
Transcriptome Analysis Reveals the Immunoregulation of Replacing Fishmeal with Cottonseed Protein Concentrates on Litopenaeus vannamei

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Method
2.1. Experimental Diets
2.2. Feeding Trial and Experimental Conditions
2.3. Sample Collection
2.4. Evaluation of Non-Specific Immune Indices
2.5. Gene Expression Analysis
2.6. Transcriptome Sequencing and Analysis
2.6.1. RNA Extraction and Transcriptome Sequencing
2.6.2. Data Analysis
2.6.3. Differential Expression Analysis and Functional Annotation
2.7. Validation of DEGs by qPCR
2.8. Statistical Analysis
3. Results
3.1. Non-Specific Immune Indices in the Serum
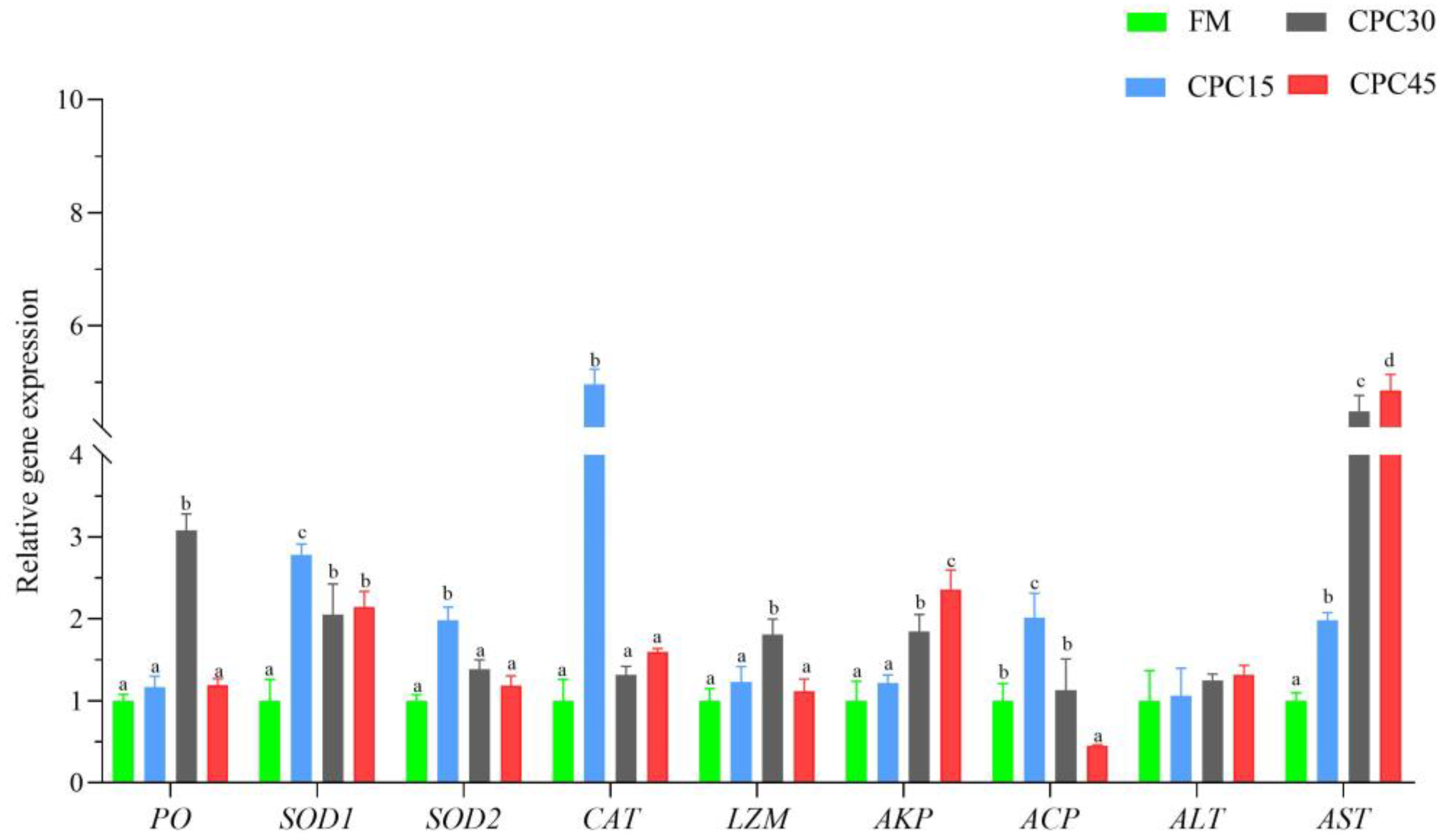
