1. Introduction
Research into the use of microalgae as an alternative ingredient to fishmeal and terrestrial-plant ingredients has recently gained attention due to several features that make them attractive to the aquaculture industry [
1]. Owing to their high nutritional value,
Chlorella species have been used in the dietary formulations of several fish species, evidencing beneficial effects on growth, nutrient utilization, fatty acid content, and fish flesh quality, as well as improving their stress resistance and survival rates [
2,
3,
4,
5,
6]. However, the nutritional effects of
Chlorella species depend on the levels at which they are included in the aquafeeds. In this regard, Ahmad et al. [
7] reviewed contradictory results on growth and feed efficiency in several fish species, the contradictions possibly being related to the inclusion level of the microalgae, since levels above 20% affected growth performance [
8,
9].
Probiotics are live microorganisms that confer a health benefit on the host when administered in adequate amounts [
10,
11]. Numerous studies have documented their application in human and farm animal health [
12,
13,
14]. The beneficial features of probiotics include (i) improved nutrient supply to the host, (ii) an increase in aquafeed digestibility, (iii) the ability to colonize mucosal surfaces, acting as a barrier against pathogens, (iv) modulation of the gut microbiota, (v) the ability to inhibit bacterial growth through the production of bacteriocins and other substances, and (vi) immune response activation [
15,
16,
17,
18,
19,
20]. Despite these benefits, live probiotic cells pose certain safety and stability issues that have been highlighted before [
21]. As an alternative to using live cells, it has been demonstrated that non-viable probiotic microorganisms can exert positive effects; these effects include improving growth performance [
22,
23], stimulating the host immune system [
24], and modifying the intestinal microbiota [
25]. In this context, non-viable microbial cells (or crude cell extracts) that confer positive effects on the host are referred to as “paraprobiotics” [
26]. Paraprobiotics offer attractive advantages when it comes to industrial use. They make feed processing easier because they can be introduced before thermal procedures while still maintaining the level of activity necessary to provide the desired health benefits [
27]. Paraprobiotics might also result in easier storage and administration, a longer shelf life, and provide supplementation for those with impaired immune systems [
28,
29].
In a previous work, Medina et al. [
30] selected probiotics based on their ability to generate antibodies that cross-react with two pathogens (
Vibrio harveyi and
Photobacterium damselae subsp.
piscicida) that are present in farmed Senegalese sole (
Solea senegalensis). Among the probiotics used, the
Vibrio proteolyticus DCF12.2 strain stood out for its great capacity to generate antibodies in fish. It also had other beneficial characteristics, such as its ability to inhibit pathogen growth, avirulence in fish, and survivability under storage conditions [
30]. Furthermore, this species showed enzymatic activity, including the production of lecithinase, gelatinase, caseinase, amylase, and lipase, which can contribute to better aquafeed digestibility; continued viability after being added to the aquafeed; and survivability under the pH conditions found in the fish gut [
31]. All the above features make this bacterium an excellent potential candidate for use in aquaculture.
The culture of Mugilidae species (mullets) is considered a priority within current European aquaculture strategies [
32].
Mugil cephalus (Linnaeus, 1758),
Chelon ramada (Risso, 1827),
Chelon labrosus (Risso, 1827), and
Chelon saliens (Risso, 1810) are the main Mugilidae fish species represented among the Mediterranean, each of which has great economic value [
33]. One of the mullet species that can potentially be used in aquaculture is the thick-lipped grey mullet (
C. labrosus). It is omnivorous in the early stages of development, becoming herbivorous with age [
34]. Similar to other mullet species,
C. labrosus has a high osmoregulatory capacity, allowing it to live in a wide range of salinities without compromising its development rate [
35,
36].
In a recent study by our group [
37], we confirmed the suitability of using
C. fusca as a dietary ingredient for
Chelon labrosus juveniles. In the study, a diet containing 15%
C. fusca enhanced the growth performance, nutrient utilization, flesh lipid quality, digestive and metabolic enzyme activity, absorptive capacity, and gut morphology of the fish.
