

Characterization of Myeloperoxidase in the Healthy Equine Endometrium
 , ,
, ,  ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals and Samples Collection
2.2. Histology
2.3. Total Myeloperoxidase by ELISA Assay
2.4. Active Myeloperoxidase by SIEFED Assay
2.5. Protein Concentration
2.6. Immunochemistry
2.7. Semiquantitative Evaluation of the Myeloperoxidase Immunoexpression
2.8. Statistical Analysis
3. Results
3.1. Histology Evaluation
3.2. Myeloperoxidase
3.3. Protein Concentration
3.4. Total Myeloperoxidase/Total Protein Ratio (RT)
3.5. Active Myeloperoxidase/Total Protein Ratio (RA)
3.6. Specific Activity of Myeloperoxidase
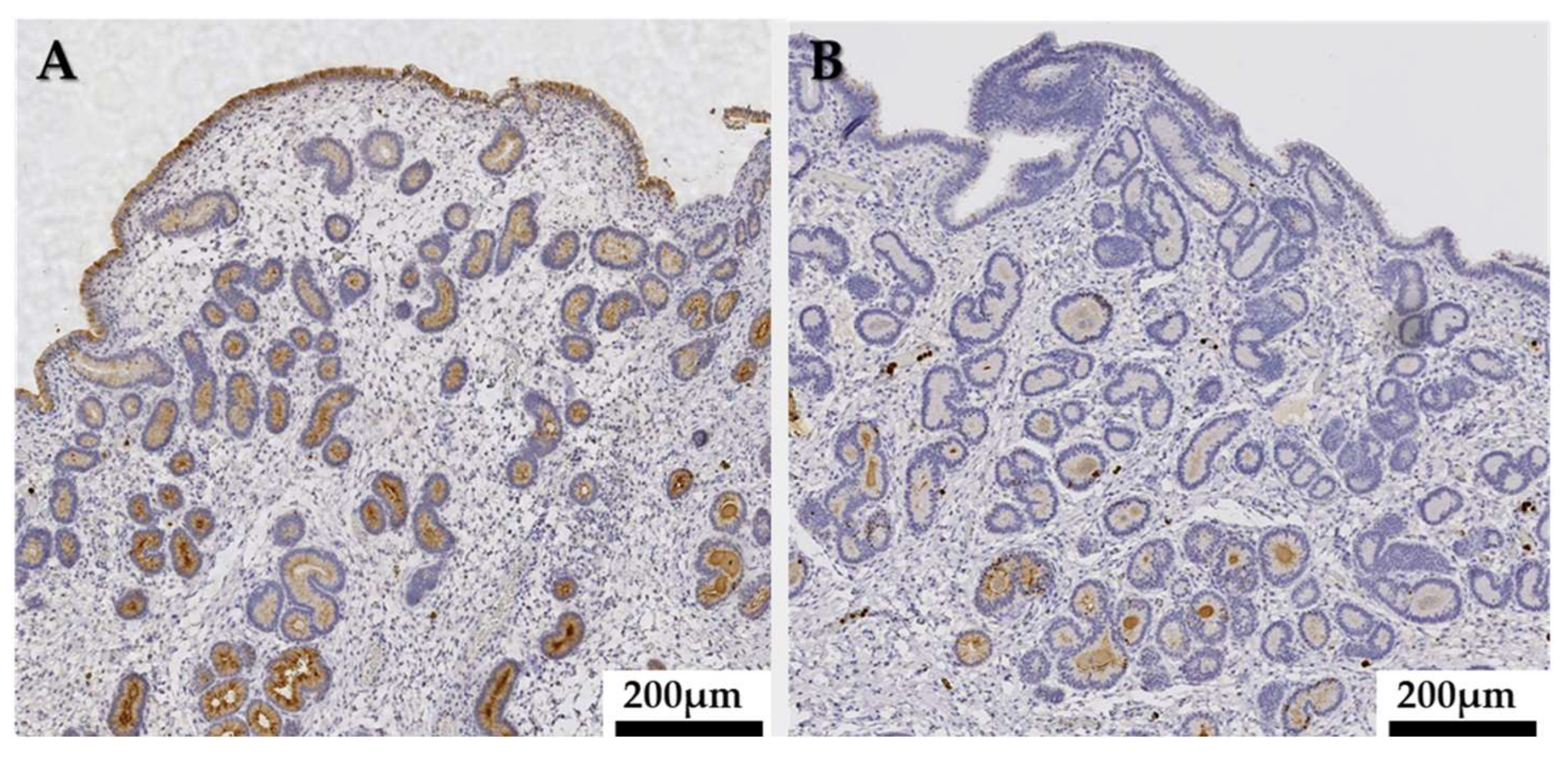
3.7. Immunohistochemistry
3.7.1. MPO Expression in the Endometrium
3.7.2. Semiquantitative MPO Immunoexpression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mayadas, T.N.; Cullere, X.; Lowell, C.A. The Multifaceted Functions of Neutrophils. Annu. Rev. Pathol. Mech. Dis. 2014, 9, 181–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klebanoff, S.J. Myeloperoxidase: Friend and foe. J. Leukoc. Biol. 2005, 77, 598–625. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Borregaard, N.; Cowland, J. Granules of the Human Neutrophilic Polymorphonuclear Leukocyte. Blood 1997, 89, 3503–3521. [Google Scholar] [CrossRef] [PubMed]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil Extra-cellular Traps Kill Bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.A.; Alsahli, M.A.; Rahmani, A.H. Myeloperoxidase as an Active Disease Biomarker: Recent Biochemical and Pathological Perspectives. Med. Sci. 2018, 6, 33. [Google Scholar] [CrossRef] [Green Version]
- Friedrichs, K.; Baldus, S.; Klinke, A. Fibrosis in Atrial Fibrillation—Role of Reactive Species and MPO. Front. Physiol. 2012, 3, 214. [Google Scholar] [CrossRef] [Green Version]
- Rudolph, V.; Andrié, R.P.; Rudolph, T.K.; Friedrichs, K.; Klinke, A.; Hirsch-Hoffmann, B.; Schwoerer, A.P.; Lau, D.; Fu, X.; Klingel, K.; et al. Myeloperoxidase acts as a profibrotic mediator of atrial fibrillation. Nat. Med. 2010, 16, 470–474. [Google Scholar] [CrossRef] [Green Version]
- Pulli, B.; Ali, M.; Iwamoto, Y.; Zeller, M.W.; Schob, S.; Linnoila, J.J.; Chen, J.W. Myeloperoxidase–Hepatocyte–Stellate Cell Cross Talk Promotes Hepatocyte Injury and Fibrosis in Experimental Nonalcoholic Steatohepatitis. Antioxid. Redox Signal. 2015, 23, 1255–1269. [Google Scholar] [CrossRef] [Green Version]
- Austin, G.E.; Zhao, W.-G.; Adjiri, A.; Lu, J.-P. Control of myeloperoxidase gene expression in developing myeloid cells. Leuk. Res. 1996, 20, 817–820. [Google Scholar] [CrossRef]