3.2. Expression of Non-Specific Immune Genes in Hemocytes
3.3. Assembly and Sequence Alignment Analysis
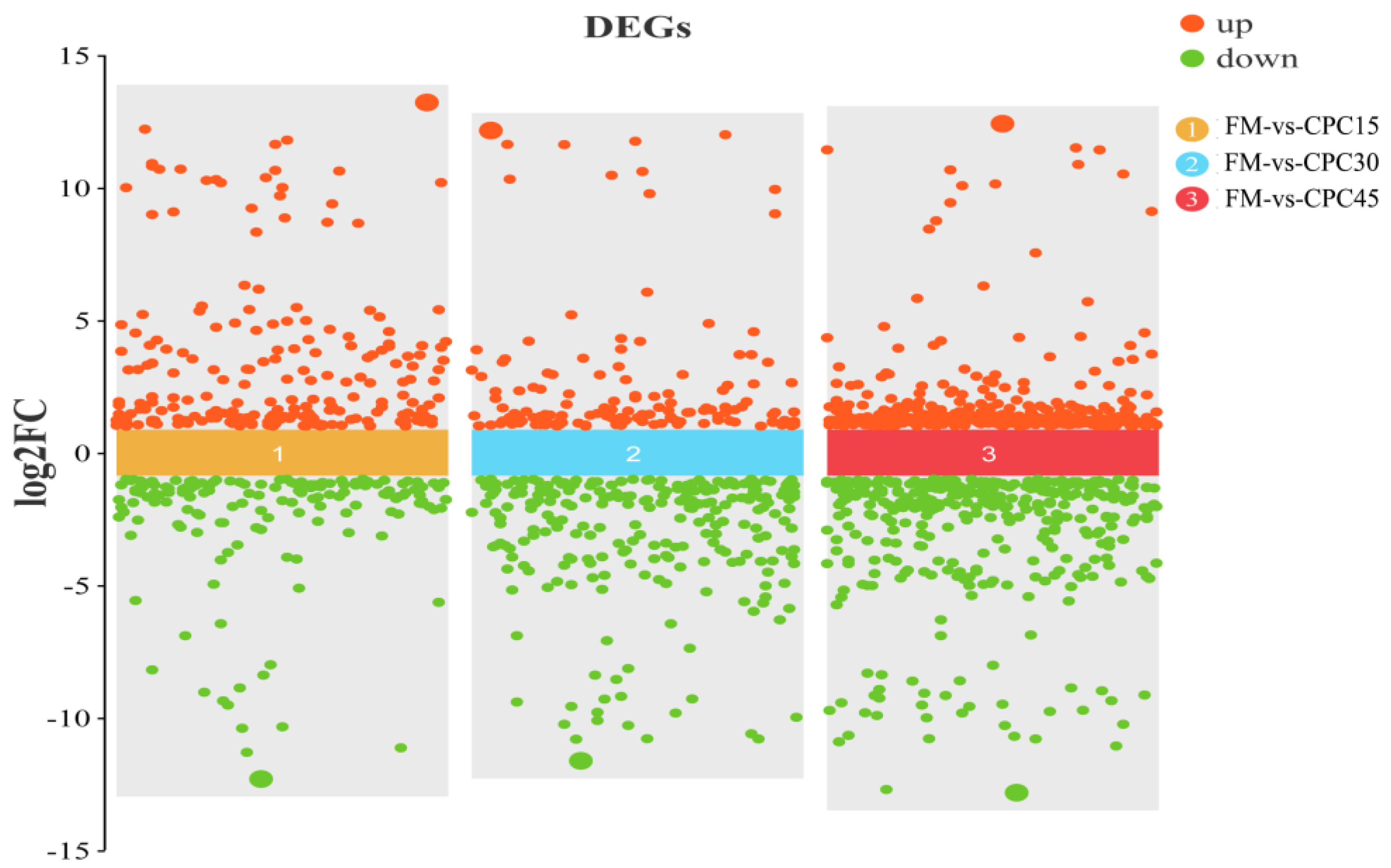
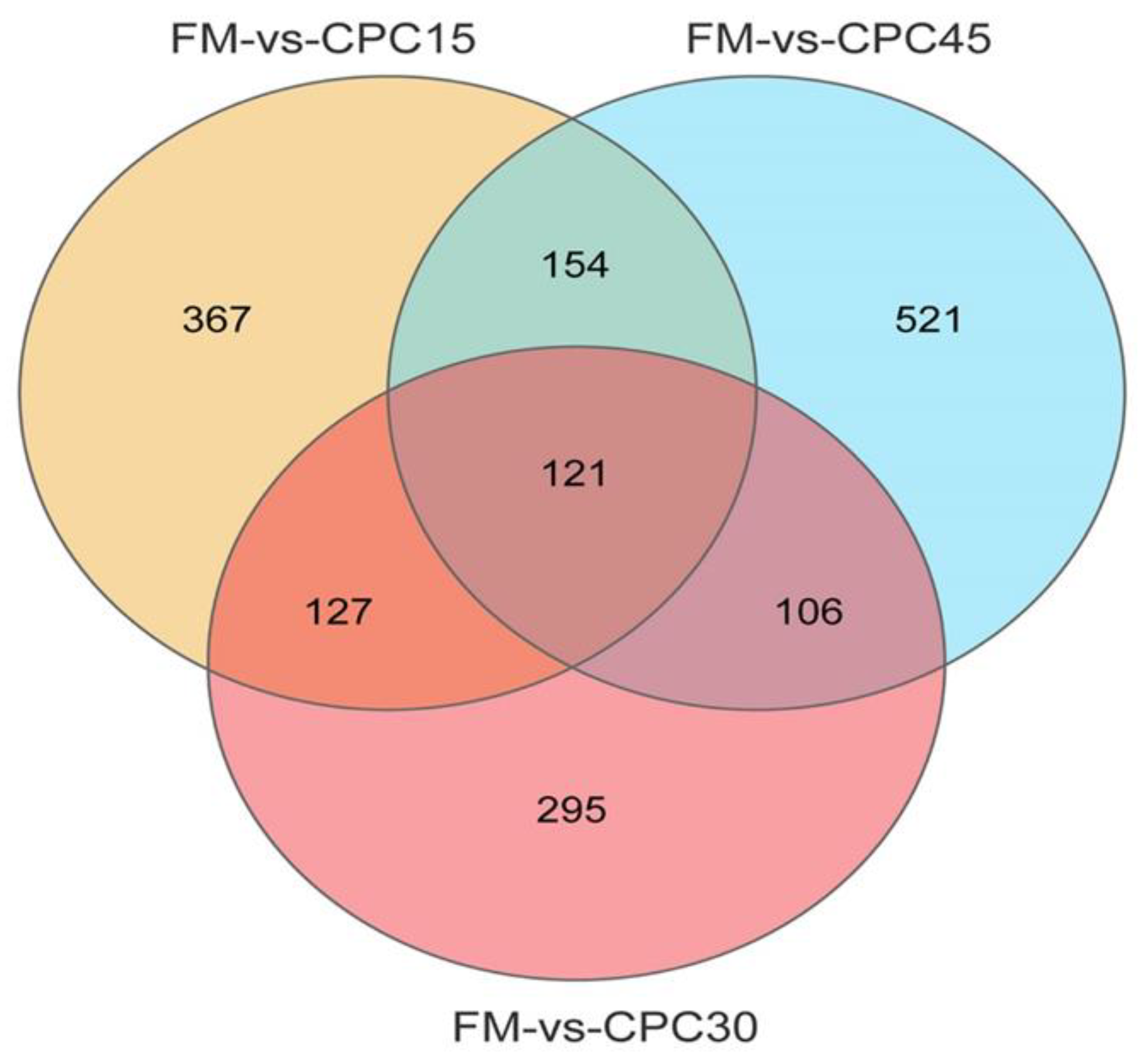
3.4. Identification of DEGs
3.5. GO Enrichment Analysis of the DEGs
