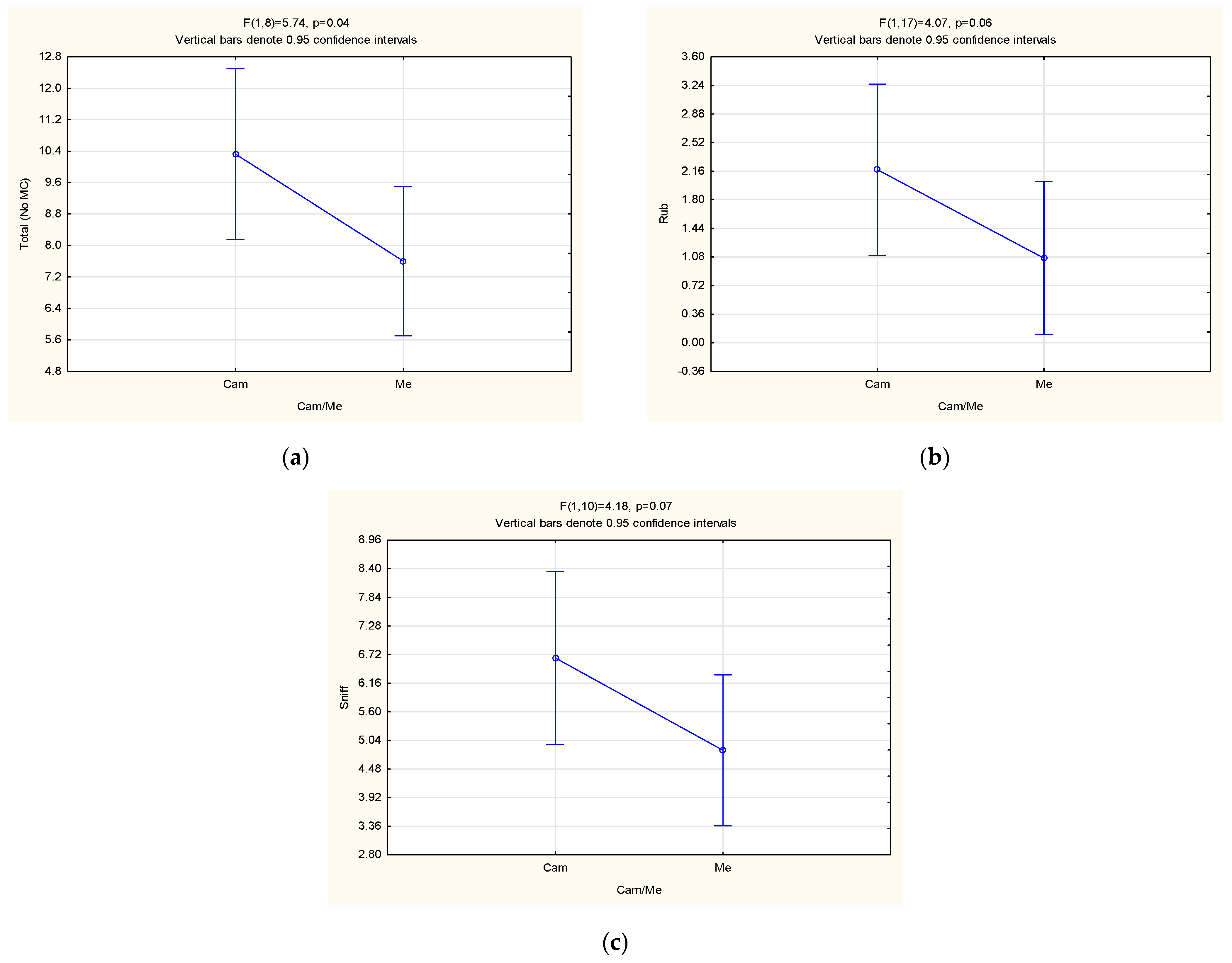
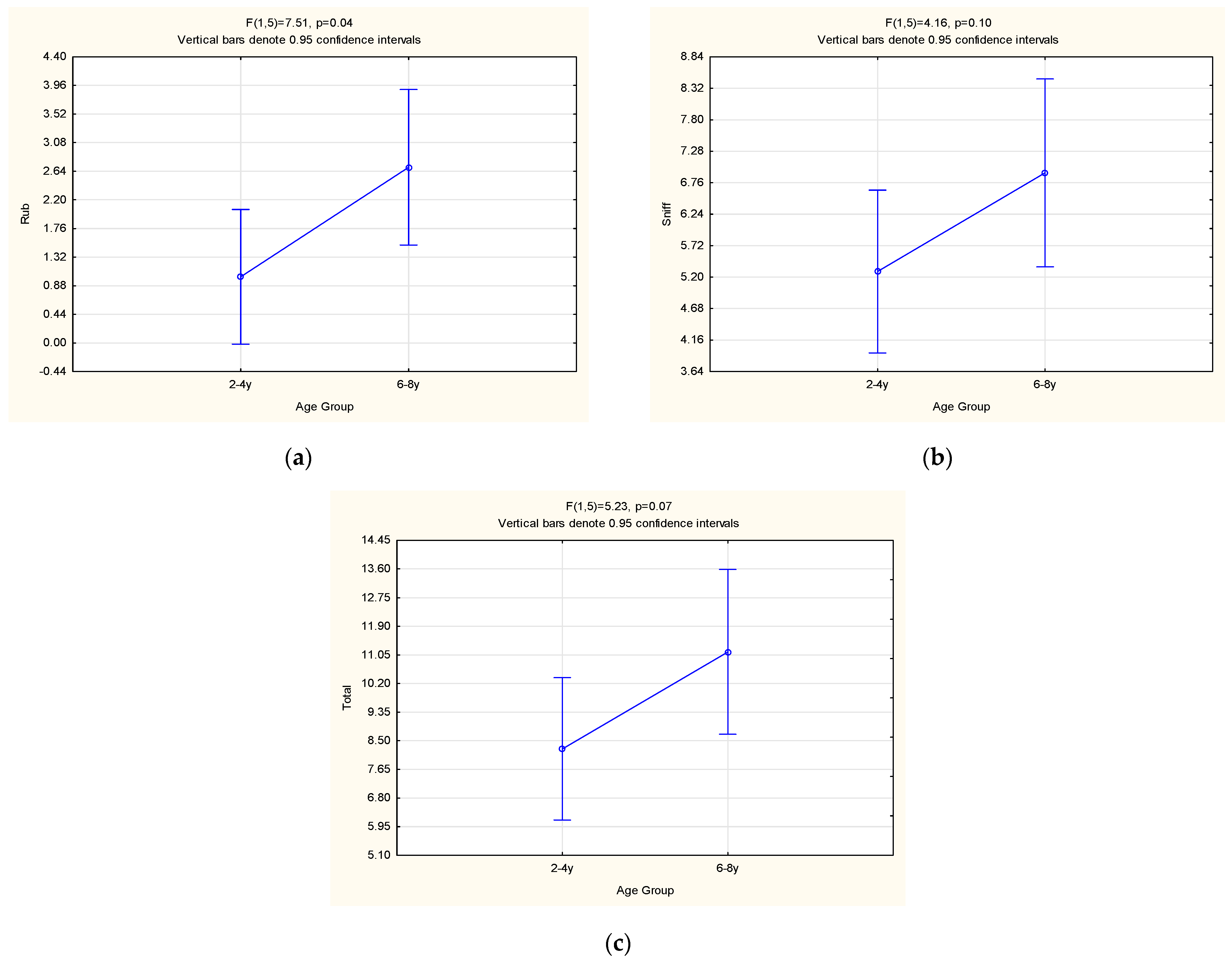
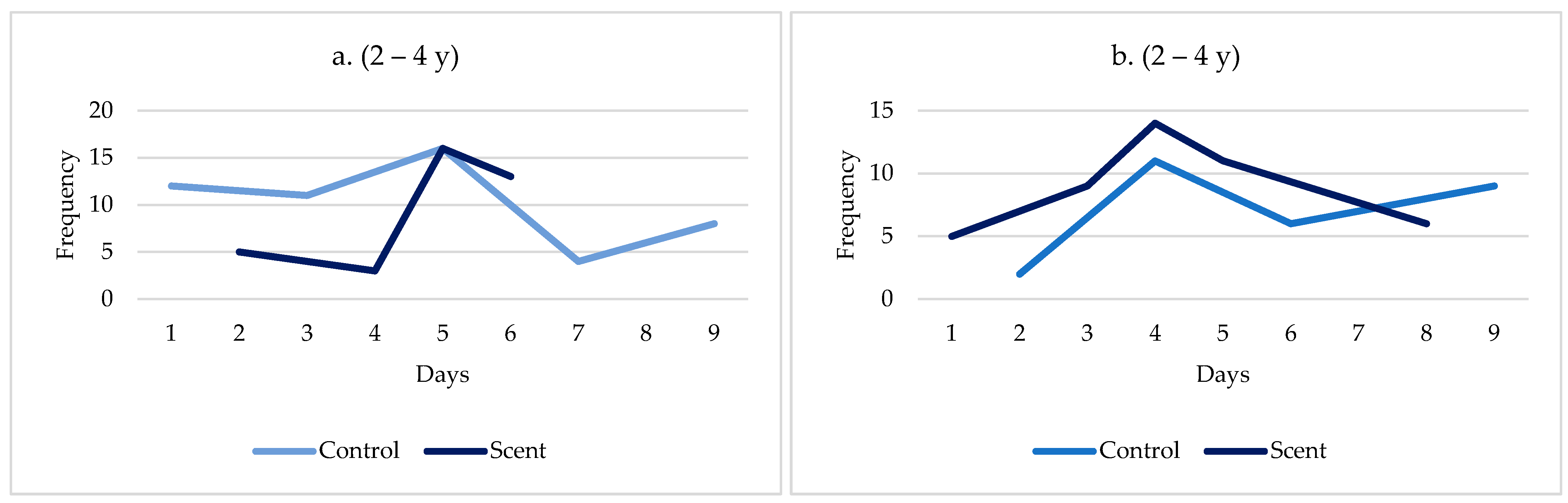
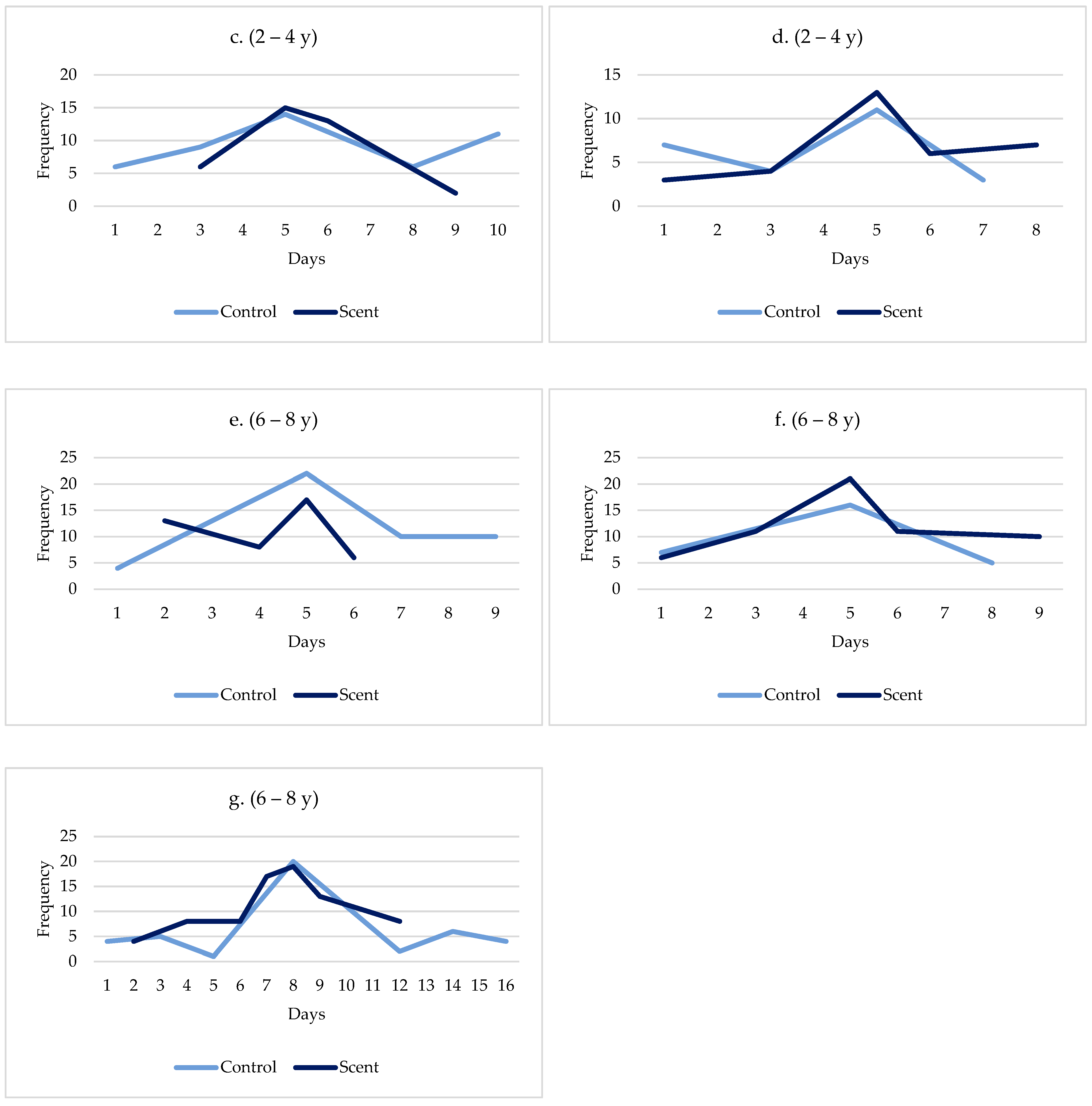