- Green, P.S.; Mendez, A.J.; Jacob, J.S.; Crowley, J.R.; Growdon, W.; Hyman, B.T.; Heinecke, J.W. Neuronal expression of myeloperoxidase is increased in Alzheimer’s disease. J. Neurochem. 2004, 90, 724–733. [Google Scholar] [CrossRef]
- Martínez-Marcos, P.; Carvajal-Serna, M.; Lázaro-Gaspar, S.; Pérez-Pé, R.; Muiño-Blanco, T.; Cebrián-Pérez, J.A.; Casao, A. Presence of melatonin-catabolizing non-specific enzymes myeloperoxidase and indoleamine 2,3-dioxygenase in the ram reproductive tract. Reprod Dom. Anim. 2019, 54, 1643–1650. [Google Scholar] [CrossRef]
- Roumeguère, T.; Delree, P.; Van Antwerpen, P.; Rorive, S.; Vanhamme, L.; Ryhove, L.D.L.K.D.; Serteyn, D.; Wespes, E.; Vanhaerverbeek, M.; Boudjeltia, K.Z. Intriguing location of myeloperoxidase in the prostate: A preliminary immunohistochemical study. Prostate 2011, 72, 507–513. [Google Scholar] [CrossRef]
- Yang, Z.; Xiao, K.; Wang, W.; Tang, J.; Sun, P.-P.; Peng, K.-M.; Song, H. The Effect of Visfatin on Inflammatory Reaction in Uterus of LPS-Induced Rats. Int. J. Morphol. 2015, 33, 194–203. [Google Scholar] [CrossRef] [Green Version]
- Haegens, A.; Vernooy, J.H.J.; Heeringa, P.; Mossman, B.T.; Wouters, E.F.M. Myeloperoxidase modulates lung epithelial responses to pro-inflammatory agents. Eur. Respir. J. 2008, 31, 252–260. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.J.; Preston, G.A.; Pendergraft, W.F.; Segelmark, M.; Heeringa, P.; Hogan, S.L.; Jennette, J.C.; Falk, R.J. Internalization of Proteinase 3 Is Concomitant with Endothelial Cell Apoptosis and Internalization of Myeloperoxidase with Generation of Intracellular Oxidants. Am. J. Pathol. 2001, 158, 581–592. [Google Scholar] [CrossRef] [Green Version]
- Jerke, U.; Rolle, S.; Purfürst, B.; Luft, F.; Nauseef, W.; Kettritz, R. β2 Integrin-mediated Cell-Cell Contact Transfers Active Myeloperoxidase from Neutrophils to Endothelial Cells*. J. Biol. Chem. 2013, 288, 12910–12919. [Google Scholar] [CrossRef] [Green Version]
- La Rocca, G.; Di Stefano, A.; Eleuteri, E.; Anzalone, R.; Magno, F.; Corrao, S.; Loria, T.; Martorana, A.; Di Gangi, C.; Colombo, M.; et al. Oxidative stress induces myeloperoxidase expression in endocardial endothelial cells from patients with chronic heart failure. Basic Res. Cardiol. 2009, 104, 307–320. [Google Scholar] [CrossRef]
- Art, T.; Franck, T.; Lekeux, P.; de Moffarts, B.; Couëtil, L.; Becker, M.; Kohnen, S.; Deby-Dupont, G.; Serteyn, D. Myeloperoxidase con-centration in bronchoalveolar lavage fluid from healthy horses and those with recurrent airway obstruction. Can. J. Vet. Res. 2006, 70, 291–296. [Google Scholar]
- Wauters, J.; Pille, F.; Martens, A.; Franck, T.; Serteyn, D.; Gasthuys, F.; Meyer, E. Equine myeloperoxidase: A novel biomarker in synovial fluid for the diagnosis of infection. Equine Vet. J. 2012, 45, 278–283. [Google Scholar] [CrossRef]
