
A Comparative Study on the Distribution Pattern of Endocrine Cells in the Gastrointestinal Tract of Two Small Alpine Mammals, Plateau Zokor (Eospalax baileyi) and Plateau Pika (Ochotona curzoniae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
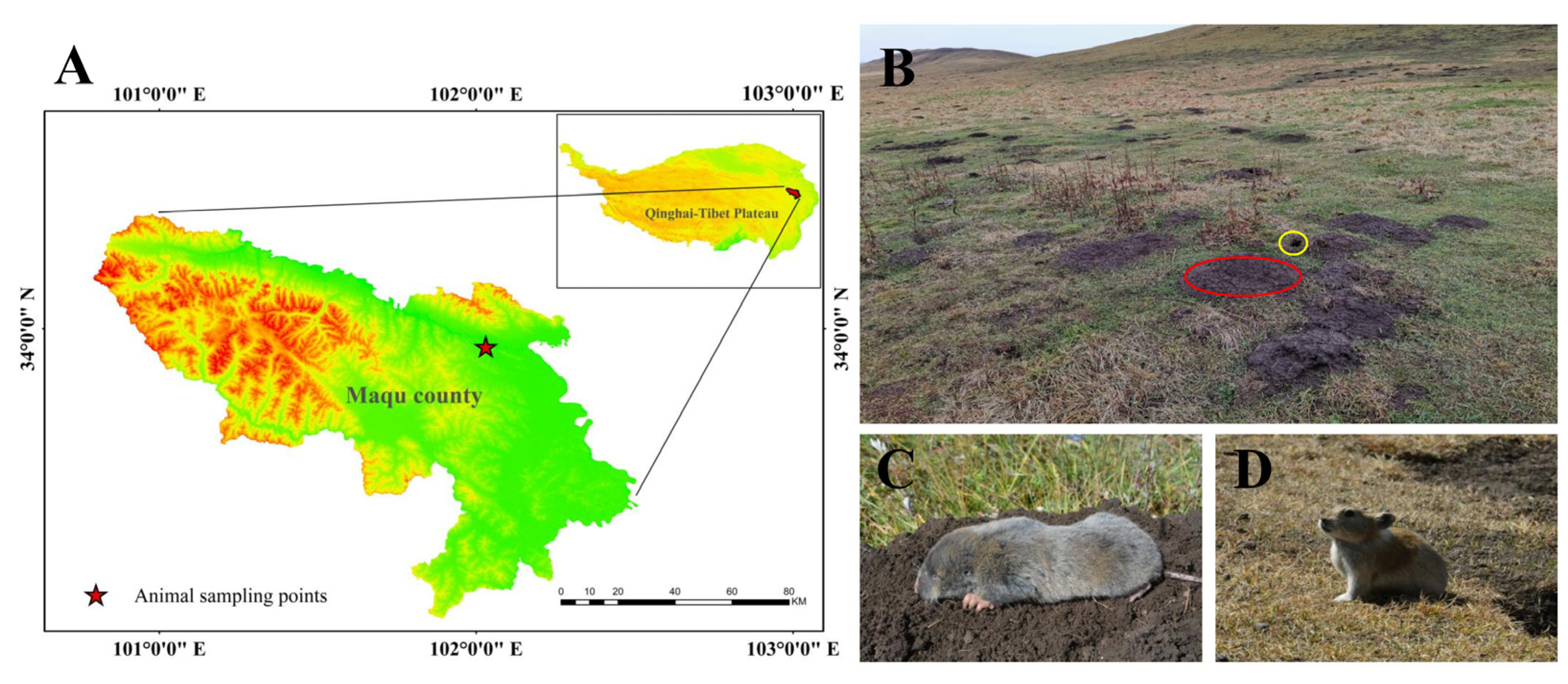
2.1. Experimental Animals and Their Habitats
2.2. Tissue Preparation
2.3. Histochemical Study
2.4. Immunohistochemical Study
2.5. Data Analysis
3. Results
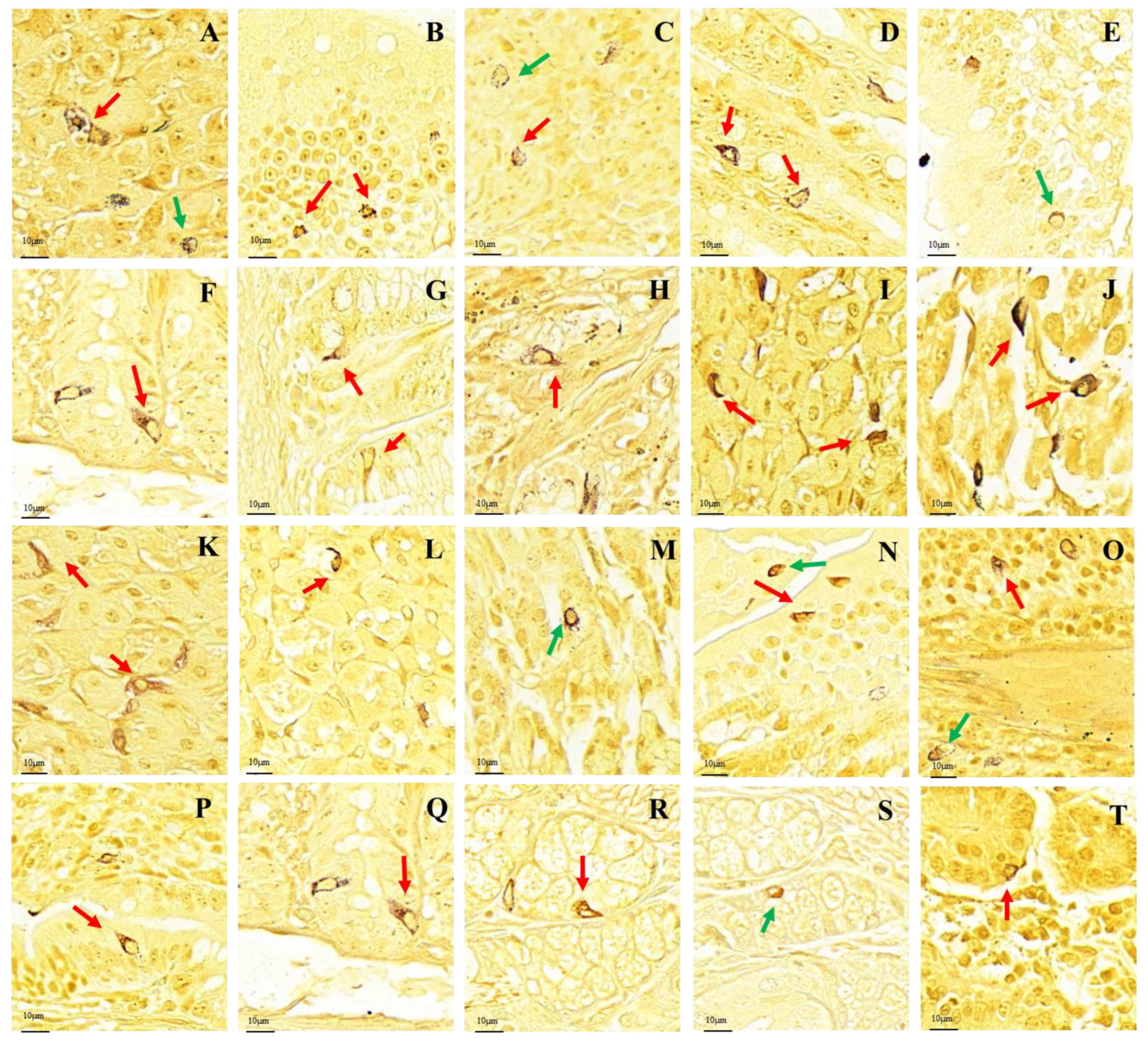
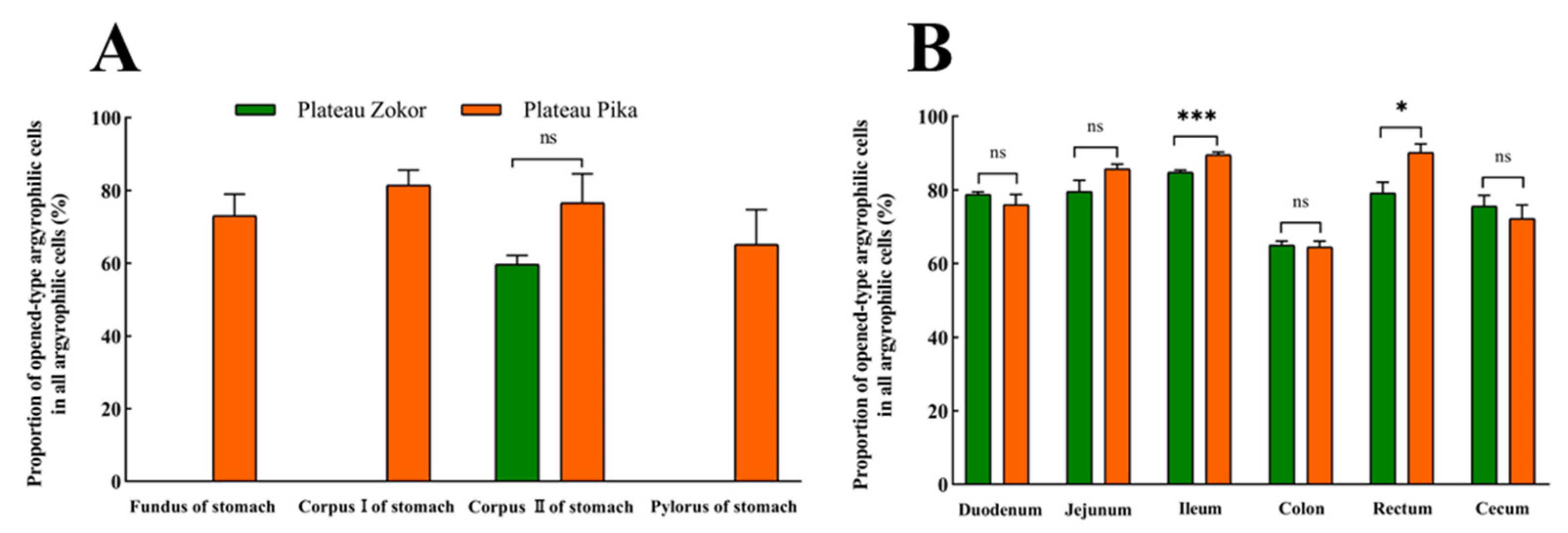
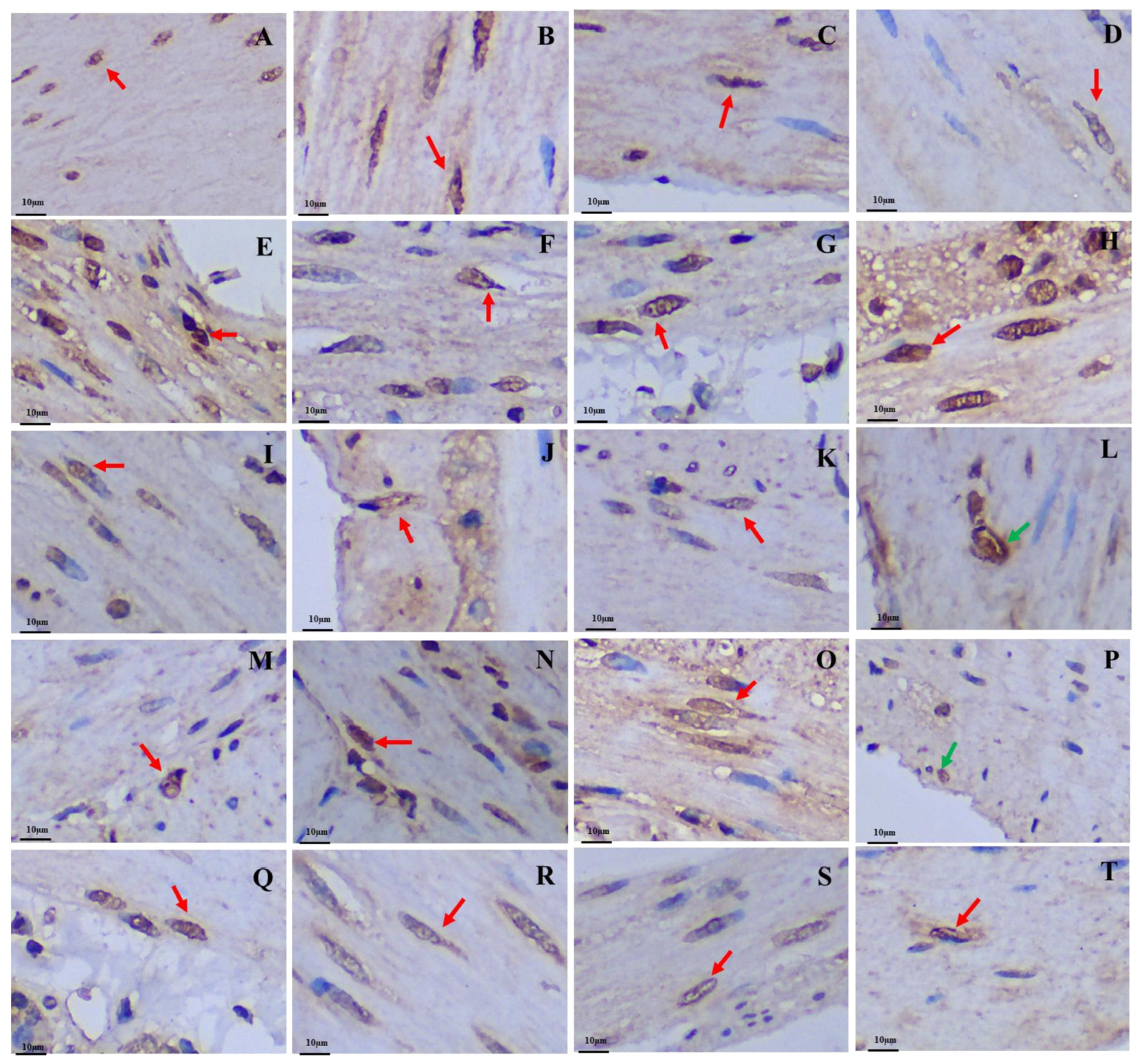
3.1. Distribution Pattern of Gastrointestinal Argyrophilic Cells
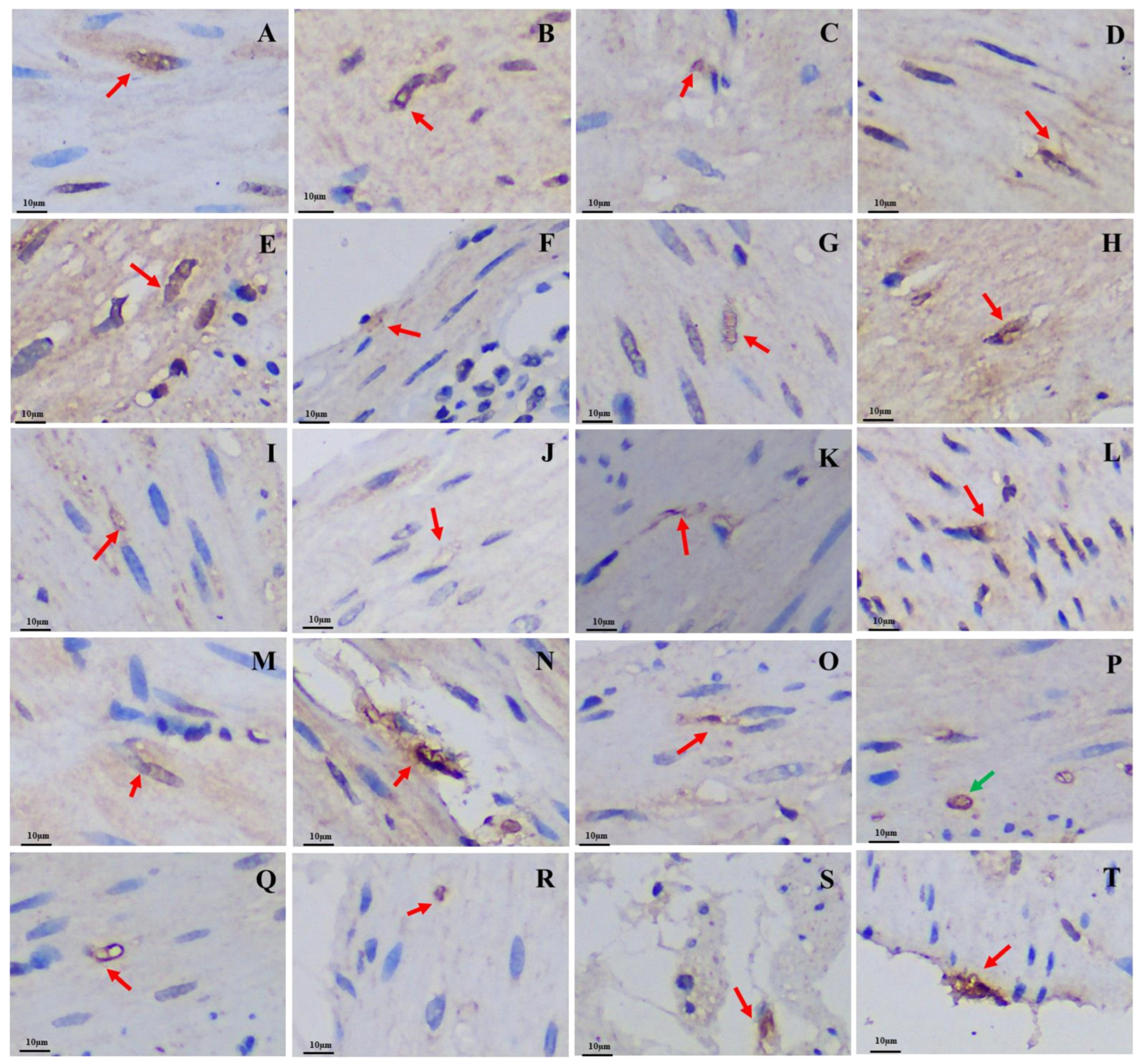
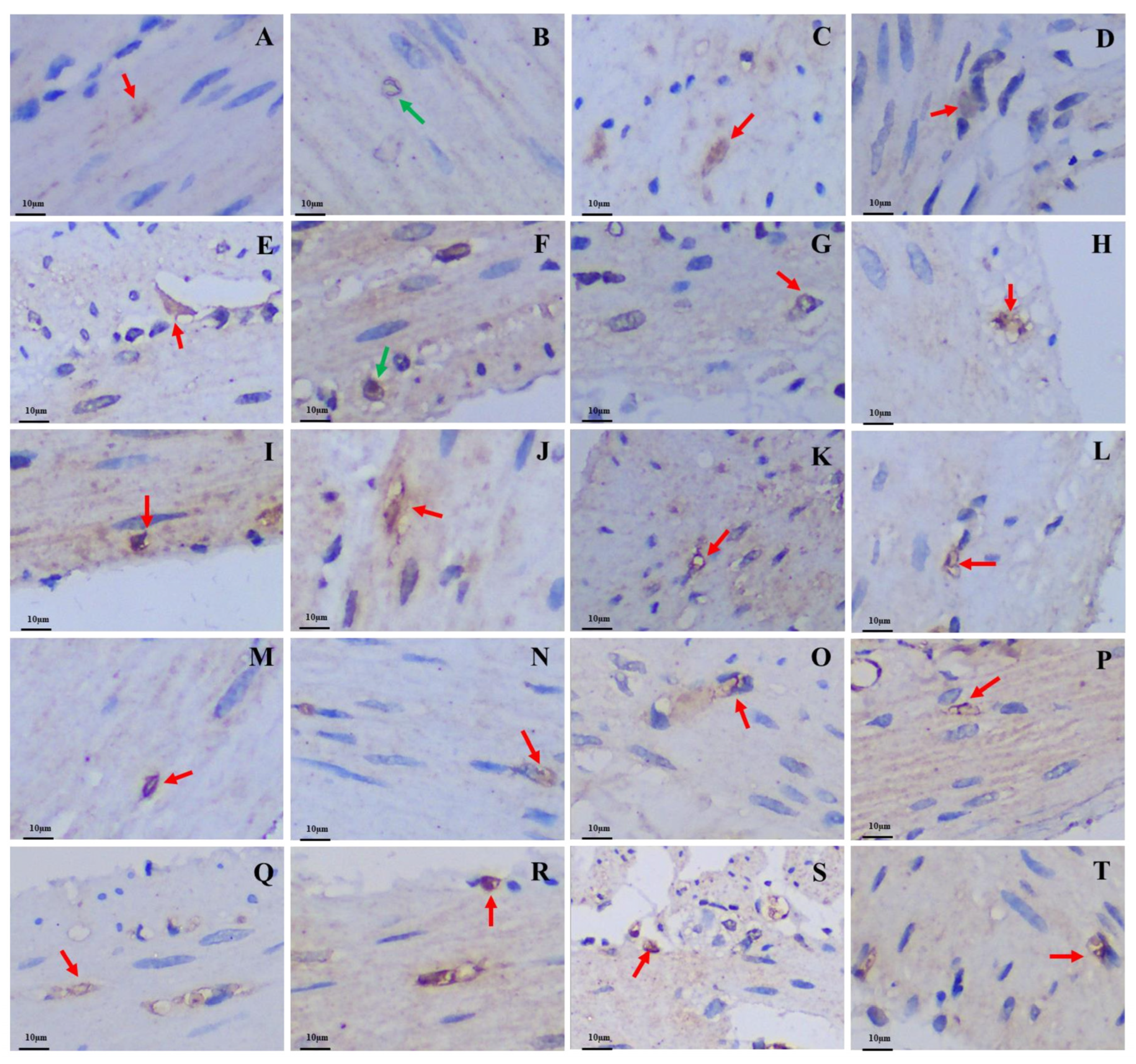
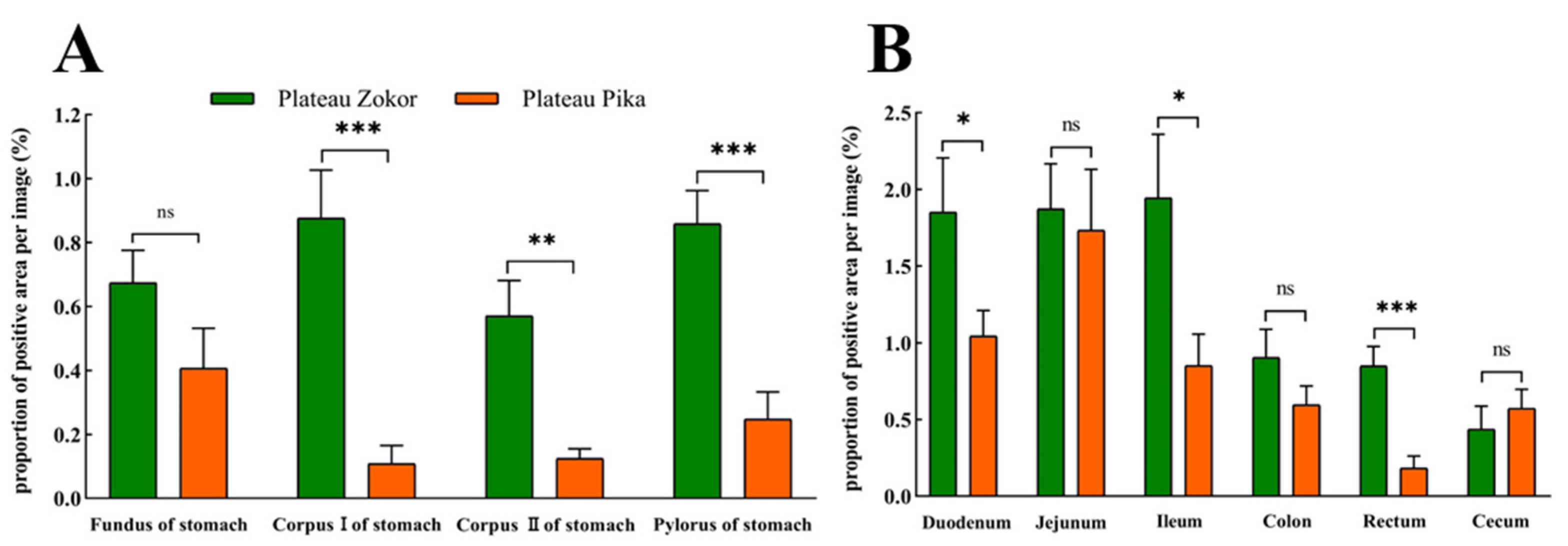
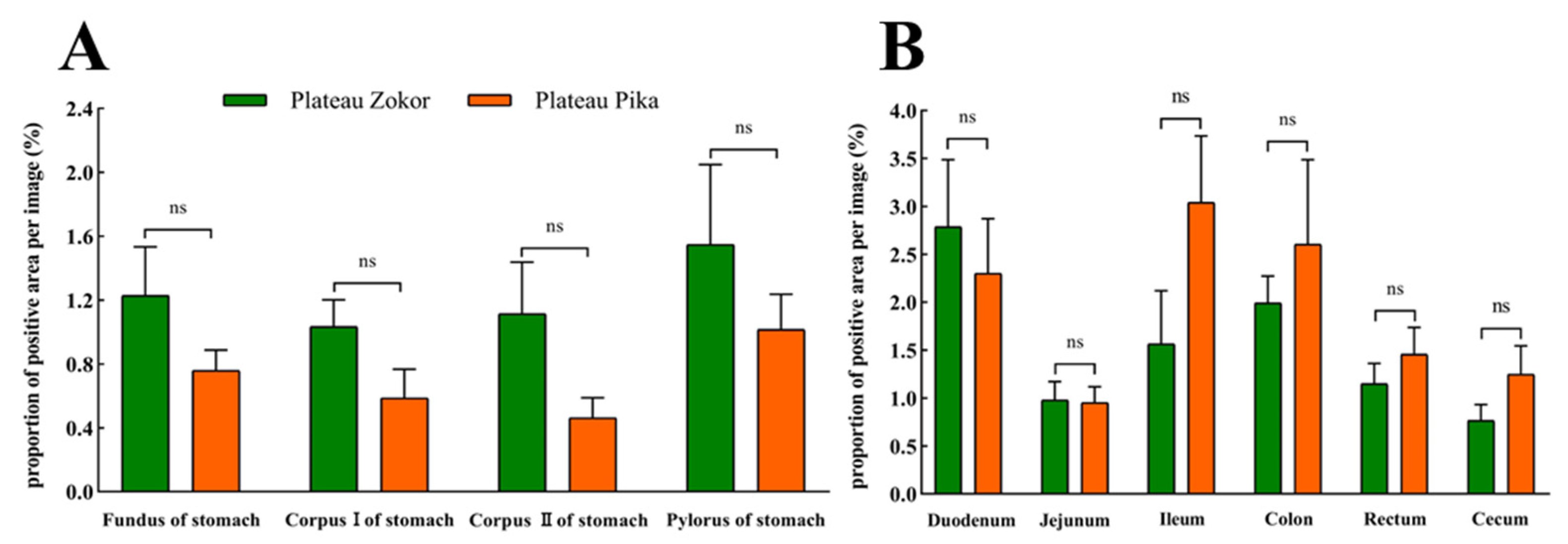
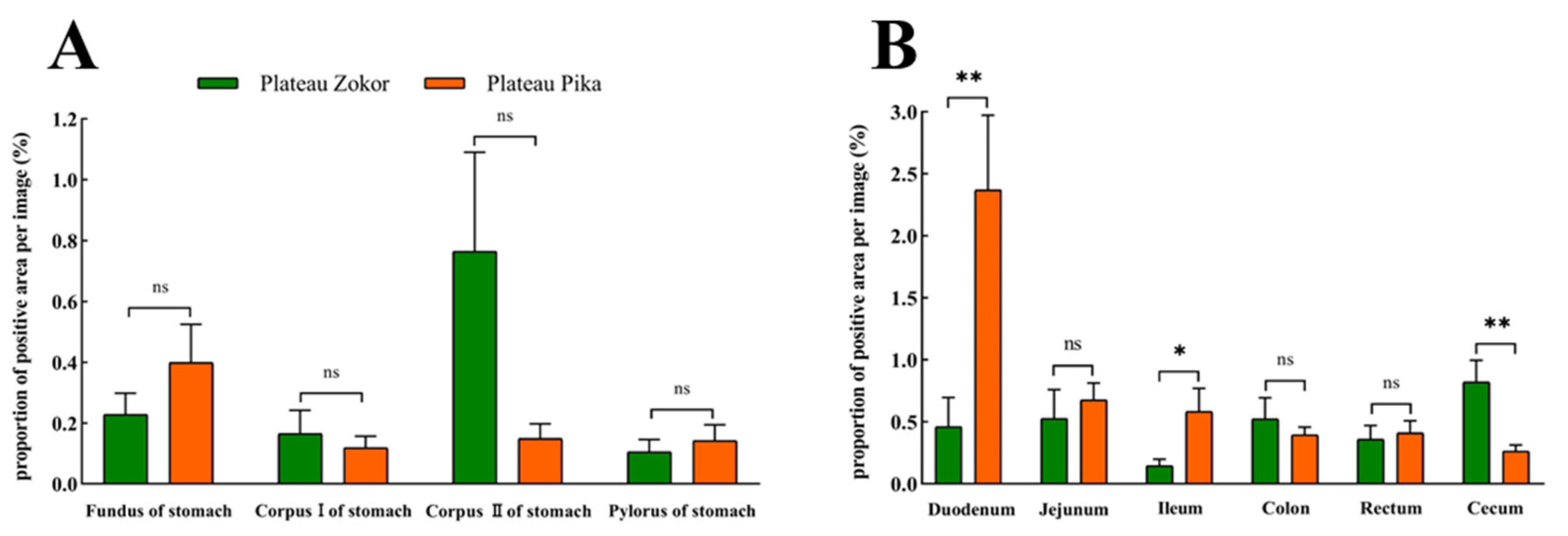
3.2. Differences in the Positive Expression of Gastrointestinal Immunoreactive Cells
3.2.1. 5-HT-Immunoreactive Cells
3.2.2. Gas-Immunoreactive Cells
3.2.3. Glu-Immunoreactive Cells
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Meerveld, G.V.; Johnson, A.C.; Grundy, D. Gastrointestinal Physiology and Function. In Gastrointestinal Pharmacology; Meerveld, G.V., Ed.; Springer: Cham, Germany, 2017; Volume 239, pp. 1–16. [Google Scholar]
- Monteiro, M.P.; Batterham, R.L. The importance of the gastrointestinal tract in controlling food intake and regulating energy balance. Gastroenterology 2017, 152, 1707–1717.e1702. [Google Scholar] [CrossRef]
- Parker, H.E.; Gribble, F.M.; Reimann, F. The role of gut endocrine cells in control of metabolism and appetite. Exp. Physiol. 2014, 99, 1116–1120. [Google Scholar] [CrossRef] [PubMed]