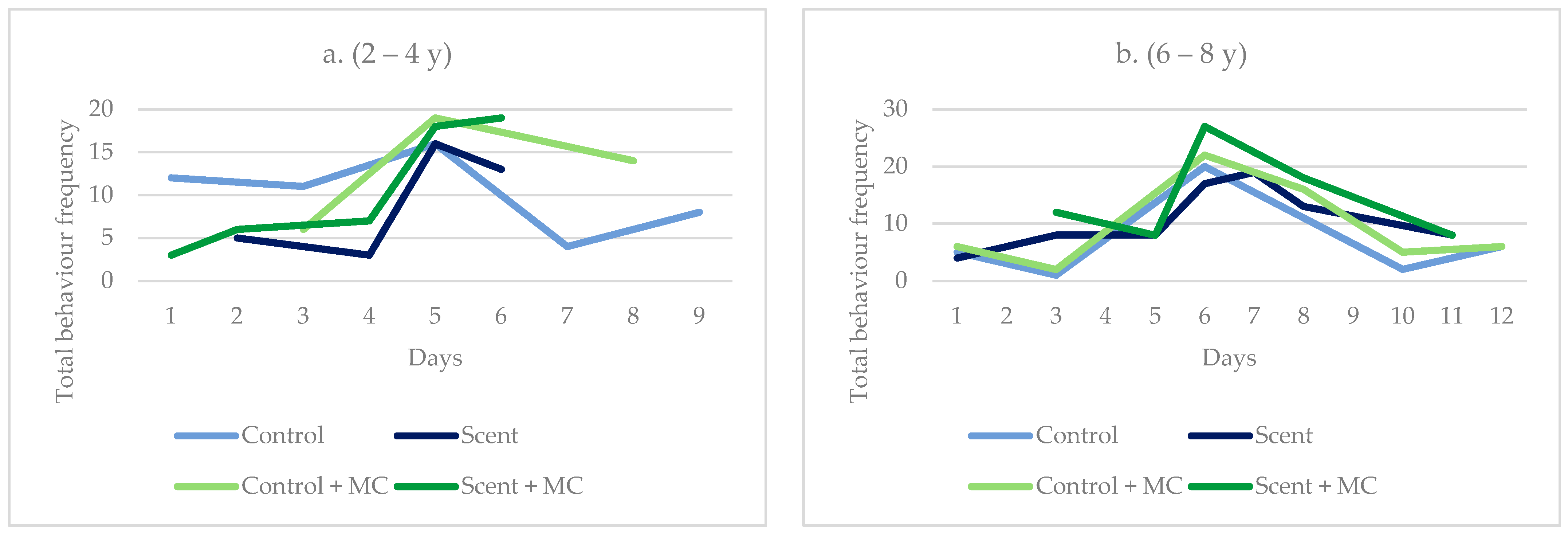
3. Results and Discussion
3.1. Sensory Analysis of the Synthetic Scent
3.2. Immediate Reactions Observed to the Components of the Synthetic Scent
3.3. Behavioural Observations
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A



References
- Cheetah Outreach. Cheetah Information. 2018. Available online: http://www.cheetah.co.za/pdf/Cheetah%20Information.doc (accessed on 1 November 2021).