- Ponthier, J.; Franck, T.; Parrilla-Hernandez, S.; Niesten, A.; De La Rebiere, G.; Serteyn, D.; Deleuze, S. Concentration, Activity and Biochemical Characterization of Myeloperoxidase in Fresh and Post-Thaw Equine Semen and their Implication on Freezability. Reprod. Domest. Anim. 2014, 49, 285–291. [Google Scholar] [CrossRef]
- Art, T.; Franck, T.; Gangl, M.; Votion, D.; Kohnen, S.; Deby-Dupont, G.; Serteyn, D. Plasma concentrations of myeloperoxidase in endurance and 3-day event horses after a competition. Equine Vet. J. 2006, 38, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Grulke, S.; Franck, T.; Gangl, M.; Péters, F.; Salciccia, A.; Deby-Dupont, G.; Serteyn, D. Myeloperoxidase assay in plasma and peri-toneal fluid of horses with gastrointestinal disease. Can. J. Vet. Res. 2008, 72, 37–42. [Google Scholar] [PubMed]
- Storms, N.; Torres, C.M.; Franck, T.; Guitart, A.S.; de la Rebière, G.; Serteyn, D. Presence of Myeloperoxidase in Lamellar Tissue of Horses Induced by an Euglycemic Hyperinsulinemic Clamp. Front. Vet. Sci. 2022, 9, 6835. [Google Scholar] [CrossRef] [PubMed]
- Parrilla-Hernandez, S.; Ponthier, J.; Franck, T.Y.; Serteyn, D.D.; Deleuze, S.C. High concentrations of myeloperoxidase in the equine uterus as an indicator of endometritis. Theriogenology 2014, 81, 936–940. [Google Scholar] [CrossRef] [Green Version]
- Rebordão, M.R.; Amaral, A.; Fernandes, C.; Silva, E.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Enzymes Present in Neutrophil Extracellular Traps May Stimulate the Fibrogenic PGF2α Pathway in the Mare Endometrium. Animals 2021, 11, 2615. [Google Scholar] [CrossRef]
- Rebordão, M.R.; Amaral, A.; Lukasik, K.; Szóstek-Mioduchowska, A.; Pinto-Bravo, P.; Galvão, A.; Skarzynski, D.J.; Ferreira-Dias, G. Constituents of neutrophil extracellular traps induce in vitro collagen formation in mare endometrium. Theriogenology 2018, 113, 8–18. [Google Scholar] [CrossRef]
- Rebordão, M.; Carneiro, C.; Alexandre-Pires, G.; Brito, P.; Pereira, C.; Nunes, T.; Galvão, A.; Leitão, A.; Vilela, C.; Ferreira-Dias, G. Neutrophil extracellular traps formation by bacteria causing endometritis in the mare. J. Reprod. Immunol. 2014, 106, 41–49. [Google Scholar] [CrossRef]
- Nazhat, S.A.; Kitahara, G.; Kozuka, N.; Mido, S.; Sadawy, M.; Ali, H.E.-S.; Osawa, T. Associations of periparturient plasma biochemical parameters, endometrial leukocyte esterase and myeloperoxidase, and bacterial detection with clinical and subclinical en-dometritis in postpartum dairy cows. J. Vet. Med. Sci. 2018, 80, 302–310. [Google Scholar] [CrossRef] [Green Version]
