


Influence of Parturition on Rumen Bacteria and SCFAs in Holstein Cows Based on 16S rRNA Sequencing and Targeted Metabolomics

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Sample Collection
2.3. Targeted GC–MS/MS Metabolomics Analysis of Rumen SCFAs
2.4. 16S rRNA High-Throughput Sequencing of Rumen Bacteria Communities
3. Results
3.1. Changes in the Metabolic Profile and Concentrations of Rumen SCFAs
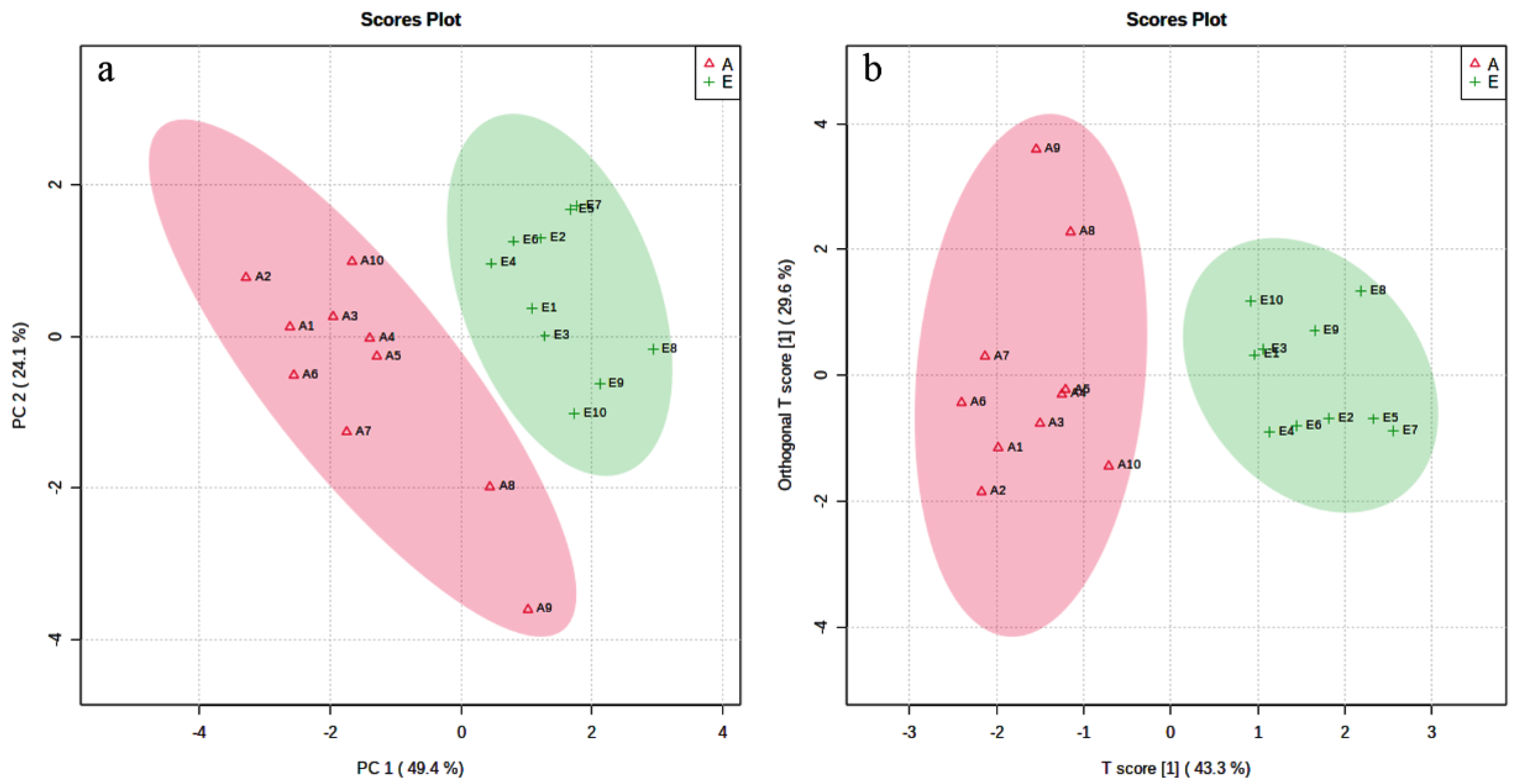
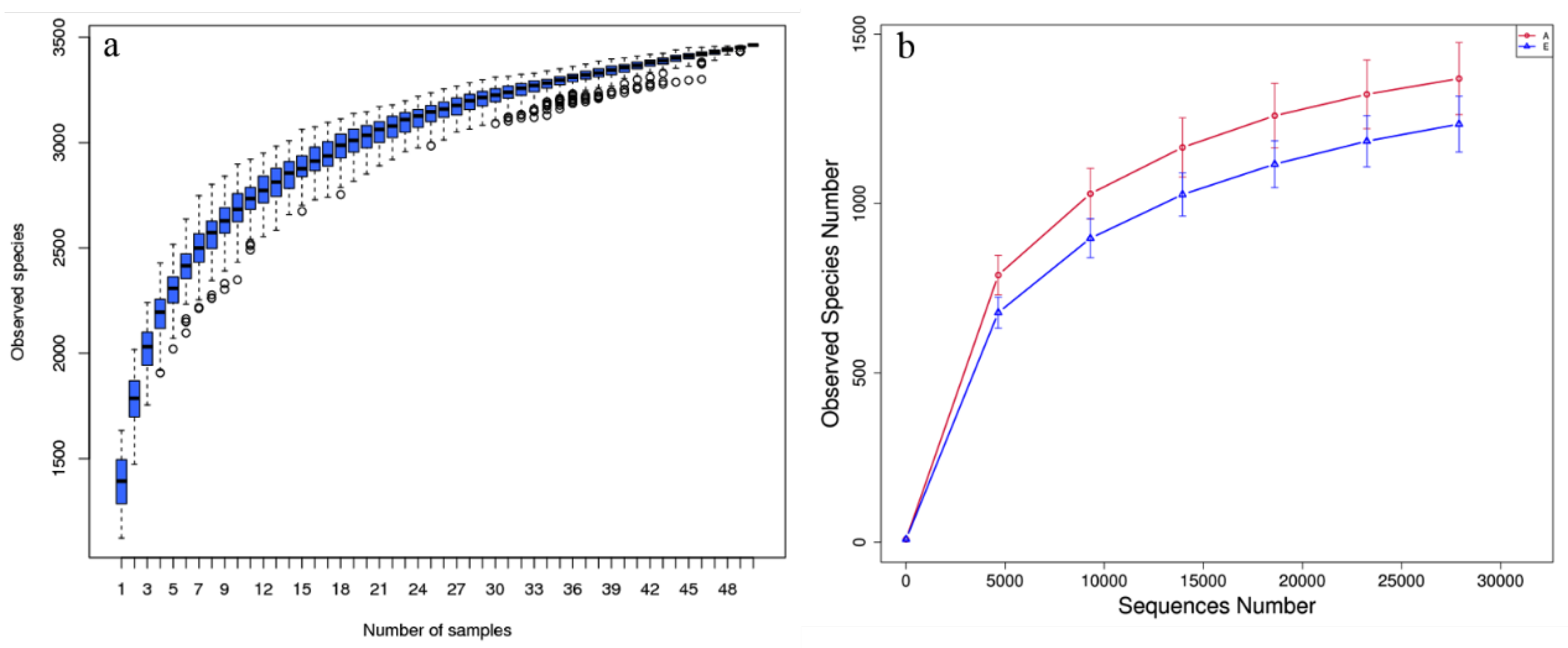
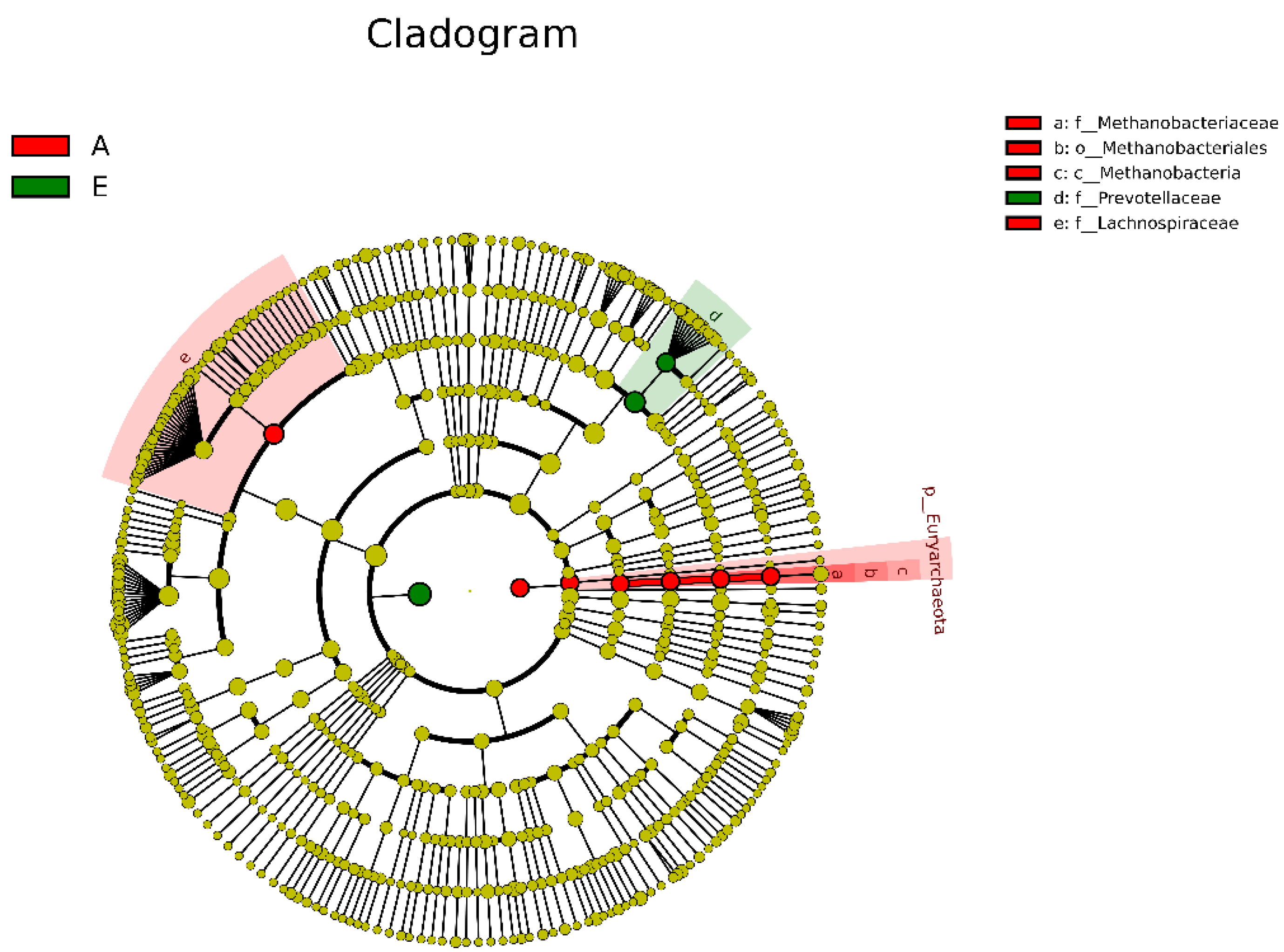
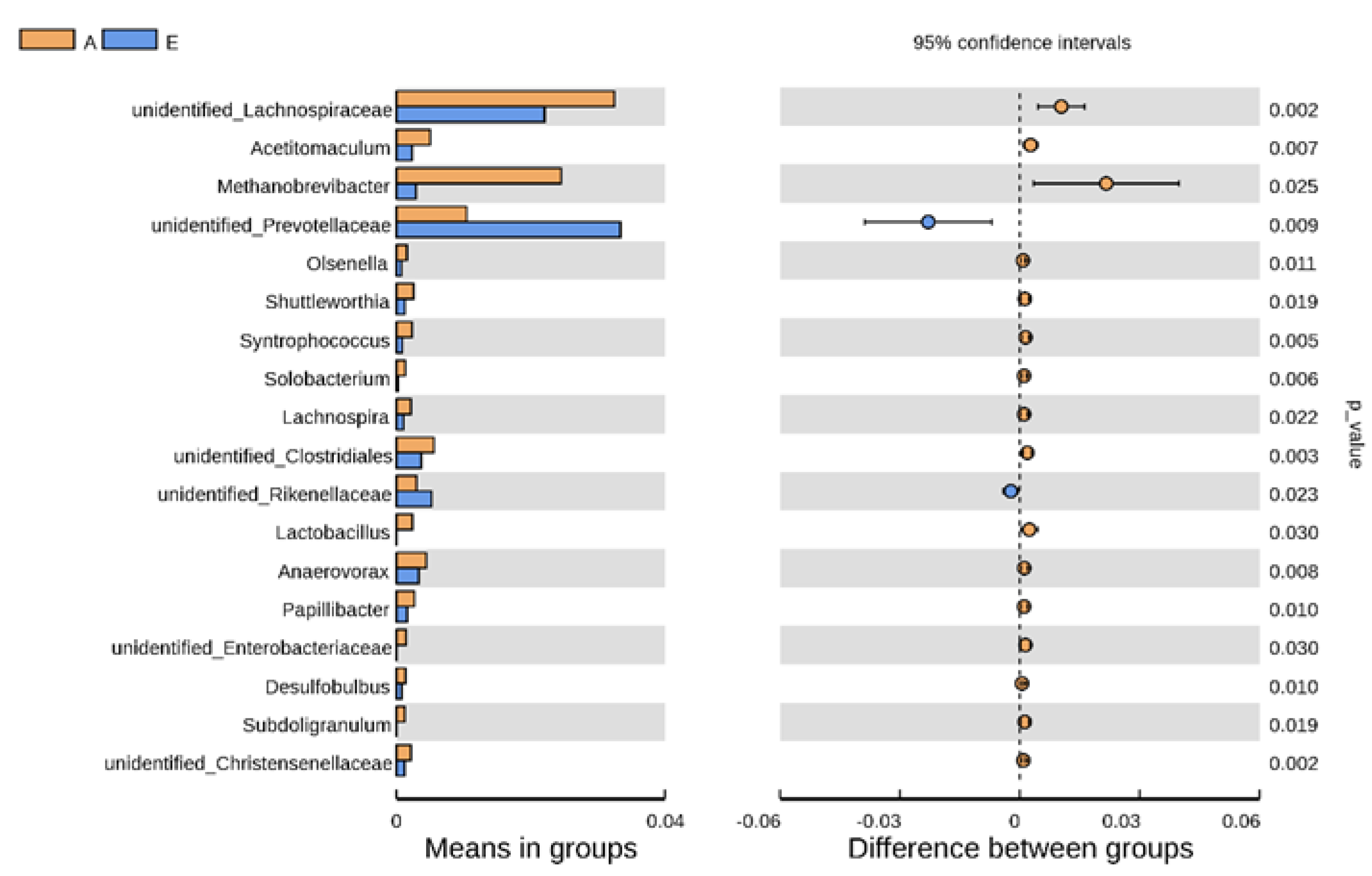
3.2. Alteration in Rumen Bacteria Communities
3.3. Correlation between Rumen Bacteria and SCFAs
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Erb, R.E.; Malven, P.V.; Monk, E.L.; Mollett, T.A.; Smith, K.L.; Schanbacher, F.L.; Willett, L.B. Hormone induced lactation in the cow. IV. Relationships between lactational performance and hormone concentrations in blood plasma. J. Dairy Sci. 1976, 59, 1420–1428. [Google Scholar] [CrossRef] [PubMed]
- Abuelo, A. Symposium review: Late-gestation maternal factors affecting the health and development of dairy calves. J. Dairy Sci. 2020, 103, 3882–3893. [Google Scholar] [CrossRef] [PubMed]
- Schirmann, K.; Chapinal, N.; Weary, D.M.; Vickers, L.; von Keyserlingk, M.A.G. Short communication: Rumination and feeding behavior before and after calving in dairy cows. J. Dairy Sci. 2013, 96, 7088–7092. [Google Scholar] [CrossRef] [PubMed]
- Beauchemin, K.A. Invited review: Current perspectives on eating and rumination activity in dairy cows. J. Dairy Sci. 2018, 101, 4762–4784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clark, C.E.; Lyons, N.A.; Millapan, L.; Talukder, S.; Cronin, G.M.; Kerrisk, K.L.; Garcia, S.C. Rumination and activity levels as predictors of calving for dairy cows. Anim. Int. J. Anim. Biosci. 2015, 9, 691–695. [Google Scholar] [CrossRef] [Green Version]
- Ku-Vera, J.C.; Jiménez-Ocampo, R.; Valencia-Salazar, S.S.; Montoya-Flores, M.D.; Molina-Botero, I.C.; Arango, J.; Gómez-Bravo, C.A.; Aguilar-Pérez, C.F.; Solorio-Sánchez, F.J. Role of Secondary Plant Metabolites on Enteric Methane Mitigation in Ruminants. Front. Vet. Sci. 2020, 7, 584. [Google Scholar] [CrossRef]