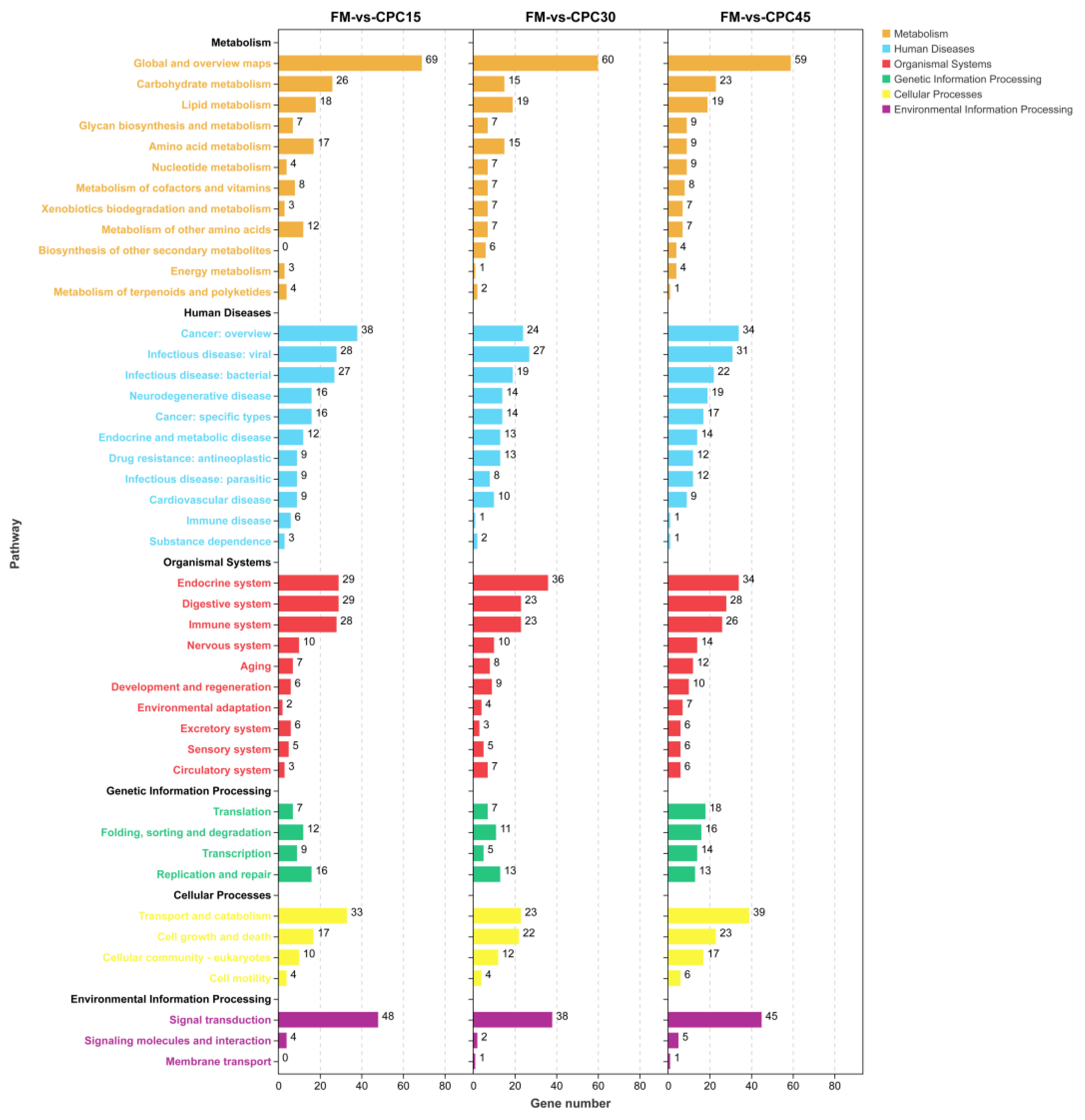
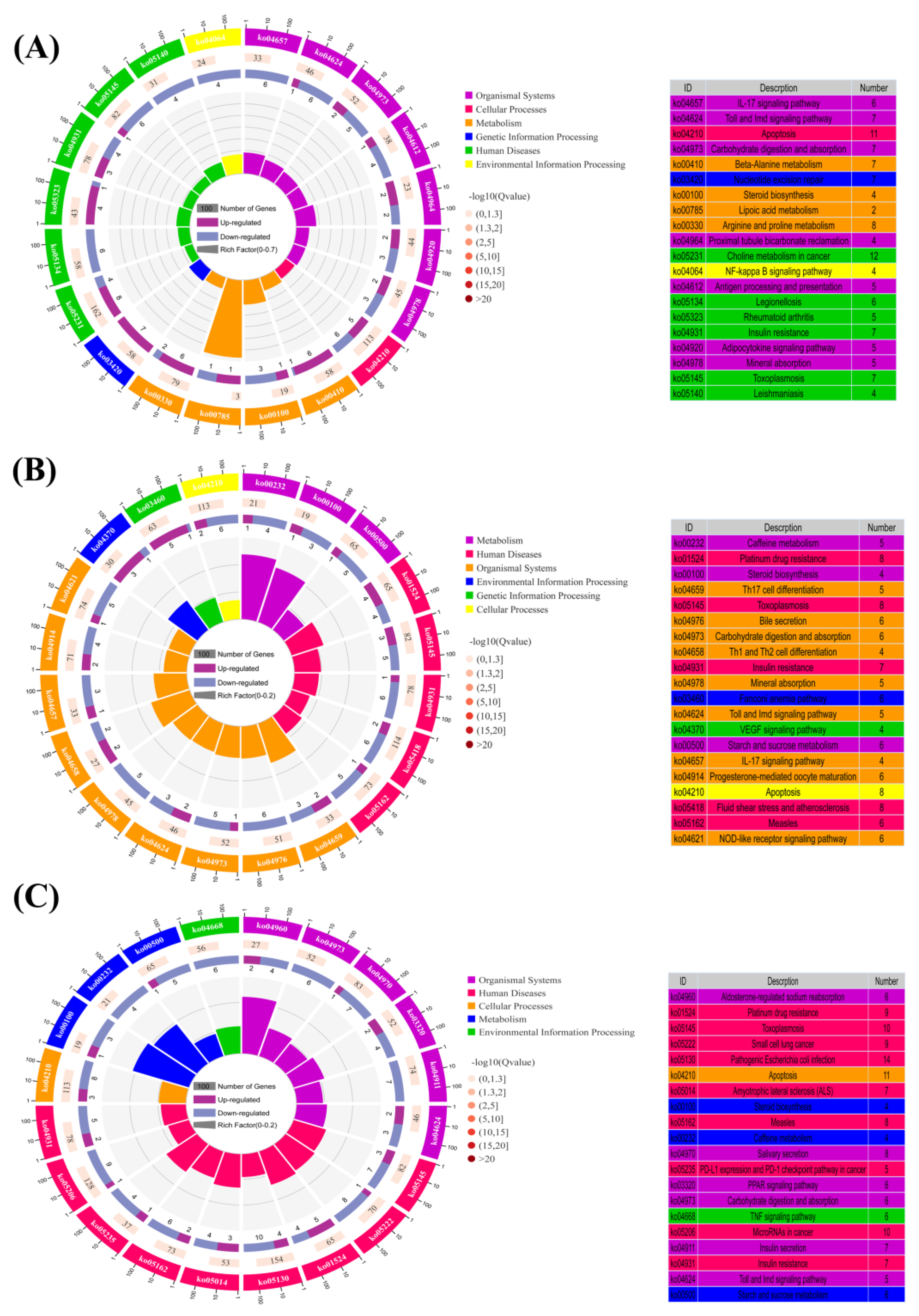
3.6. KEGG Enrichment Analysis of the DEGs
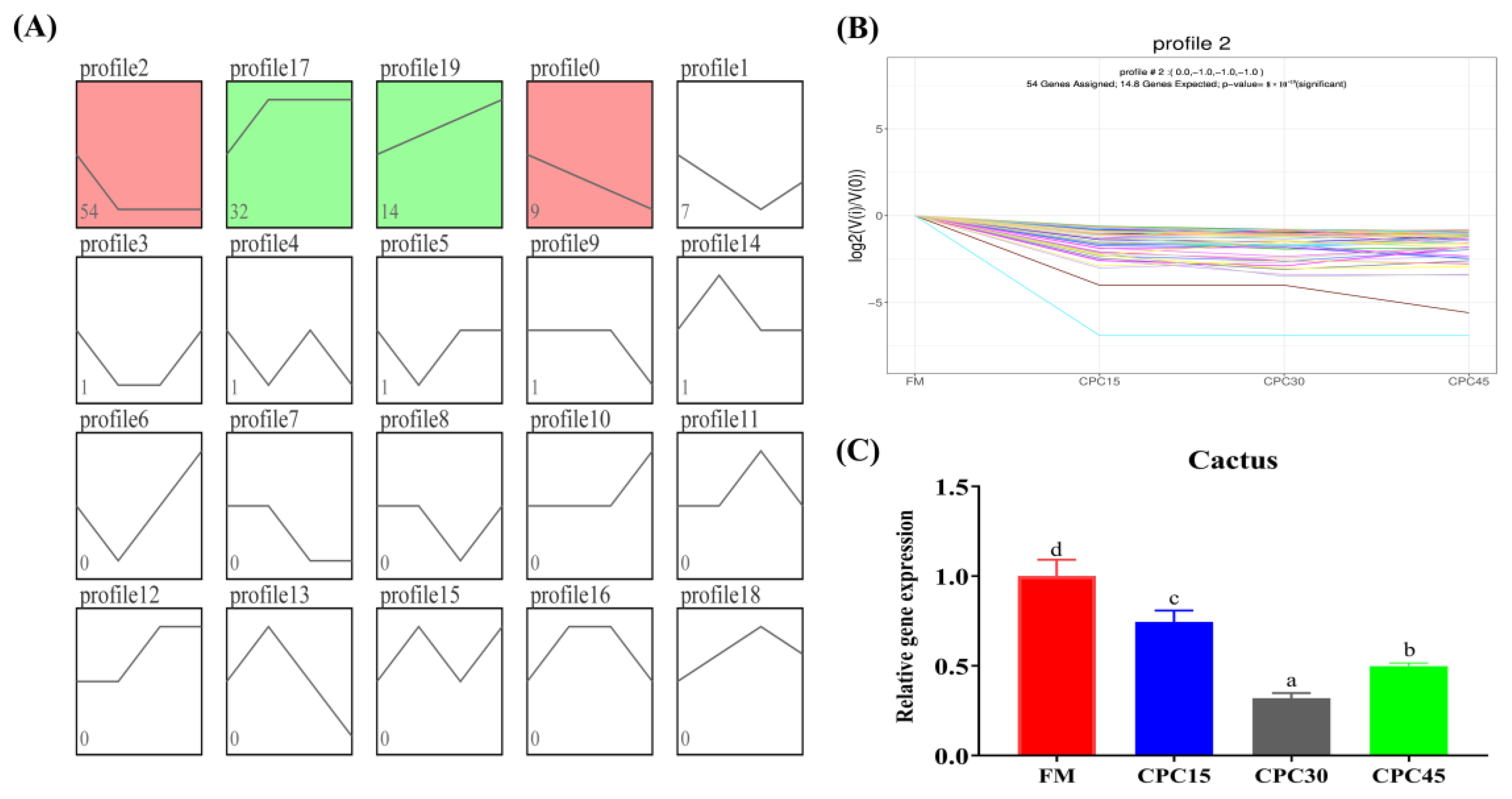
3.7. Series Test of Cluster
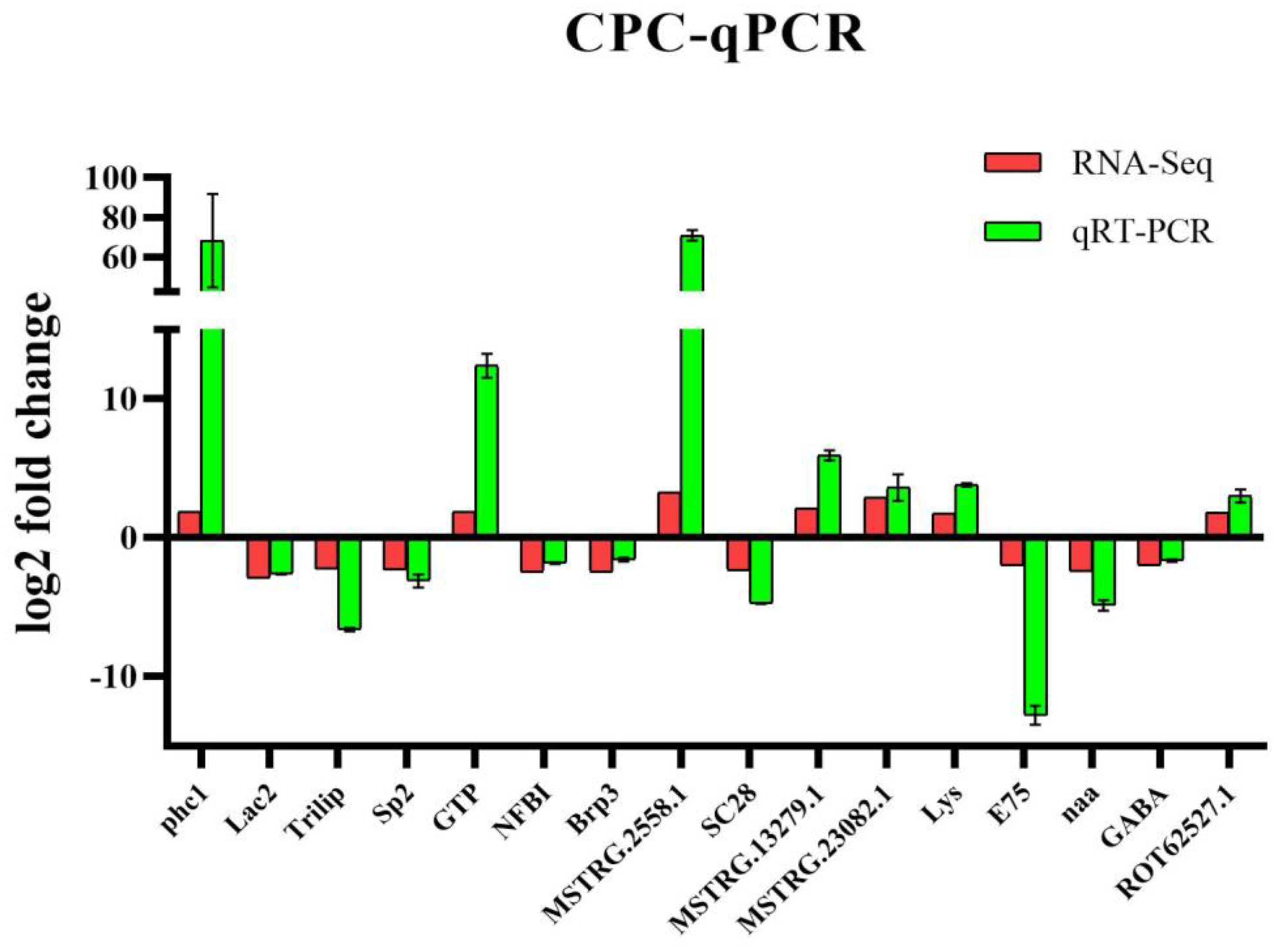