- Amaral, A.; Fernandes, C.; Rebordão, M.; Szóstek-Mioduchowska, A.; Lukasik, K.; Pinto-Bravo, P.; da Gama, L.T.; Skarzynski, D.J.; Ferreira-Dias, G. Myeloperoxidase Inhibition Decreases the Expression of Collagen and Metallopeptidase in Mare Endometria under In Vitro Conditions. Animals 2021, 11, 208. [Google Scholar] [CrossRef]
- Chapman, A.L.P.; Mocatta, T.J.; Shiva, S.; Seidel, A.; Chen, B.; Khalilova, I.; Paumann-Page, M.E.; Jameson, G.N.L.; Winterbourn, C.C.; Kettle, A.J. Ceruloplasmin Is an Endogenous Inhibitor of Myeloperoxidase. J. Biol. Chem. 2013, 288, 6465–6477. [Google Scholar] [CrossRef] [Green Version]
- Segelmark, M.; Persson, B.; Hellmark, T.; Wieslander, J. Binding and inhibition of myeloperoxidase (MPO): A major function of ceruloplasmin? Clin. Exp. Immunol. 1997, 108, 167–174. [Google Scholar] [CrossRef]
- Panzani, D.; Di Vita, M.; Lainé, A.-L.; Guillaume, D.; Rota, A.; Tesi, M.; Vannozzi, I.; Camillo, F. Corpus Luteum Vascularization and Progesterone Production in Autumn and Winter Cycles of the Mare: Relationship Between Ultrasonographic Characteristics of Corpora Lutea and Plasma Progesterone Concentration in the Last Cycles Before Anestrus. J. Equine Vet. Sci. 2017, 56, 35–39. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, M.M.; Magsig, J.; Stromberg, A.J. Use of a low-volume uterine flush for diagnosing endometritis in chronically infertile mares. Theriogenology 2007, 68, 403–412. [Google Scholar] [CrossRef]
- Kozdrowski, R.; Sikora, M.; Buczkowska, J.; Nowak, M.; Raś, A.; Dzięcioł, M. Effects of cycle stage and sampling procedure on interpretation of endometrial cytology in mares. Anim. Reprod. Sci. 2015, 154, 56–62. [Google Scholar] [CrossRef]
- Overbeck, W.; Witte, T.; Heuwieser, W. Comparison of three diagnostic methods to identify subclinical endometritis in mares. Theriogenology 2011, 75, 1311–1318. [Google Scholar] [CrossRef]
- Kenney, R.M.D.P. Equine Endometrial Biopsy. In Current Therapy in Theriogenology; Da, M., Ed.; W.B Saunders Company: Philadelphia, PA, USA, 1986; pp. 723–729. [Google Scholar]
- Schoon, H.-A.; Schoon, D.; Klug, E. Endometrial biopsies as an ancillary aid in diagnosis and prognosis of subfertility in the mare. Pferdeheilkunde Equine Med. 1992, 8, 355–362. [Google Scholar] [CrossRef]
- Franck, T.; Grulke, S.; Deby-Dupont, G.; Deby, C.; Duvivier, H.; Péters, F.; Serteyn, D. Development of an enzyme-linked im-munosorbent assay for specific equine neutrophil myeloperoxidase measurement in blood. J. Vet. Diagn. Invest. 2005, 17, 412–419. [Google Scholar] [CrossRef] [Green Version]
- Franck, T.; Kohnen, S.; Deby-Dupont, G.; Grulke, S.; Deby, C.; Serteyn, D. A Specific Method for Measurement of Equine Active Myeloperoxidase in Biological Samples and in in Vitro Tests. J. Vet. Diagn. Investig. 2006, 18, 326–334. [Google Scholar] [CrossRef] [Green Version]