- Mason, P.M.; Stuckey, D.C. Biofilms, bubbles and boundary layers—A new approach to understanding cellulolysis in anaerobic and ruminant digestion. Water Res. 2016, 104, 93–100. [Google Scholar] [CrossRef]
- Johnson, J.S.; Spakowicz, D.J.; Hong, B.Y.; Petersen, L.M.; Demkowicz, P.; Chen, L.; Leopold, S.R.; Hanson, B.M.; Agresta, H.O.; Gerstein, M.; et al. Evaluation of 16S rRNA gene sequencing for species and strain-level microbiome analysis. Nat. Commun. 2019, 10, 5029. [Google Scholar] [CrossRef] [Green Version]
- Thoetkiattikul, H.; Mhuantong, W.; Laothanachareon, T.; Tangphatsornruang, S.; Pattarajinda, V.; Eurwilaichitr, L.; Champreda, V. Comparative analysis of microbial profiles in cow rumen fed with different dietary fiber by tagged 16S rRNA gene pyrosequencing. Curr. Microbiol. 2013, 67, 130–137. [Google Scholar] [CrossRef]
- Guo, W.; Li, Y.; Wang, L.; Wang, J.; Xu, Q.; Yan, T.; Xue, B. Evaluation of composition and individual variability of rumen microbiota in yaks by 16S rRNA high-throughput sequencing technology. Anaerobe 2015, 34, 74–79. [Google Scholar] [CrossRef]
- Dervishi, E.; Zhang, G.; Dunn, S.M.; Mandal, R.; Wishart, D.S.; Ametaj, B.N. GC-MS Metabolomics Identifies Metabolite Alterations That Precede Subclinical Mastitis in the Blood of Transition Dairy Cows. J. Proteome Res. 2017, 16, 433–446. [Google Scholar] [CrossRef] [PubMed]
- Xue, F.; Pan, X.; Jiang, L.; Guo, Y.; Xiong, B. GC-MS analysis of the ruminal metabolome response to thiamine supplementation during high grain feeding in dairy cows. Metab. Off. J. Metab. Soc. 2018, 14, 67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, Q.; Qiao, Q.; Gao, Y.; Hou, J.; Hu, M.; Du, Y.; Zhao, K.; Li, X. Gut Microbiota and Their Role in Health and Metabolic Disease of Dairy Cow. Front. Nutr. 2021, 8, 701511. [Google Scholar] [CrossRef] [PubMed]