- Durant, S.M.; Groom, R.; Ipavec, A.; Mitchell, N.; Khalatbari, L. Acinonyx Jubatus. The IUCN Red List of Threatened Species 2022: E.T219A124366642. IUCN Red List of Threat. Species. Available online: https://www.iucnredlist.org/species/219/50649567 (accessed on 1 November 2021).
- Koester, D.C. The Effect of Environment on the Reproductive Potential of Ex Situ Cheetahs (Acinonyx jubatus). Ph.D. Thesis, George Mason University, Fairfax, VA, USA, 2014. [Google Scholar]
- Terio, K.A.; Marker, L.; Overstrom, E.W.; Brown, J.L. Analysis of Ovarian and Adrenal Activity in Namibian Cheetahs. S. Afr. J. Wildl. Res. 2003, 33, 71–78. [Google Scholar]
- Brown, J.L.; Wildt, D.E.; Wielebnowski, N.; Goodrowe, K.L.; Graham, L.H.; Wells, S.; Howard, J.G. Reproductive activity in captive female cheetahs (Acinonyx jubatus) assessed by faecal steroids. Reproduction 1996, 106, 337–346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wielebnowski, N.; Brown, J.L. Behavioral correlates of physiological estrus in cheetahs. Zoo Biol. 1998, 17, 193–209. [Google Scholar] [CrossRef]
- Wielebnowski, N.C.; Ziegler, K.; Wildt, D.E.; Lukas, J.; Brown, J.L. Impact of social management on reproductive, adrenal and behavioural activity in the cheetah (Acinonyx jubatus). Anim. Conserv. 2002, 5, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L. Female reproductive cycles of wild female felids. Anim. Reprod. Sci. 2011, 124, 155–162. [Google Scholar] [CrossRef]
- Visser, R.R.C. Chemical Communication: Chemical Characterization of Volatile Constituents of Urine of the Southern African Cheetah, Acinonyx jubatus jubatus, Using Headspace Sampling and GC-MS. Ph.D. Thesis, Stellenbosch University, Stellen-bosch, South Africa, 2002. [Google Scholar]
- Silva, L.; Antunes, A. Vomeronasal Receptors in Vertebrates and the Evolution of Pheromone Detection. Annu. Rev. Anim. Biosci. 2017, 5, 353–370. [Google Scholar] [CrossRef] [Green Version]
- Wachter, B.; Thalwitzer, S.; Hofer, H.; Lonzer, J.; Hildebrandt, T.B.; Hermes, R. Reproductive history and absence of predators are important determinants of reproductive fitness: The cheetah controversy revisited. Conserv. Lett. 2010, 4, 47–54. [Google Scholar] [CrossRef]
- Crosier, A.E.; Comizzoli, P.; Baker, T.; Davidson, A.; Munson, L.; Howard, J.; Marker, L.L.; Wildt, D.E. Increasing Age Influences Uterine Integrity, But Not Ovarian Function or Oocyte Quality, in the Cheetah (Acinonyx jubatus). Biol. Reprod. 2011, 85, 243–253. [Google Scholar] [CrossRef] [Green Version]
- Penfold, L.M.; Powell, D.; Traylor-Holzer, K.; Asa, C.S. “Use it or lose it”: Characterization, implications, and mitigation of female infertility in captive wildlife. Zoo Biol. 2013, 33, 20–28. [Google Scholar] [CrossRef]
- Bertschinger, H.; Meltzer, D.; van Dyk, A. Captive Breeding of Cheetahs in South Africa 30Years of Data from the de Wildt Cheetah and Wildlife Centre. Reprod. Domest. Anim. 2008, 43, 66–73. [Google Scholar] [CrossRef]
- Everaert, I.; (Ann van Dyk Cheetah Centre, Pretoria, South Africa). Personal communication, 9 April 2019.