- Rezzani, R.; Franco, C.; Franceschetti, L.; Gianò, M.; Favero, G. A Focus on Enterochromaffin Cells among the Enteroendocrine Cells: Localization, Morphology, and Role. Int. J. Mol. Sci. 2022, 23, 3758. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.X.; Li, P.; Zhang, X.T.; Ye, L.X. Distribution and developmental changes of ghrelin-immunoreactive cells in the pancreas of African ostrich chicks (Struthio camelus). Poultry Sci. 2017, 96, 3445–3451. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.T.; de Freitas, T.R.; de Oliveira, I.R.C.; Costa, L.S.; Vigliano, F.A.; Rosa, P.V. Endocrine cells producing peptide hormones in the intestine of Nile tilapia: Distribution and effects of feeding and fasting on the cell density. Fish Physiol. Biochem. 2017, 43, 1399–1412. [Google Scholar] [CrossRef] [PubMed]
- Matsumoto, M.; Takemi, S.; Sakai, T.; Sakata, I. Identification of motilin in Japanese fire bellied newt. Gen Comp Endocr 2022, 323, 114031. [Google Scholar] [CrossRef]
- Guo, X.T.; Lv, J.Y.; Xi, R.W. The specification and function of enteroendocrine cells in Drosophila and mammals: A comparative review. FEBS J. 2022, 289, 4773–4796. [Google Scholar] [CrossRef]
- Pereira, R.T.; Nebo, C.; de Paula Naves, L.; Fortes-Silva, R.; Regina Cardoso de Oliveira, I.; Paulino, R.R.; Drummond, C.D.; Rosa, P.V. Distribution of goblet and endocrine cells in the intestine: A comparative study in Amazonian freshwater Tambaqui and hybrid catfish. J. Morphol. 2020, 281, 55–67. [Google Scholar] [CrossRef] [PubMed]