- Bianchi, F.; Dall’Asta, M.; Del Rio, D.; Mangia, A.; Musci, M.; Scazzina, F. Development of a headspace solid-phase microextraction gas chromatography–mass spectrometric method for the determination of short-chain fatty acids from intestinal fermentation. Food Chem. 2011, 129, 200–205. [Google Scholar] [CrossRef]
- Göker, M.; Held, B.; Lucas, S.; Nolan, M.; Yasawong, M.; Del Rio, T.G.; Tice, H.; Cheng, J.F.; Bruce, D.; Detter, J.C.; et al. Complete genome sequence of Olsenella uli type strain (VPI D76D-27C). Stand. Genom. Sci. 2010, 3, 76–84. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Brejnrod, A.D.; Ernst, M.; Rykær, M.; Herschend, J.; Olsen, N.M.C.; Dorrestein, P.C.; Rensing, C.; Sørensen, S.J. Heavy metal exposure causes changes in the metabolic health-associated gut microbiome and metabolites. Environ. Int. 2019, 126, 454–467. [Google Scholar] [CrossRef]
- Liu, C.; Wu, H.; Liu, S.; Chai, S.; Meng, Q.; Zhou, Z. Dynamic Alterations in Yak Rumen Bacteria Community and Metabolome Characteristics in Response to Feed Type. Front. Microbiol. 2019, 10, 1116. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Magoč, T.; Salzberg, S.L. FLASH: Fast length adjustment of short reads to improve genome assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Bokulich, N.A.; Subramanian, S.; Faith, J.J.; Gevers, D.; Gordon, J.I.; Knight, R.; Mills, D.A.; Caporaso, J.G. Quality-filtering vastly improves diversity estimates from Illumina amplicon sequencing. Nat. Methods 2013, 10, 57–59. [Google Scholar] [CrossRef] [PubMed]
- Caporaso, J.G.; Kuczynski, J.; Stombaugh, J.; Bittinger, K.; Bushman, F.D.; Costello, E.K.; Fierer, N.; Peña, A.G.; Goodrich, J.K.; Gordon, J.I.; et al. QIIME allows analysis of high-throughput community sequencing data. Nat. Methods 2010, 7, 335–336. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haas, B.J.; Gevers, D.; Earl, A.M.; Feldgarden, M.; Ward, D.V.; Giannoukos, G.; Ciulla, D.; Tabbaa, D.; Highlander, S.K.; Sodergren, E.; et al. Chimeric 16S rRNA sequence formation and detection in Sanger and 454-pyrosequenced PCR amplicons. Genome Res. 2011, 21, 494–504. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Mulder, T.; Goossens, K.; Peiren, N.; Vandaele, L.; Haegeman, A.; De Tender, C.; Ruttink, T.; de Wiele, T.V.; De Campeneere, S. Exploring the methanogen and bacterial communities of rumen environments: Solid adherent, fluid and epimural. FEMS Microbiol. Ecol. 2017, 93, fiw251. [Google Scholar] [CrossRef] [Green Version]