- Beekman, S.P.A.; De Wit, M.; Louwman, J.; Louwman, H. Breeding and observations on the behaviour of Cheetah Acinonyx jubatus at Wassenaar Wildlife Breeding Centre. Int. Zoo Yearb. 1997, 35, 43–50. [Google Scholar] [CrossRef]
- Andersen, K.F.; Vulpius, T. Urinary Volatile Constituents of the Lion, Panthera Leo. Chem. Senses 1999, 24, 179–189. [Google Scholar] [CrossRef] [PubMed]
- Poddar-Sarkar, M. Mammalian Semiochemicals: Chemical and Behavioural Aspects with Special Reference to Tiger. Ph.D. Thesis, University of Calcutta, Kolkata, India, 1995. [Google Scholar]
- Brahmachary, R.L.; Poddar-Sarkar, M. Fifty Years of Tiger Pheromone Research. Curr. Sci. 2015, 108, 2178–2185. [Google Scholar]
- Soso, S.B.; Koziel, J.A. Characterizing the scent and chemical composition of Panthera leo marking fluid using solid-phase microextraction and multidimensional gas chromatography–mass spectrometry-olfactometry. Sci. Rep. 2017, 7, 5137. [Google Scholar] [CrossRef] [Green Version]
- Soso, S.B.; Koziel, J.A. Analysis of Odorants in Marking Fluid of Siberian Tiger (Panthera tigris altaica) Using Simultaneous Sensory and Chemical Analysis with Headspace Solid-Phase Microextraction and Multidimensional Gas Chromatography-Mass Spectrometry-Olfactometry. Molecules 2016, 21, 834. [Google Scholar] [CrossRef] [Green Version]
- Wemmer, C.; Scow, K. Communication in the Felidae with Emphasis on Scent Marking and Contact Marking and Contact Patterns. In How Animals Communicate; Sebeok, T.A., Ed.; Indiana University Press: Bloomington, IN, USA, 1977; pp. 749–766. [Google Scholar]
- Bland, K.P. Tom-cat odour and other pheromones in feline reproduction. Vet-. Res. Commun. 1979, 3, 125–136. [Google Scholar] [CrossRef]
- Verberne, G.; de Boer, J. Chemocommunication among Domestic Cats, Mediated by the Olfactory and Vomeronasal Senses: I. Chemocommunication. Z. Tierpsychol. 1976, 42, 86–109. [Google Scholar]
- Mellen, J.D. A Comparative Analysis of Scent-Marking, Social and Reproductive Behavior in 20 Species of Small Cats (Felis). Am. Zool. 1993, 33, 151–166. [Google Scholar] [CrossRef] [Green Version]
- Martínez-Macipe, M.; Lafont-Lecuelle, C.; Manteca, X.; Pageat, P.; Cozzi, A. Evaluation of an innovative approach for sensory enrichment in zoos: Semiochemical stimulation for captive lions (Panthera leo). Anim. Welf. 2015, 24, 455–461. [Google Scholar] [CrossRef]
- Soso, S.B. Chemical Constituency and Odor of Semiochemicals: Studying the Chemical Composition and Odor of Volatile Organic Compounds of Great Cat Marking Fluid in an Effort to Aid Tiger and Lion Conservation. Ph.D. Thesis, Iowa State University, Ames, IA, USA, 2018. [Google Scholar] [CrossRef]