- El-Salhy, M.; Mazzawi, T.; Hausken, T.; Hatlebakk, J.G. Interaction between diet and gastrointestinal endocrine cells. Biomed. Rep. 2016, 4, 651–656. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, S.; Zhou, N.; Wang, C.; Wu, X. Distribution of endocrine cells in the digestive tract of Alligator sinensis during the active and hibernating period. Tissue Cell 2014, 46, 343–351. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.Y.; Qi, Y.; Wang, X.N.; Zhao, W. Histology of and argyrophil cells in digestive tract of Phrynocephalus vlangalii. Acta Ecol. Sin. 2020, 40, 5855–5861. [Google Scholar]
- Wang, J.X.; Peng, K.M.; Liu, H.Z.; Song, H.; Chen, X.; Min, L. Distribution and morphology of argyrophilic cells in the digestive tract of the African ostrich. Tissue Cell 2010, 42, 65–68. [Google Scholar] [CrossRef]
- Dos Santos, D.C.M.; do Carmo Cupertino, M.; Novaes, R.D.; da Costa Soares, Í.A.; Fonseca, C.C.; da Matta, S.L.P.; Sartori, S.S.R. Morphologic characterization and distribution of endocrine cells in the large intestine of the opossum Didelphis aurita (Wied-Neuwied, 1826). Tissue Cell 2013, 45, 338–349. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.M.; Chen, X. Effect of Cyclic Starvation on the Morphology and Distribution of Argyrophilic Cells in the Digestive Tract of Rats. Chin. J. Zool. 2020, 55, 222–228. [Google Scholar]
- Zhao, S.Q.; He, M.; Liang, X.X.; Li, X.; Liao, L. Effects of the different processed products from Terminalia chebula Retz on the morphology and distribution of argentaffin cells in the digestive tract of mice. Chin. Vet. Sci. 2016, 46, 1048–1056. [Google Scholar]
- Gribble, F.M.; Reimann, F.; Roberts, G.P. Gastrointestinal Hormones. In Physiology of the Gastrointestinal Tract, 6th ed.; Said, H.M., Ed.; Academic Press: Cambridge, MA, USA, 2018; pp. 31–70. [Google Scholar]