- McLoughlin, S.; Spillane, C.; Claffey, N.; Smith, P.E.; O’Rourke, T.; Diskin, M.G.; Waters, S.M. Rumen Microbiome Composition Is Altered in Sheep Divergent in Feed Efficiency. Front. Microbiol. 2020, 11, 1981. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Mu, C.; Xu, Y.; Shen, J.; Zhu, W. Changes in the Solid-, Liquid-, and Epithelium-Associated Bacterial Communities in the Rumen of Hu Lambs in Response to Dietary Urea Supplementation. Front. Microbiol. 2020, 11, 244. [Google Scholar] [CrossRef] [Green Version]
- McGovern, E.; Kenny, D.A.; McCabe, M.S.; Fitzsimons, C.; McGee, M.; Kelly, A.K.; Waters, S.M. 16S rRNA Sequencing Reveals Relationship Between Potent Cellulolytic Genera and Feed Efficiency in the Rumen of Bulls. Front. Microbiol. 2018, 9, 1842. [Google Scholar] [CrossRef]
- Schären, M.; Kiri, K.; Riede, S.; Gardener, M.; Meyer, U.; Hummel, J.; Urich, T.; Breves, G.; Dänicke, S. Alterations in the Rumen Liquid-, Particle- and Epithelium-Associated Microbiota of Dairy Cows during the Transition from a Silage- and Concentrate-Based Ration to Pasture in Spring. Front. Microbiol. 2017, 8, 744. [Google Scholar] [CrossRef]
- Kim, S.H.; Mamuad, L.L.; Islam, M.; Lee, S.S. Reductive acetogens isolated from ruminants and their effect on in vitro methane mitigation and milk performance in Holstein cows. J. Anim. Sci. Technol. 2020, 62, 239. [Google Scholar] [CrossRef] [Green Version]
- Lemosquet, S.; Raggio, G.; Lobley, G.E.; Rulquin, H.; Guinard-Flament, J.; Lapierre, H. Whole-body glucose metabolism and mammary energetic nutrient metabolism in lactating dairy cows receiving digestive infusions of casein and propionic acid. J. Dairy Sci. 2009, 92, 6068–6082. [Google Scholar] [CrossRef] [Green Version]