- Hoffmann, C.; Bazer, F.W.; Klug, J.; Aupperle, H.; Ellenberger, C.; Schoon, H.A. Immunohistochemical and histochemical identifi-cation of proteins and carbohydrates in the equine endometrium. Theriogenology 2009, 71, 264–274. [Google Scholar] [CrossRef]
- Khalilova, I.S.; Dickerhof, N.; Mocatta, T.J.; Bhagra, C.J.; McClean, D.R.; Obinger, C.; Kettle, A.J. A myeloperoxidase precursor, pro-myeloperoxidase, is present in human plasma and elevated in cardiovascular disease patients. PLoS ONE 2018, 13, e0192952. [Google Scholar] [CrossRef] [Green Version]
- Kenney, R.; Doig, P. Equine endometrial biopsy. Curr. Ther. Ther. 1986, 2, 723–729. [Google Scholar]
- Hoffmann, C.; Bazer, F.W.; Klug, J.; Aupperle, H.; Ellenberger, C.; Özgen, S.; Schoon, D.; Schoon, H.-A. Immunohistochemical and histochemical identification of proteins and carbohydrates in the equine endometrium: Reaction patterns in the cycling mare. Pferdeheilkunde Equine Med. 2009, 25, 212–219. [Google Scholar] [CrossRef]
- Schöniger, S.; Gräfe, H.; Richter, F.; Schoon, H.-A. Expression of indoleamine 2,3-dioxygenase 1 as transcript and protein in the healthy and diseased equine endometrium. Res. Vet. Sci. 2018, 118, 278–287. [Google Scholar] [CrossRef] [PubMed]
- Schöniger, S.; Gräfe, H.; Schoon, H.-A. Beta-defensin is a component of the endometrial immune defence in the mare. Pferdeheilkunde Equine Med. 2013, 29, 335–346. [Google Scholar] [CrossRef] [Green Version]
- Schöniger, S.; Böttcher, D.; Theuß, T.; Schoon, H.-A. Expression of Toll-like receptors 2, 4 and 6 in equine endometrial epithelial cells: A comparative in situ and in vitro study. Res. Vet. Sci. 2017, 112, 34–41. [Google Scholar] [CrossRef]
- Churg, A.; Anderson, W.A. Induction of Endometrial Peroxidase Synthesis and Secretion by Estrogen and Estrogen Antagonist. J. Cell Biol. 1974, 62, 449–459. [Google Scholar] [CrossRef]
- McRae, A.C. The blood-uterine lumen barrier and exchange between extracellular fluids. Reproduction 1988, 82, 857–873. [Google Scholar] [CrossRef] [Green Version]
- Ceusters, J.D.; Mouithys-Mickalad, A.A.; Franck, T.J.; Derochette, S.; Vanderplasschen, A.; Deby-Dupont, G.P.; Serteyn, D.A. Effect of myeloperoxidase and anoxia/reoxygenation on mitochondrial respiratory function of cultured primary equine skeletal myoblasts. Mitochondrion 2013, 13, 410–416. [Google Scholar] [CrossRef]
- de Araujo, T.H.; Okada, S.S.; Ghosn, E.E.; Taniwaki, N.N.; Rodrigues, M.R.; de Almeida, S.R.; Mortara, R.A.; Russo, M.; Campa, A.; Albu-querque, R.C. Intracellular localization of myeloperoxidase in murine peritoneal B-lymphocytes and macrophages. Cell Immunol. 2013, 281, 27–30. [Google Scholar] [CrossRef]