The present study aimed to evaluate the dietary administration of C. fusca and ethanol-inactivated V. proteolyticus DCF12.2 in C. labrosus juveniles, highlighting the nutritional, physiological, and morphological effects. In addition, a comparative approach was also evidenced, in which C. fusca alone was included in the aquafeeds.
4. Discussion
Microalgae and bacteria have been assessed separately as functional ingredients in feed for various fish species. However, little is known about the effects of combining them in aquaculture species. Studies on the combined effect of microalgae and probiotics mainly focus on immune responses, gene expression, intestinal morphology, and the microbiota [
57,
58,
59].
The results of the present study have shown that the combined inclusion of
C. fusca and
V. proteolyticus in
C. labrosus diets significantly promoted growth performance and feed utilization compared to the control group (a non-supplemented diet). High growth performance was also reported by Reyes-Becerril et al. [
60] after 8 weeks of feeding a combination of the microalgae
Navicula sp. and the probiotic
Lactobacillus sakei to Pacific yellow snapper (
Lutjanus peru). Our research group recently investigated the effects of
C. fusca in
C. labrosus diets, finding increased growth performance in the fish receiving the microalgae-supplemented diet [
37]. In this regard, mixing microalgae and bacteria enhanced feed efficiency (FCR and PER) and the fish condition factor, which remained unaltered when
C. fusca was incorporated separately in aquafeeds [
37]. This difference may be due to enzymatic activity (lecithinase, gelatinase, caseinase, amylase, and lipase) in
V. proteolyticus whole cells [
30], which continues after ethanol inactivation. This activity might allow the host to better digest the aquafeed and improve the availability of nutrients. This agrees with the study by De Schrijver and Ollevier [
61], which reported that
V. proteolyticus ingestion stimulates protein digestibility in turbot (
Scophthalmus maximus).
Muscle proximate composition is considered an indicator of the physiological and health status of fish [
62]. Despite the fact that none of the dietary treatments tested in this work modified the muscle composition, it is known that the inclusion of probiotic microorganisms and microalgae in aquafeeds may exert a significant effect on the chemical composition of the animal. Authors such as Pandey et al. [
63] observed an increase in muscle protein content in common carp (
Cyprinus carpio) specimens fed
Lactobacillus plantarum-supplemented diets, which seems to be associated with improved feed efficiency rates. Meanwhile, microalgae inclusion is related to fish metabolism, especially with lipids used as an energy source, which lead to a reduction in lipid tissue storage [
64].
Regarding the fatty acid composition, dietary changes are reflected in the fatty acid composition of marine fish tissues [
65]. As expected, the fatty acid profiles of experimental feeds were reflected in an increased proportion of 16:0, 18:1n9, and 18:2n6 in the muscle of both groups. Furthermore, the C + V diet induced a selective retention of ARA, EPA, and DHA in the muscle, which agrees with findings from other marine fish species fed with algae-supplemented diets [
66,
67,
68]. This points to a relationship between the algae inclusion level and the higher efficiency of mobilizing lipids. Inadequate intake of these fatty acids, which are essential for the cellular membrane structure and function, results in decreased growth and increased fish mortality, as well as other pathologies, such as liver or intestinal steatosis [
69]. The relative increase in structural fatty acids (ARA, EPA, and DHA) in the muscle is also reflected in the significant increase in the fish lipid quality index (FLQ), n-3 polyunsaturated fatty acid, and n-3/n-6 ratio in the fish fed the C + V diet, which could be beneficial from a human nutrition standpoint [
70,
71].