- Nathália, D.N.C.; da Silveira Firmiano, E.M.; Gomes, I.D.; do Nascimento, A.A.; Sales, A.; Araújo, F.G. Histochemical and immunohistochemical study on endocrine cells (5HT, GAS, and SST) of the gastrointestinal tract of a teleost, the characin Astyanax bimaculatus. Acta Histochem. 2015, 117, 595–604. [Google Scholar]
- Andersen, D.B.; Holst, J.J. Peptides in the regulation of glucagon secretion. Peptides 2022, 148, 170683. [Google Scholar] [CrossRef] [PubMed]
- Gershon, M.D. 5-Hydroxytryptamine (serotonin) in the gastrointestinal tract. Curr. Opin. Endocrinol. 2013, 20, 14. [Google Scholar] [CrossRef]
- Rehfeld, J.F. Gastrin and the moderate hypergastrinemias. Int. J. Mol. Sci. 2021, 22, 6977. [Google Scholar] [CrossRef] [PubMed]
- Chu, B.; Ji, C.P.; Zhou, J.W.; Zhou, Y.S.; Hua, L.M. Why does the plateau zokor (Myospalax fontanieri: Rodentia: Spalacidae) move on the ground in summer in the eastern Qilian Mountains? J. Mammal. 2021, 102, 346–357. [Google Scholar] [CrossRef]
- Nie, H.; Liu, J.; Chen, B. Life-history traits and fitness of plateau pika (Ochotona curzoniae) in alpine meadow ecosystem. Ecol. Evol. 2022, 12, e8548. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.Q.; An, Z.F.; Wei, L.N.; Zhang, J.Y.; Li, J.M.; Wang, Z.J.; Gao, C.H.; Wei, D.B. Vitamin D3 Metabolic Enzymes in Plateau Zokor (Myospalax baileyi) and Plateau Pika (Ochotona curzoniae): Expression and Response to Hypoxia. Animals 2022, 12, 2371. [Google Scholar] [CrossRef] [PubMed]
- Cai, X.C.; Bao, D.; Ye, G.H.; Chu, B.; Tang, Z.S.; Hua, R.; Hua, L.M. The Complex and Well-Developed Morphological and Histological Structures of the Gastrointestinal Tract of the Plateau Zokor Improve Its Digestive Adaptability to High-Fiber Foods. Animals 2022, 12, 2447. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Tian, H.; Yu, Z.; Zhao, H.; Li, W.; Sang, D.; Lin, K.; Cui, Y.; Liao, M.; Xu, Z. A highland-adaptation mutation of the Epas1 protein increases its stability and disrupts the circadian clock in the plateau pika. Cell Rep. 2022, 39, 110816. [Google Scholar] [CrossRef]