- Lv, F.; Wang, X.; Pang, X.; Liu, G. Effects of supplementary feeding on the rumen morphology and bacterial diversity in lambs. PeerJ 2020, 8, e9353. [Google Scholar] [CrossRef] [PubMed]
- Gonçalves, P.; Araújo, J.R.; Di Santo, J.P. A Cross-Talk Between Microbiota-Derived Short-Chain Fatty Acids and the Host Mucosal Immune System Regulates Intestinal Homeostasis and Inflammatory Bowel Disease. Inflamm. Bowel Dis. 2018, 24, 558–572. [Google Scholar] [CrossRef] [PubMed]
- Block, S.S.; Butler, W.R.; Ehrhardt, R.A.; Bell, A.W.; Van Amburgh, M.E.; Boisclair, Y.R. Decreased concentration of plasma leptin in periparturient dairy cows is caused by negative energy balance. J. Endocrinol. 2001, 171, 339–348. [Google Scholar] [CrossRef] [PubMed]
- Myers, M.N.; Zachut, M.; Tam, J.; Contreras, G.A. A proposed modulatory role of the endocannabinoid system on adipose tissue metabolism and appetite in periparturient dairy cows. J. Anim. Sci. Biotechnol. 2021, 12, 21. [Google Scholar] [CrossRef] [PubMed]
- Bines, J.A.; Hart, I.C. Metabolic limits to milk production, especially roles of growth hormone and insulin. J. Dairy Sci. 1982, 65, 1375–1389. [Google Scholar] [CrossRef] [PubMed]
- Schleicher, L.; Trautmann, A.; Stegmann, D.P.; Fritz, G.; Gätgens, J.; Bott, M.; Hein, S.; Simon, J.; Seifert, J.; Steuber, J. A Sodium-Translocating Module Linking Succinate Production to Formation of Membrane Potential in Prevotella bryantii. Appl. Environ. Microbiol. 2021, 87, e0121121. [Google Scholar] [CrossRef]
- Reynolds, C.K.; Huntington, G.B.; Tyrrell, H.F.; Reynolds, P.J. Net metabolism of volatile fatty acids, D-beta-hydroxybutyrate, nonesterifield fatty acids, and blood gasses by portal-drained viscera and liver of lactating Holstein cows. J. Dairy Sci. 1988, 71, 2395–2405. [Google Scholar] [CrossRef] [PubMed]
- Esposito, G.; Irons, P.C.; Webb, E.C.; Chapwanya, A. Interactions between negative energy balance, metabolic diseases, uterine health and immune response in transition dairy cows. Anim. Reprod. Sci. 2014, 144, 60–71. [Google Scholar] [CrossRef] [Green Version]