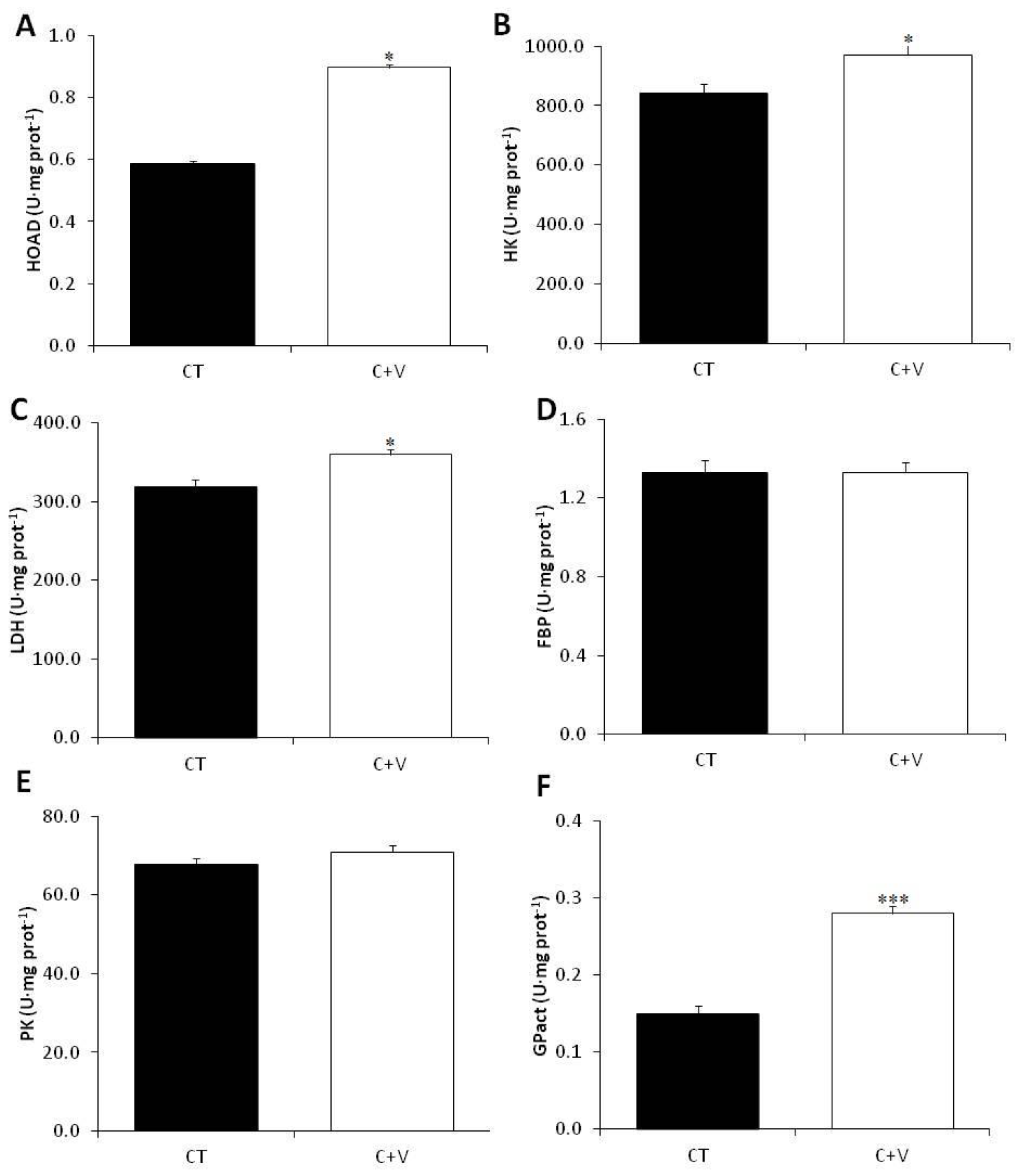
The metabolic response to the experimental diets was also investigated. The C-V diet considerably activated the carbohydrate metabolism by statistically boosting the plasma glucose and lowering (not statistically) the plasma lactate. Thus, we propose that the C + V diet may promote plasma lactate depletion, which originates in the white muscle due to anaerobic metabolism and is then partially incorporated in the liver; this is consistent with the findings of other authors [
37,
49]. Furthermore, a significant increase in LDH activity was found in fish fed the C + V diet; this is consistent with prior research in which gilthead seabream were fed diets enriched with microalgae [
72]. This increase in LDH activity appears to indicate the conversion of lactate to pyruvate via the Cori cycle, which may then be converted and stored as glycogen or for energy within the liver or elsewhere.