- Ye, G.; Chu, B.; Zhou, R.; Zhang, F.; Hua, X.; Hua, R.; Wang, T.; Dong, K.; Hua, L. Relationship between plant functional group distribution and soil properties under the disturbance of plateau pika (Ochotona curzoniae). Chin. J. Ecol. 2019, 38, 2382–2388. [Google Scholar]
- Zhang, C.J.; Yao, B.H.; Wang, X.Y.; Sun, X.M.; Su, J.H. Effects of grassland sowing on the feeding habits of Eospalax baileyi in the Gannan alpine meadow. Pratacult. Sci. 2021, 38, 967–975. [Google Scholar]
- Hou, C.; Shang, G.Z.; Wu, X.Q.; Cao, Y.F.; Chen, J.H.; Bian, J.H. Comparative studies on food habit analysis of gastric contents and colon contents of plateau pika (Ochotona curzoniae). Acta Theriol. Sin. 2019, 39, 662–669. [Google Scholar]
- Grimelius, L.; Wilander, E. Silver stains in the study of endocrine cells of the gut and pancreas. Investig. Cell. Pathol. 1980, 3, 3–12. [Google Scholar]
- Hsu, S.M.; Raine, L.; Fanger, H. Use of avidin-biotin-peroxidase complex (ABC) in immunoperoxidase techniques: A comparison between ABC and unlabeled antibody (PAP) procedures. J. Histochem. Cytochem. 1981, 29, 577–580. [Google Scholar] [CrossRef]
- Gribble, F.M.; Reimann, F. Enteroendocrine Cells: Chemosensors in the Intestinal Epithelium. Annu. Rev. Physiol. 2016, 78, 277–299. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.T.; Yin, C.; Yang, F.; Zhang, Y.C.; Huang, H.W.; Wang, J.W.; Deng, B.W.; Cai, T.; Rao, Y.; Xi, R.W. The cellular diversity and transcription factor code of Drosophila enteroendocrine cells. Cell Rep. 2019, 29, 4172–4185.e4175. [Google Scholar] [CrossRef]