- Lan, W.; Yang, C. Ruminal methane production: Associated microorganisms and the potential of applying hydrogen-utilizing bacteria for mitigation. Sci. Total Environ. 2019, 654, 1270–1283. [Google Scholar] [CrossRef]
- Tian, Y.; Zhang, H.; Zheng, L.; Li, S.; Hao, H.; Huang, H. Effect of Zn Addition on the Cd-Containing Anaerobic Fermentation Process: Biodegradation and Microbial Communities. Int. J. Environ. Res. Public Health 2019, 16, 2998. [Google Scholar] [CrossRef] [Green Version]
- Wallace, R.J.; Rooke, J.A.; McKain, N.; Duthie, C.A.; Hyslop, J.J.; Ross, D.W.; Waterhouse, A.; Watson, M.; Roehe, R. The rumen microbial metagenome associated with high methane production in cattle. BMC Genom. 2015, 16, 839. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kong, F.; Gao, Y.; Tang, M.; Fu, T.; Diao, Q.; Bi, Y.; Tu, Y. Effects of dietary rumen-protected Lys levels on rumen fermentation and bacterial community composition in Holstein heifers. Appl. Microbiol. Biotechnol. 2020, 104, 6623–6634. [Google Scholar] [CrossRef]
- Hua, C.; Tian, J.; Tian, P.; Cong, R.; Luo, Y.; Geng, Y.; Tao, S.; Ni, Y.; Zhao, R. Feeding a High Concentration Diet Induces Unhealthy Alterations in the Composition and Metabolism of Ruminal Microbiota and Host Response in a Goat Model. Front. Microbiol. 2017, 8, 138. [Google Scholar] [CrossRef] [Green Version]
- Markowiak-Kopeć, P.; Śliżewska, K. The Effect of Probiotics on the Production of Short-Chain Fatty Acids by Human Intestinal Microbiome. Nutrients 2020, 12, 1107. [Google Scholar] [CrossRef] [Green Version]
- Patra, A.K.; Yu, Z. Essential oils affect populations of some rumen bacteria in vitro as revealed by microarray (RumenBactArray) analysis. Front. Microbiol. 2015, 6, 297. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dusková, D.; Marounek, M. Fermentation of pectin and glucose, and activity of pectin-degrading enzymes in the rumen bacterium Lachnospira multiparus. Lett. Appl. Microbiol. 2001, 33, 159–163. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Component | Retention Time (min) | Regression Equation | Concentrations (Mean ± SD) (μg/mL) | Statistical Parameters (Before Parturition vs. After Parturition) | |||