Hepatic TAG levels increased in the C + V fed fish compared to those fed the control diets. The liver is the primary site of lipogenesis in fish, and hepatic carbohydrates may regulate the total lipogenesis [
73]. Because both metabolites, glycogen and TAG, served as energy reserves, our findings imply that more energy is invested in maintenance and development than in storage, which is consistent with the biometric data obtained, and those reported by other authors [
72].
It was also shown that fish fed the C + V diet had significantly increased hepatic HK activity. Nevertheless, the PK activity remained unchanged, which is consistent with the findings of Perera et al. [
49] and our earlier work evaluating
C. fusca as a dietary ingredient for
C. labrosus [
37]. HK catalyzes the initial stage of glycolysis, phosphorylating glucose so that it can be used by the cells, whereas PK catalyzes the last step, creating pyruvate and ATP. Interestingly, Perera et al. [
49] found that 1% microalgae supplementation increased the hepatocyte capacity for glucose uptake and glycogen storage but not the oxidation for energy; this differs from the findings of Molina-Roque et al. [
72], who found that higher inclusion (10%) in identical diets to those used by Perera et al. [
49] increased the oxidation for energy. Although our work and the studies mentioned above were conducted in two different species, it appears that the inclusion of microalgae-derived products in aquafeeds consistently promotes glucose uptake by hepatocytes. Nonetheless, the final fate (i.e., storage or oxidation) varies depending on the level of microalgae inclusion. Furthermore, the studies by García-Márquez et al. [
37] and Molina-Roque et al. [
72] showed a decrease in hepatic gluconeogenesis-related FBP enzyme activity [
74], supporting the glucose oxidation hypothesis. In our research, however, the hepatic FBP levels were unchanged.
Our results also show a significant increase in GPase activity in fish fed the C + V diet. Using glycogen as a substrate, this enzyme catalyzes the dephosphorylation of glucose-6-phosphate to glucose [
75]. Thus, the increased enzyme activity correlates well with the reduction in hepatic glycogen reported in the C + V-fed fish. Lastly, fish fed the C + V diet had significantly increased HOAD activity. The HOAD enzyme is involved in fatty acid β-oxidation [
76], and its activity is directly related to lipid availability [
77]. This finding may be linked to the observed increased in hepatic glucose oxidation for energy, which would enhance triglyceride levels in the liver, making them more accessible to promote muscle development, as postulated by Molina-Roque et al. [
72].
On the other hand, the evaluation of digestive enzymes and the knowledge of their involvement in the digestion and absorption processes are key tools that can be used as a reliable indicator of the nutritional status of aquaculture fish, as well as to select new ingredients to use in aquafeed production [
50]. Previous works have shown that using both probiotic microorganisms and microalgae induces significant changes in some of the enzymes involved in digestion and absorption processes, increasing the activity levels of both pancreatic and brush border secretion enzymes [
14,
37,
55]. Overall, the results from the assays in this study indicate that the dietary inclusion of
C. fusca together with
V. proteolylicus increased the enzyme activity assayed in fish fed the control diet. It is widely known that higher secretion of digestive enzymes induces an improvement in protein digestion, which contributes to better feed utilization [
55]. This could explain the positive effects observed on growth and the feed utilization rate derived from the inclusion of microalgae and probiotics in feed.