- Sartori, S.S.R.; Peixoto, J.V.; Lopes, V.d.P.G.; Barbosa, A.J.A.; Neves, C.A.; Fonseca, C.C. Neuroendocrine structures of the small intestine of the capybara Hydrochoerus hydrochaeris (Mammalia, Rodentia). Anim. Biol. 2018, 68, 89–104. [Google Scholar] [CrossRef]
- Barbosa Lopes, F.; Assis Rodrigues, M.L.; Souza Silva, C.H.d.; Souza Rodrigues Sartori, S. Identification of enteroendocrine cells of Tropidurus torquatus and Salvator merianae (Squamata: Lacertilia). Cuad. Herpetol. 2021, 35, 97–107. [Google Scholar]
- Treuting, P.M.; Arends, M.J.; Dintzis, S.M. Lower gastrointestinal tract. In Comparative Anatomy and Histology; Elsevier: Amsterdam, The Netherlands, 2018; pp. 213–228. [Google Scholar]
- Sahd, L.; Pereira, D.L.; Bennett, N.C.; Kotzé, S.H. Comparative gastrointestinal morphology of Tachyoryctes splendens (rüppell, 1835) and Heliophobius emini,(noack, 1894) two species of East African mole-rats. J. Morphol. 2017, 278, 780–790. [Google Scholar] [CrossRef] [PubMed]
- Walters, J.; Marais, S.; Johnson, O.; Bennett, N.C.; Alagaili, A.N.; Mohammed, O.B.; Kotze, S. The comparative gastrointestinal morphology of five species of muroid rodents found in Saudi Arabia. J. Morphol. 2014, 275, 980–990. [Google Scholar] [CrossRef]
- Valle, J.; Maanes, A.A.L. Digestive flexibility in females of the subterranean rodent ctenomys talarum in their natural habitat. J. Exp. Zool. Part A 2011, 315, 141–148. [Google Scholar] [CrossRef]
- Zhang, C.J.; Wang, X.Y.; Yao, B.H.; Wang, C.; Kang, Y.K.; Zhang, Q.; Su, J.H. Diet Composition and Trophic Niche Characteristics of Three Rodents in Gannan Meadow. Acta Agrestia Sin. 2021, 29, 1484–1490. [Google Scholar]