- Wira, C.R.; Grant-Tschudy, K.S.; Crane-Godreau, M.A. Epithelial Cells in the Female Reproductive Tract: A Central Role as Sentinels of Immune Protection. Am. J. Reprod. Immunol. 2005, 53, 65–76. [Google Scholar] [CrossRef]
- Kolm, G.; Klein, D.; Knapp, E.; Watanabe, K.; Walter, I. Lactoferrin expression in the horse endometrium: Relevance in persisting mating-induced endometritis. Veter- Immunol. Immunopathol. 2006, 114, 159–167. [Google Scholar] [CrossRef]
- Katila, T.; Lock, T.; Hoffmann, W.; Smith, A. Lysozyme, alkaline phosphatase and neutrophils in uterine secretions of mares with differing resistance to endometritis. Theriogenology 1990, 33, 723–732. [Google Scholar] [CrossRef] [PubMed]
- Shibata, T.; Sakamoto, J.; Osaka, Y.; Neyatani, N.; Fujita, S.; Oka, Y.; Takagi, H.; Mori, H.; Fujita, H.; Tanaka, Y.; et al. Myelop-eroxidase in blood neutrophils during normal and abnormal menstrual cycles in women of reproductive age. Int. J. Lab. Hematol. 2017, 39, 169–174. [Google Scholar] [CrossRef]
- Békési, G.; Kakucs, R.; Varbiro, S.; Feher, J.; Pazmany, T.; Magyar, Z.; Sprintz, D.; Szekacs, B. Induced myeloperoxidase activity and related superoxide inhibition during hormone replacement therapy. BJOG Int. J. Obstet. Gynaecol. 2001, 108, 474–481. [Google Scholar] [CrossRef]
- Houdeau, E.; Larauche, M.; Monnerie, R.; Bueno, L.; Fioramonti, J. Uterine motor alterations and estrous cycle disturbances as-sociated with colonic inflammation in the rat. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2005, 288, 630–637. [Google Scholar] [CrossRef] [Green Version]
- Baiza-Gutman, L.; Flores-Sánchez, M.; Diaz-Flores, M.; Hicks, J. Presence of uterine peroxidase activity in the rat early pregnancy. Int. J. Biochem. Cell Biol. 2000, 32, 255–262. [Google Scholar] [CrossRef]
- Cummings, A.M.; Metcalf, J.L. Mechanisms of the stimulation of rat uterine peroxidase activity by methoxychlor. Reprod. Toxicol. 1994, 8, 477–486. [Google Scholar] [CrossRef]
- Farley, D.B.; Ford, S.P.; Rosazza, J.P.N. Increase in uterine peroxidase activity in the rat uterus during oestrogen hyperaemia. Reproduction 1992, 95, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Okada, H.; Nawa, M.; Kimura, J. In Vitro Effect of Estrogens on the Peroxidase Activity of Human Endometrium*. J. Clin. Endocrinol. Metab. 1985, 61, 404–407. [Google Scholar] [CrossRef]
- Batra, S.C.; Iosif, C.S. Effect of Estrogen Treatment on the Peroxidase Activity and Estrogen Receptors in the Female Rabbit Urogenital Tissues. J. Urol. 1992, 148, 935–938. [Google Scholar] [CrossRef]
- Grant-Tschudy, K.S.; Wira, C.R. Effect of Estradiol on Mouse Uterine Epithelial Cell Transepithelial Resistance (TER). Am. J. Reprod. Immunol. 2004, 52, 252–262. [Google Scholar] [CrossRef] [PubMed]