It is also worth mentioning the favorable effects of the
C. fusca +
V. proteolyticus-supplemented diet on brush border digestive activity (leucine aminopeptidase and alkaline phosphatase) that we observed in this work. These enzymes, especially alkaline phosphatase, a dominant enzyme in the intestinal brush border, can be used as an indicator of intestinal integrity and nutrient absorption [
78,
79]. Indeed, these positive effects concur with the histological and ultrastructural determinations of the intestinal mucosa.
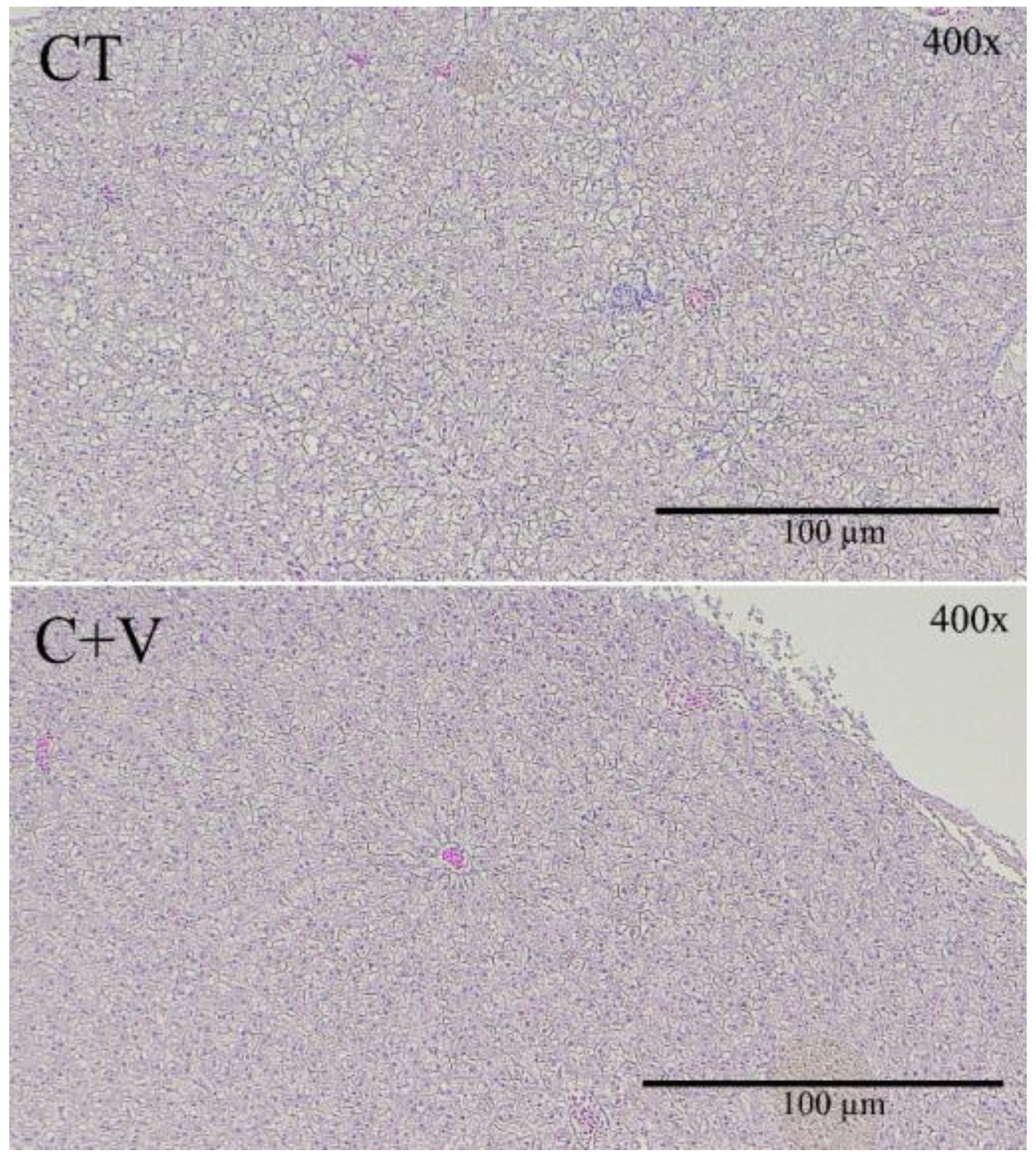
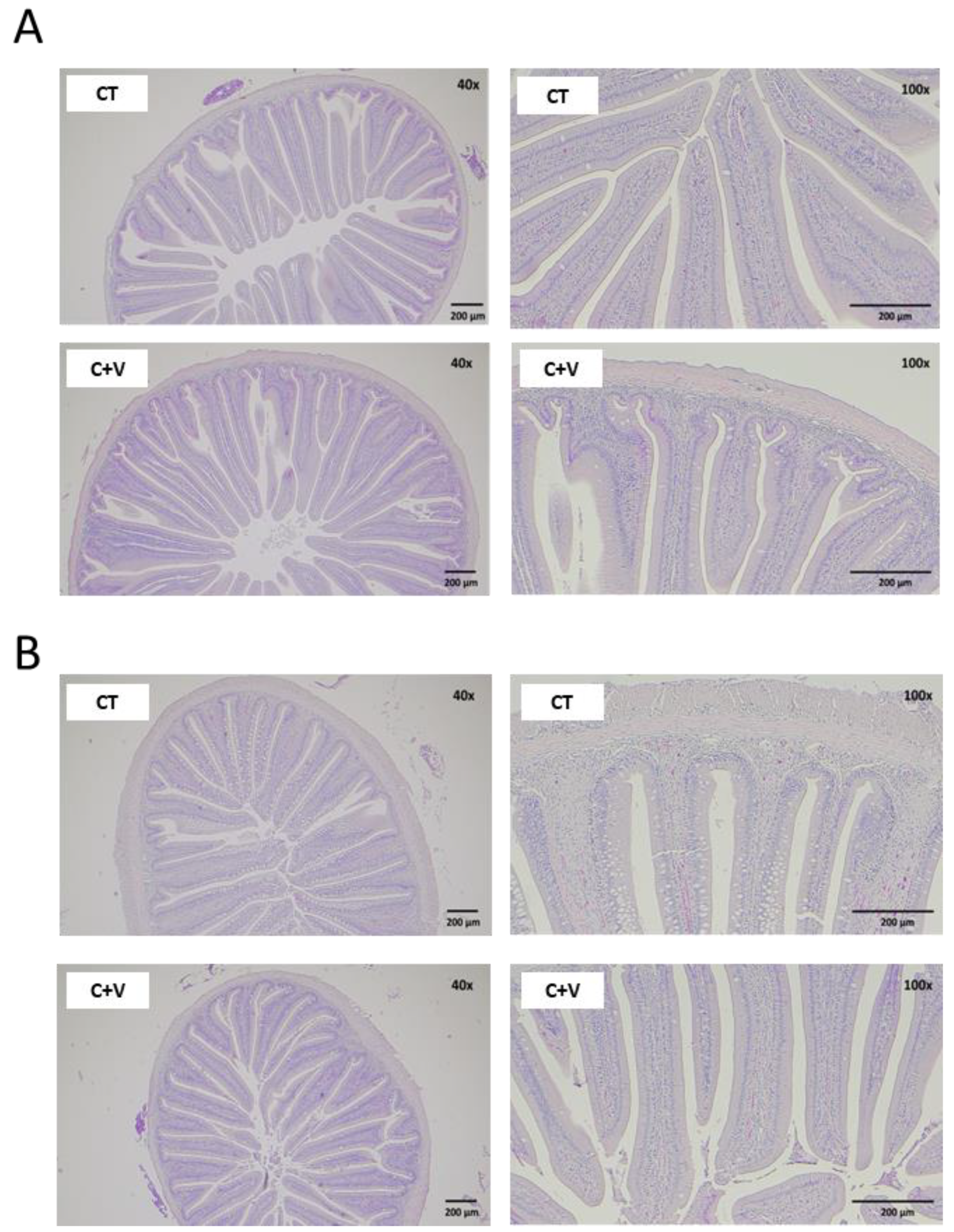
A healthy gut is essential for optimal animal performance [
55,
80], so any alteration in the intestinal mucosa’s integrity strongly activates immune cells and may cause chronic inflammation of the intestinal tissue. There is evidence to confirm that the use of probiotic microorganisms and microalgae in aquafeeds seems to have a positive effect on the gut morphology of various fish species [
68,
81,
82]. In our study, although an increase in the mucosal fold length was observed in the fish fed the
C. fusca and