- Cai, X.C.; Bao, D.; Zhou, R.; MA, Y.J.; Dong, K.C.; Hua, L.M. Comparison of the Gastrointestinal Tract Morphology between Plateau Pika and Plateau Zokor. Acta Agrestia Sin. 2022, 30, 1237–1245. [Google Scholar]
- Speakman, J.R.; Chi, Q.; Ołdakowski, Ł.; Fu, H.; Fletcher, Q.E.; Hambly, C.; Togo, J.; Liu, X.; Piertney, S.B.; Wang, X. Surviving winter on the Qinghai-Tibetan Plateau: Pikas suppress energy demands and exploit yak feces to survive winter. Proc. Natl. Acad. Sci. USA 2021, 118, e2100707118. [Google Scholar] [CrossRef]
- Wang, H.; Zhou, N.Z.; Zhang, R.; Wu, Y.Y.; Zhang, R.D.; Zhang, S.Z. Identification and localization of gastrointestinal hormones in the skin of the bullfrog Rana catesbeiana during periods of activity and hibernation. Acta Histochem. 2014, 116, 1418–1426. [Google Scholar] [CrossRef]
- Zhang, M.; Chen, X. Effects of 8-Week Aerobic Exercise on the Morphology and Distribution Density of Argyrophilic Cells in the Digestive Tract of Obese Rats. Chin. J. Zool. 2020, 55, 752–759. [Google Scholar]
- Wang, H.; Wu, Y.Y.; Zhang, R.; Zhu, X.; Zhang, S.Z. Identification and characterization of gastrointestinal hormone immunoreactive cells in the skin and parotoids of Chinese toad Bufo gargarizans. Folia Histochem. Cytobiol. 2014, 52, 149–156. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Location | Plateau Zokor (Eospalax baileyi) | Plateau Pika (Ochotona curzoniae) | t Value | Significance |
|---|---|---|---|---|
| Fundus of stomach | - | 32.58 ± 2.48 a | - | - |
| Corpus I of stomach | - | 17.28 ± 3.72 b | - | - |
| Corpus II of stomach | 33.70 ± 2.23 a | 22.30 ± 5.25 b | −2.23 | * |
| Pylorus of stomach | - | 8.60 ± 1.41 c | - | - |