- Lewis, G.S. Role of ovarian progesterone and potential role of prostaglandin F2α and prostaglandin E2 in modulating the uterine response to infectious bacteria in postpartum ewes. J. Anim. Sci. 2003, 81, 285–293. [Google Scholar] [CrossRef] [PubMed]
- Watson, E.D.; Stokes, C.R.; Bourne, F.J. Influence of administration of ovarian steroids on the function of neutrophils isolated from the blood and uterus of ovariectomized mares. J. Endocrinol. 1987, 112, 443–448. [Google Scholar] [CrossRef]
- Chadwick, J.W.; Fine, N.; Khoury, W.; Tasevski, N.; Sun, C.; Boroumand, P.; Klip, A.; Glogauer, M. Tissue-specific murine neutrophil activation states in health and inflammation. J. Leukoc. Biol. 2020, 110, 187–195. [Google Scholar] [CrossRef] [PubMed]
- Seals, R.C.; Wulster-Radcliffe, M.C.; Lewis, G.S. Modulation of the Uterine Response to Infectious Bacteria in Postpartum Ewes. Am. J. Reprod. Immunol. 2002, 47, 57–63. [Google Scholar] [CrossRef]
- Klebanoff, S.J. Iodination of Bacteria: A Bactericidal Mechanism. J. Exp. Med. 1967, 126, 1063–1078. [Google Scholar] [CrossRef] [Green Version]
- Davies, M.J.; Hawkins, C.L. The Role of Myeloperoxidase in Biomolecule Modification, Chronic Inflammation, and Disease. Antioxid. Redox Signal. 2020, 32, 957–981. [Google Scholar] [CrossRef] [Green Version]
- Kargapolova, Y.; Geißen, S.; Zheng, R.; Baldus, S.; Winkels, H.; Adam, M. The Enzymatic and Non-Enzymatic Function of Myeloperoxidase (MPO) in Inflammatory Communication. Antioxidants 2021, 10, 562. [Google Scholar] [CrossRef]
- Reilas, T.; del Alamo, M.M.R.; Liepina, E.; Yeste, M.; Katila, T. Effects on the equine endometrium of cervical occlusion after insemination. Theriogenology 2016, 85, 617–624. [Google Scholar] [CrossRef]
- Ricketts, S.W.; Alonso, S. The effect of age and parity on the development of equine chronic endometrial disease. Equine Vet. J. 1991, 23, 189–192. [Google Scholar] [CrossRef]
- Bradley, P.P.; Christensen, R.D.; Rothstein, G. Cellular and Extracellular Myeloperoxidase in Pyogenic Inflammation. Blood 1982, 60, 618–622. [Google Scholar] [CrossRef] [Green Version]
- Edwards, S.W.; Nurcombe, H.L.; A Hart, C. Oxidative inactivation of myeloperoxidase released from human neutrophils. Biochem. J. 1987, 245, 925–928. [Google Scholar] [CrossRef]
- King, C.C.; Jefferson, M.M.; Thomas, E.L. Secretion and inactivation of myeloperoxidase by isolated neutrophils. J. Leukoc. Biol. 1997, 61, 293–302. [Google Scholar] [CrossRef]
- Klebanoff, S.J. Effect of estrogens on the myeloperoxidase-mediated antimicrobial system. Infect. Immun. 1979, 25, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Galijasevic, S.; Abdulhamid, I.; Abu-Soud, H.M. Melatonin Is a Potent Inhibitor for Myeloperoxidase. Biochemistry 2008, 47, 2668–2677. [Google Scholar] [CrossRef]