- Burger, B.V.; Visser, R.; Moses, A.; Le Roux, M. Elemental Sulfur Identified in Urine of Cheetah, Acinonyx jubatus. J. Chem. Ecol. 2006, 32, 1347–1352. [Google Scholar] [CrossRef] [Green Version]
- Tommasi, A.; Tredoux, A.G.J.; Koziel, J.A.; Esposito, G. Chemical Characterization of the Marking Fluid of Breeding and Non-Breeding Male Cheetahs. Animals 2022, 12, 2284. [Google Scholar] [CrossRef] [PubMed]
- Mossotti, R.H. Female Reaction to Male Urine Scents as Potential Indicator of Mate Choice in Captive Cheetahs (Acinonyx jubatus). Ph.D. Thesis, Southern Illinois University, Carbondale, IL, USA, 2010. [Google Scholar]
- Cuadros-Rodríguez, L.; Gámiz-Gracia, L.; Almansa-López, E.M.; Bosque-Sendra, J.M. Calibration in chemical measurement processes. II. A methodological approach. TrAC Trends Anal. Chem. 2001, 20, 620–636. [Google Scholar] [CrossRef]
- Yu, Y.-S.; Sun, L.; Huang, X.; Zhou, J.-Z. Evaporation of ethanol/water mixture droplets on a pillar-like PDMS surface. Colloids Surf. A Physicochem. Eng. Asp. 2019, 574, 215–220. [Google Scholar] [CrossRef] [Green Version]
- Chaudhuri, S.R.; Gajjar, R.M.; Krantz, W.B.; Kasting, G.B. Percutaneous absorption of volatile solvents following transient liquid exposures II. Ethanol. Chem. Eng. Sci. 2009, 64, 1665–1672. [Google Scholar] [CrossRef]
- Vuilleumier, C.; Flament, I.; Sauvegrain, P. Headspace analysis study of evaporation rate of perfume ingredients applied onto skin. Int. J. Cosmet. Sci. 1995, 17, 61–76. [Google Scholar] [CrossRef] [PubMed]
- Little, C.J.L.; Ferasin, L.; Ferasin, H.; Holmes, M.A. Purring in Cats during Auscultation: How Common Is It, and Can We Stop It? J. Small Anim. Pract. 2014, 55, 33–38. [Google Scholar] [CrossRef]
- Graham, L. Investigating the Use of the Pheromone Feliway to Reduce Relocation Stress in Tigers (Panthera tigris). In Proceedings of the CCSAW Research Day, Guelph, ON, Canada, 11 May 2016. [Google Scholar]
- Pereira, J.S.; Fragoso, S.; Beck, A.; Lavigne, S.; Varejão, A.S.; da Graça Pereira, G. Improving the feline veterinary consultation: The usefulness of Feliway spray in reducing cats’ stress. J. Feline Med. Surg. 2016, 18, 959–964. [Google Scholar] [CrossRef]
- Tommasi, A.; Koziel, J.A.; Molotsi, A.H.; Esposito, G. Understanding the Role of Semiochemicals on the Reproductive Behaviour of Cheetahs (Acinonyx jubatus)—A Review. Animals 2021, 11, 3140. [Google Scholar] [CrossRef] [PubMed]
- Wright, D.W.; Koziel, J.A.; Parker, D.B.; Iwasinska, A.; Hartman, T.G.; Kolvig, P.; Wahe, L. Qualitative Exploration of the ‘Rolling Unmasking Effect’ for Downwind Odor Dispersion from a Model Animal Source. Int. J. Environ. Res. Public Health 2021, 18, 13085. [Google Scholar] [CrossRef]
- Good Scents Company. Available online: http://www.thegoodscentscompany.com/search.html (accessed on 18 November 2021).