|---|---|---|---|---|---|---|---|
| Before Parturition (n = 10) | After Parturition (n = 10) | p Value | FC | VIP | |||
| Acetic acid | 3.36 | y = 0.64x + 0.03 | 9575 ± 117.54 | 6340 ± 294.36 | 0.01 | 1.51 ↓ | 1.21 |
| Propionic acid | 3.97 | y = 0.46x + 0.001 | 6109 ± 72.89 | 3236.8 ± 165.61 | 0.0004 | 1.89 ↓ | 1.36 |
| Isobutyric acid | 4.14 | y = 1.00x − 0.001 | 339.4 ± 8.47 | 366.4 ± 23.23 | 0.75 | 0.93 | 1.18 |
| Butyric acid | 4.55 | y = 5.57x + 0.20 | 5218 ± 75.27 | 3281.7 ± 149.00 | 0.004 | 1.59 ↓ | 1.01 |
| Isovaleric acid | 4.8 | y = 6.49x − 0.001 | 345.5 ± 7.17 | 349.6 ± 20.46 | 0.96 | 0.99 | 1.36 |
| Valeric acid | 5.22 | y = 7.12x + 0.13 | 716.7 ± 12.10 | 416.8 ± 21.69 | 0.003 | 1.72 | 0.63 |
| Caproic acid | 5.79 | y = 3.39x + 0.23 | 310.8 ± 8.51 | 141.7 ± 8.78 | 0.0006 | 2.19 ↓ | 1.15 |
| pH | 6.62 ± 0.0624 | 6.82 ± 0.03 | 0.0081 | 0.97 ↑ | |||
| Items | Before Parturition, n = 10 | After Parturition, n = 10 | p Value |
|---|---|---|---|
| Shannon | 7.25 ± 0.48 | 7.86 ± 0.32 | 0.0042 |
| Simpson | 0.96 ± 0.02 | 0.97 ± 0.008 | 0.1762 |
| Chao1 | 1495.27 ± 115.85 | 1673.59 ± 157.62 | 0.0205 |
| ACE | 15,224.87 ± 84.09 | 1701.15 ± 164.32 | 0.0177 |
| Taxonomy | Percentage of the Relative Abundance | SEM | p Value | Trend | Difference Multiple | ||
|---|---|---|---|---|---|---|---|
| Before Parturition | After Parturition | □AVE | |||||
| Unidentified_Lachnospiraceae | 2.01 | 3.99 | 2.99 | 0.99 | 0.002 | up | 2.00 |
| Acetitomaculum | 0.23 | 0.50 | 0.37 | 0.14 | 0.007 | up | 2.16 |
| Methanobrevibacter | 0.29 | 2.46 | 1.37 | 1.08 | 0.025 | up | 8.43 |
| Unidentified_Prevotellaceae | 3.34 | 1.05 | 2.19 | 1.14 | 0.009 | down | 3.18 |
| Olsenella | 0.08 | 0.16 | 0.12 | 0.04 | 0.011 | up | 2.08 |
| Syntrophococcus | 0.09 | 0.23 | 0.16 | 0.07 | 0.005 | up | 2.63 |
| Lachnospira | 0.11 | 0.22 | 0.16 | 0.06 | 0.022 | up | 2.03 |
| Lactobacillus | 0.0004 | 0.24 | 0.12 | 0.11 | 0.030 | up | 600.25 |
| SCFAs | Bacterial Genera | Correlation Coefficient (r) | p-Value | AdjPvalue |
|---|---|---|---|---|