V. proteolyticus-supplemented diet, a significant reduction in the thickness of the serosa and submucosa layer was also observed in comparison to the control group. It is important to note that the dietary inclusion of microalgae and probiotics induced a marked reduction in the level of vacuolization observed in the distal intestine of fish fed the control diet, as well as in the thickness of the lamina propria. In this regard, some studies carried out on zebrafish pointed out that certain microalgae (
Tetraselmis sp.;
Phaeodactylum tricornutum;
Chlorella sp.;
Nannochloropsis oculata or
Nannochloropsis gaditana) and probiotic microorganisms are efficient in reducing the intestinal inflammatory response [
83,
84].
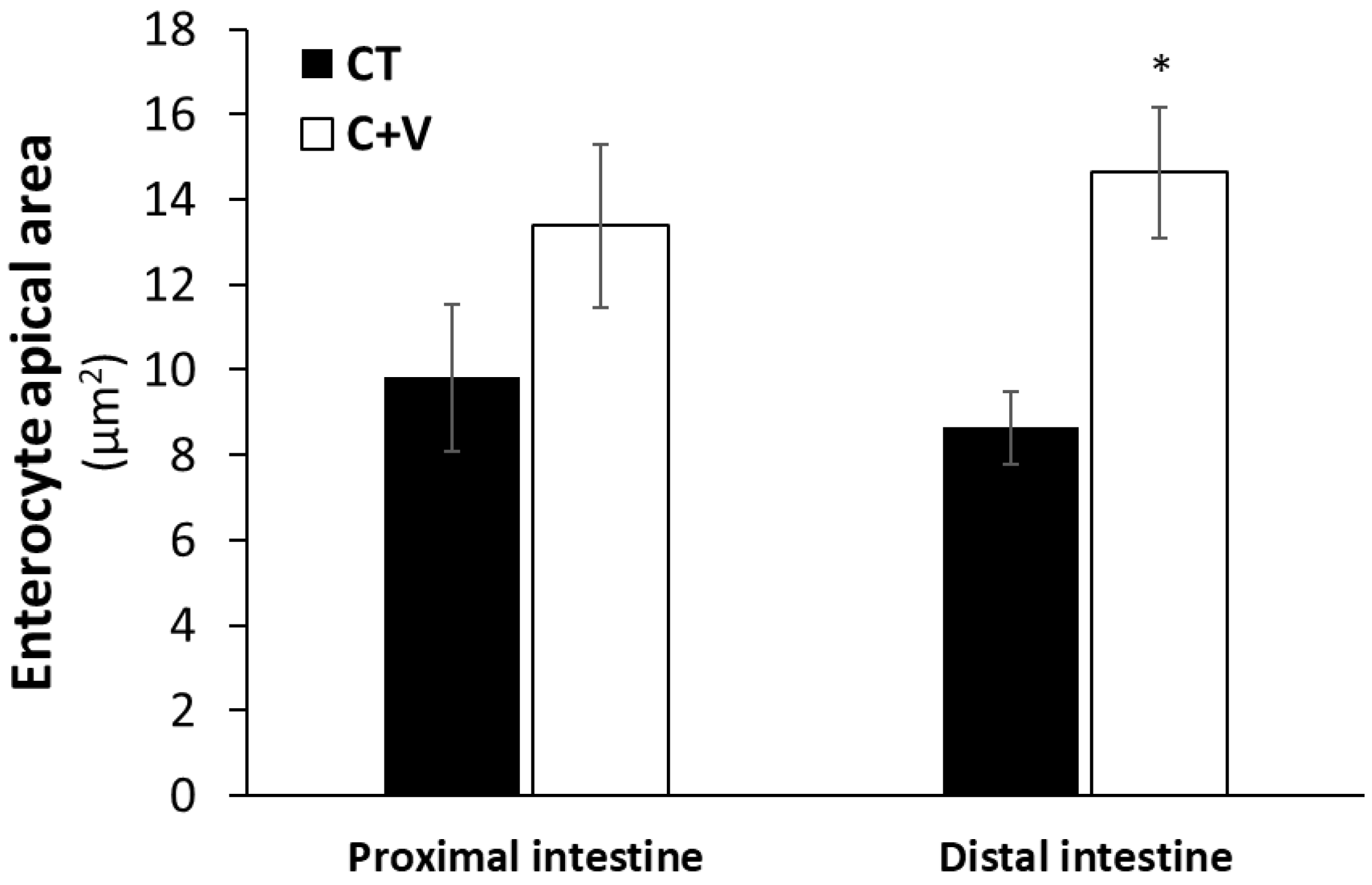
Similarly, the scanning electron microscopy analysis showed no signs of damage in the apical brush border. In agreement with that observed in previous works carried out on a variety of marine fish species (gilthead seabream, Senegalese sole, thick-lipped grey mullet) at different developmental stages (fry and juvenile) [
37,
64,
68], the inclusion of microalgae in aquafeeds exerted positive changes in the enterocyte apical area, which can translate into an overall increase in the enterocyte absorption surface and, consequently, an enhanced intestinal absorption capacity [