- Banik, C.; Koziel, J.A.; Li, J.Z. Simultaneous Chemical and Sensory Analysis of Domestic Cat Urine and Feces with Headspace Solid-Phase Microextraction and GC-MS-Olfactometry. Separations 2021, 8, 15. [Google Scholar] [CrossRef]
- Lee, M.; Li, P.; Koziel, J.A.; Ahn, H.; Wi, J.; Chen, B.; Meiirkhanuly, Z.; Banik, C.; Jenks, W.S. Pilot-Scale Testing of UV-A Light Treatment for Mitigation of NH3, H2S, GHGs, VOCs, Odor, and O3 Inside the Poultry Barn. Front. Chem. 2020, 8, 613. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.; Koziel, J.A.; Murphy, W.; Jenks, W.S.; Chen, B.; Li, P.; Banik, C. Evaluation of TiO2 Based Photocatalytic Treatment of Odor and Gaseous Emissions from Swine Manure with UV-A and UV-C. Animals 2021, 11, 1289. [Google Scholar] [CrossRef] [PubMed]
- Rowe, C. Receiver psychology and the evolution of multicomponent signals. Anim. Behav. 1999, 58, 921–931. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Volatile Organic Compound (VOC) | Total in 2 L of Ethanol (mg) |
|---|---|
| Benzaldehyde | 68 |
| Dimethyl disulphide | 5.48 |
| Phenol | 3.95 |
| Acetophenone | 28.18 |
| Indole | 3.72 |
| Behaviour | Definition |
|---|---|
| Object rub | Rubs face, head, neck, or flanks on object (e.g., on fence, tree). |
| Roll | Rolls on back, rubbing the back on the ground while all paws are in the air, or rolls from one side to the other (each roll is then recorded as one occurrence). |
| Object sniff | Olfactory examination of ground (e.g., urine or faeces) or structures. |
| Urine spray | Urinating in standing position with tail raised against a vertical structure (frequently trees or huts). Visually the same as male urine spraying; however, females are not able to spray directionally like males. |
| Meow-chirp | Meow: a soft call, low-pitched, similar to domestic cat. Occasionally chirps would be emitted together with meows. Chirps were more high-pitched than meows and very short. However, these chirps were not as high-pitched, short, and loud as the chirps emitted when threatened. |
| Behaviour | Age Group | Treatment (SS) | Observation Method | ||||||
|---|---|---|---|---|---|---|---|---|---|
| 2–4 Years | 6–8 Years | p-Value | Pre | Post | p-Value | Camera | In Person | p-Value | |
| Roll | 1.73 ± 0.67 | 1.24 ± 1.29 | 0.36 | 1.98 ± 1.07 | 1.06 ± 0.66 | 0.11 | 1.52 ± 1.81 | 1.40 ± 1.72 | 0.42 |
| Rub | 1.02 ± 0.69 | 2.70 ± 1.41 | 0.04 | 1.50 ± 0.89 | 1.98 ± 1.71 | 0.38 | 1.57 ± 1.71 | 1.21 ± 1.93 | 0.06 |
| Sniff | 5.29 ± 1.41 | 6.92 ± 1.63 | 0.10 | 5.11 ± 1.73 | 6.87 ± 1.12 | 0.02 | 6.63 ± 3.00 | 4.81 ± 3.29 | 0.07 |
| Spray | 0.22 ± 0.41 | 0.28 ± 0.53 | 0.87 | 0.33 ± 0.58 | 0.15 ± 0.29 | 0.23 | 0.00 ± 0.00 | 0.24 ± 0.52 | 0.37 |
| Total (No MC) | 8.26 ± 1.56 | 11.14 ± 2.91 | 0.07 | 8.92 ± 3.18 | 10.07 ± 1.92 | 0.39 | 9.72 ± 3.89 | 7.67 ± 4.66 | 0.04 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tommasi, A.; Tredoux, A.G.J.; Koziel, J.A.; Esposito, G. The Effect of a Synthetic Scent on Cheetah Behaviour. Animals 2023, 13, 369. https://doi.org/10.3390/ani13030369
Tommasi A, Tredoux AGJ, Koziel JA, Esposito G. The Effect of a Synthetic Scent on Cheetah Behaviour. Animals. 2023; 13(3):369. https://doi.org/10.3390/ani13030369
Chicago/Turabian StyleTommasi, Alexia, Andreas G. J. Tredoux, Jacek A. Koziel, and Giulia Esposito. 2023. "The Effect of a Synthetic Scent on Cheetah Behaviour" Animals 13, no. 3: 369. https://doi.org/10.3390/ani13030369
APA StyleTommasi, A., Tredoux, A. G. J., Koziel, J. A., & Esposito, G. (2023). The Effect of a Synthetic Scent on Cheetah Behaviour. Animals, 13(3), 369. https://doi.org/10.3390/ani13030369