| Acetic acid | Methanobrevibacter | −0.601 | 0.005 ** | 0.013 |
| Acetitomaculum | −0.469 | 0.037 * | 0.037 | |
| Syntrophococcus | −0.472 | 0.036 * | 0.036 | |
| Lactobacillus | −0.763 | 0.000 ** | 0.000 | |
| Unidentified_Enterobacteriaceae | −0.680 | 0.001 ** | 0.001 | |
| Propionic acid | Unidentified_Prevotellaceae | 0.477 | 0.035 * | 0.144 |
| Unidentified_Lachnospiraceae | −0.567 | 0.010 * | 0.052 | |
| Lachnospira | −0.447 | 0.048 * | 0.241 | |
| Lactobacillus | −0.807 | 0.000 ** | 0.000 | |
| Olsenella | −0.629 | 0.003 ** | 0.008 | |
| Desulfobulbus | −0.515 | 0.020 * | 0.020 | |
| Unidentified_Enterobacteriaceae | −0.730 | 0.000 ** | 0.001 | |
| Butyric acid | Acetitomaculum | −0.476 | 0.034 * | 0.037 |
| Syntrophococcus | −0.489 | 0.029 * | 0.036 | |
| Lactobacillus | −0.746 | 0.000 ** | 0.000 | |
| Desulfobulbus | −0.642 | 0.002 ** | 0.006 | |
| Solobacterium | −0.716 | 0.000 ** | 0.000 | |
| Unidentified_Enterobacteriaceae | −0.673 | 0.001 ** | 0.001 | |
| Subdoligranulum | −0.786 | 0.000 ** | 0.000 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Guo, Y.; Wang, F.; Mao, Y.; Kong, W.; Wang, J.; Zhang, G. Influence of Parturition on Rumen Bacteria and SCFAs in Holstein Cows Based on 16S rRNA Sequencing and Targeted Metabolomics. Animals 2023, 13, 782. https://doi.org/10.3390/ani13050782
Guo Y, Wang F, Mao Y, Kong W, Wang J, Zhang G. Influence of Parturition on Rumen Bacteria and SCFAs in Holstein Cows Based on 16S rRNA Sequencing and Targeted Metabolomics. Animals. 2023; 13(5):782. https://doi.org/10.3390/ani13050782
Chicago/Turabian StyleGuo, Yansheng, Feifei Wang, Yongxia Mao, Weiyi Kong, Jiandong Wang, and Guijie Zhang. 2023. "Influence of Parturition on Rumen Bacteria and SCFAs in Holstein Cows Based on 16S rRNA Sequencing and Targeted Metabolomics" Animals 13, no. 5: 782. https://doi.org/10.3390/ani13050782
APA StyleGuo, Y., Wang, F., Mao, Y., Kong, W., Wang, J., & Zhang, G. (2023). Influence of Parturition on Rumen Bacteria and SCFAs in Holstein Cows Based on 16S rRNA Sequencing and Targeted Metabolomics. Animals, 13(5), 782. https://doi.org/10.3390/ani13050782