| Duodenum | 8.07 ± 0.34 b | 15.72 ± 1.40 b | 5.32 | ** |
| Jejunum | 7.47 ± 0.88 b | 18.89 ± 2.34 b | 4.57 | ** |
| Ileum | 9.43 ± 0.16 b | 16.35 ± 1.51 b | 4.54 | * |
| Colon | 19.54 ± 3.84 c | 18.56 ± 1.87 b | -0.23 | ns |
| Rectum | 2.81 ± 0.27 d | 6.49 ± 1.63 cd | 2.41 | * |
| Cecum | 7.25 ± 0.67 b | 1.66 ± 0.13 d | -8.21 | *** |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cai, X.; Bao, D.; Hua, R.; Cai, B.; Wang, L.; Dong, R.; Hua, L. A Comparative Study on the Distribution Pattern of Endocrine Cells in the Gastrointestinal Tract of Two Small Alpine Mammals, Plateau Zokor (Eospalax baileyi) and Plateau Pika (Ochotona curzoniae). Animals 2023, 13, 640. https://doi.org/10.3390/ani13040640
Cai X, Bao D, Hua R, Cai B, Wang L, Dong R, Hua L. A Comparative Study on the Distribution Pattern of Endocrine Cells in the Gastrointestinal Tract of Two Small Alpine Mammals, Plateau Zokor (Eospalax baileyi) and Plateau Pika (Ochotona curzoniae). Animals. 2023; 13(4):640. https://doi.org/10.3390/ani13040640
Chicago/Turabian StyleCai, Xincheng, Darhan Bao, Rui Hua, Bin Cai, Lei Wang, Rui Dong, and Limin Hua. 2023. "A Comparative Study on the Distribution Pattern of Endocrine Cells in the Gastrointestinal Tract of Two Small Alpine Mammals, Plateau Zokor (Eospalax baileyi) and Plateau Pika (Ochotona curzoniae)" Animals 13, no. 4: 640. https://doi.org/10.3390/ani13040640
APA StyleCai, X., Bao, D., Hua, R., Cai, B., Wang, L., Dong, R., & Hua, L. (2023). A Comparative Study on the Distribution Pattern of Endocrine Cells in the Gastrointestinal Tract of Two Small Alpine Mammals, Plateau Zokor (Eospalax baileyi) and Plateau Pika (Ochotona curzoniae). Animals, 13(4), 640. https://doi.org/10.3390/ani13040640