- Galijasevic, S.; Abdulhamid, I.; Abu-Soud, H.M. Potential role of tryptophan and chloride in the inhibition of human myelop-eroxidase. Free Radic. Biol. 2008, 44, 1570–1577. [Google Scholar] [CrossRef] [Green Version]
- Eklinke, A.; Nussbaum, C.; Kubala, L.; Friedrichs, K.; Rudolph, T.K.; Rudolph, V.; Paust, H.-J.; Schröder, C.; Benten, D.; Lau, D.; et al. Myeloperoxidase attracts neutrophils by physical forces. Blood 2011, 117, 1350–1358. [Google Scholar] [CrossRef] [Green Version]
- Lefkowitz, D.L.; Gelderman, M.P.; Fuhrmann, S.R.; Graham, S.; Starnes, J.D.; Lefkowitz, S.S.; Bollen, A.; Moguilevsky, N. Neutrophilic Myeloperoxidase–Macrophage Interactions Perpetuate Chronic Inflammation Associated with Experimental Arthritis. Clin. Immunol. 1999, 91, 145–155. [Google Scholar] [CrossRef]
- Lefkowitz, D.L.; Roberts, E.; Grattendick, K.; Schwab, C.; Stuart, R.; Lincoln, J.; Allen, R.C.; Moguilevsky, N.; Bollen, A.; Lefkowitz, S.S. The Endothelium and Cytokine Secretion: The Role of Peroxidases as Immunoregulators. Cell Immunol. 2000, 202, 23–30. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total MPO (ng/mL) | Active MPO (ng/mL) | Protein (mg/mL) | RT (ng/mg) | RA (ng/mg) | |
|---|---|---|---|---|---|
| Anestrus | 3798 (+/− 3464) a | 4.42 (+/− 0.84) a | 0.24 (+/− 0.26) a | 18512 (+/− 3994) a | 22.08 (+/− 3.46) a |
| Diestrus | 2319 (+/− 2334) a | 0.52 (+/− 0.54) b | 0.41 (+/− 0.17) a | 4320 (+/− 2517) b | 1.75 (+/− 2.04) b |
| Estrus | 8867 (+/− 1947) a | 0.85 (+/− 0.36) b | 0.77 (+/− 0.14) a | 13930 (+/− 2132) a | 1.48 (+/− 1.06) b |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Parrilla Hernández, S.; Franck, T.; Munaut, C.; Feyereisen, É.; Piret, J.; Farnir, F.; Reigner, F.; Barrière, P.; Deleuze, S. Characterization of Myeloperoxidase in the Healthy Equine Endometrium. Animals 2023, 13, 375. https://doi.org/10.3390/ani13030375
Parrilla Hernández S, Franck T, Munaut C, Feyereisen É, Piret J, Farnir F, Reigner F, Barrière P, Deleuze S. Characterization of Myeloperoxidase in the Healthy Equine Endometrium. Animals. 2023; 13(3):375. https://doi.org/10.3390/ani13030375
Chicago/Turabian StyleParrilla Hernández, Sonia, Thierry Franck, Carine Munaut, Émilie Feyereisen, Joëlle Piret, Frédéric Farnir, Fabrice Reigner, Philippe Barrière, and Stéfan Deleuze. 2023. "Characterization of Myeloperoxidase in the Healthy Equine Endometrium" Animals 13, no. 3: 375. https://doi.org/10.3390/ani13030375
APA StyleParrilla Hernández, S., Franck, T., Munaut, C., Feyereisen, É., Piret, J., Farnir, F., Reigner, F., Barrière, P., & Deleuze, S. (2023). Characterization of Myeloperoxidase in the Healthy Equine Endometrium. Animals, 13(3), 375. https://doi.org/10.3390/ani13030375