55].
In order to elucidate whether the positive effects found in this work are attributable to the microalgae alone or the combined use of the microalgae and the inactivated bacteria, we compared the growth performance, nutrient utilization, and the n-3 long-chain polyunsaturated fatty acids of the muscle samples from the current experiment with the results obtained by García-Márquez et al. [
37]. The results showed that there were no differences between the two experimental diets (C-15 from García-Márquez et al. [
37] and C + V from the present study) (
Table S2,
Figure S1), despite the fact that the experiments were carried out on fish of different sizes and over different photoperiods. Although the values of polyunsaturated fatty acids in the experimental diets were not as high as the values obtained in wild fish (García-Márquez et al. [
56]), they were significantly higher than in fish fed the control feeds. This demonstrates that the use of microalgae in the diet can be a good strategy for increasing the polyunsaturated fatty acids in the flesh of farmed fish, bearing in mind that these fatty acids are important for human nutrition. In this regard, the Food and Agriculture Organization (FAO), and the European Food Safety Authority (EFSA) recommend a daily EPA and DHA intake of at least 250 mg [
85,
86]. Both the diets that included microalgae (C-15 from García-Márquez et al. [
37] and C + V from the present study) were higher than the reference value for good health, indicating that the specimens can be considered a good source of EPA and DHA in human diets.
The similar values obtained with both diets show that the inclusion of the inactivated bacteria did not affect the growth and lipid composition of the fish. However, the inclusion of inactivated bacteria in the experimental diet may have affected the enterocyte morphology and the disappearance of vacuoles in the enterocytes compared to fish fed the control diet. In this sense, García de La Banda et al. [
87] observed that the inclusion of the probiotic
Shewanella putrefaciens pdp11 in the diet of
Solea senegalensis caused the removal of the lipid droplets inside the enterocytes of those fish compared to fish fed the control feed.

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}