3.8. Validation of qPCR
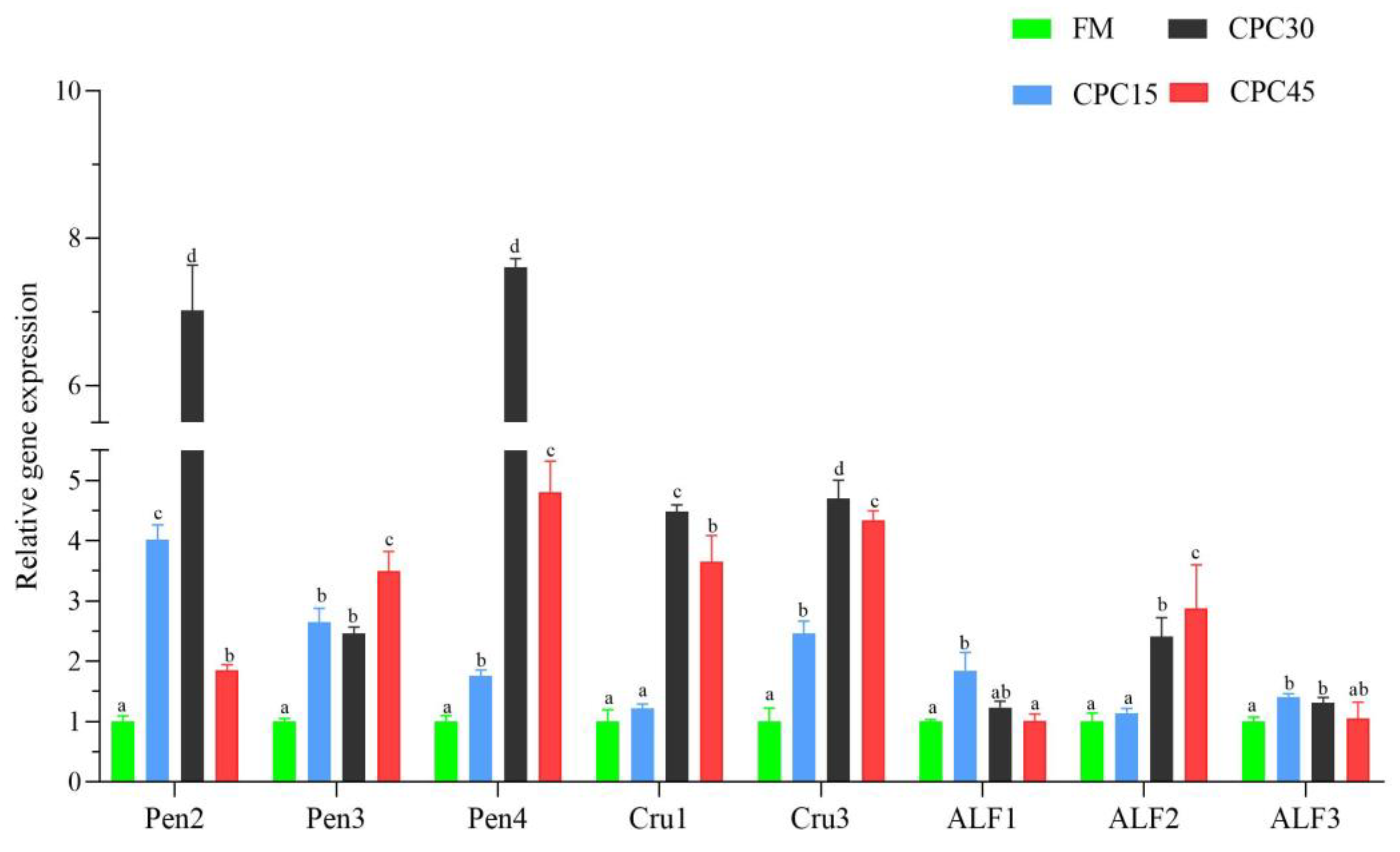
3.9. Expression of AMP Genes in Hemocytes
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Sookying, D.; Davis, D.A.; Soller Dias da Silva, F. A Review of the Development and Application of Soybean-Based Diets for Pacific White Shrimp Litopenaeus Vannamei. Aquac. Nutr. 2013, 19, 441–448. [Google Scholar] [CrossRef]
- Lu, X.; Luan, S.; Luo, K.; Meng, X.; Li, W.; Sui, J.; Cao, B.; Kong, J. Genetic Analysis of the Pacific White Shrimp (Litopenaeus Vannamei): Heterosis and Heritability for Harvest Body Weight. Aquac. Res. 2016, 47, 3365–3375. [Google Scholar] [CrossRef]
- Wang, A.; Yang, Q.; Tan, B.; Weiwei, X.; Jing, J.; Xiaohui, D.; Shuyan, C.; Hongyu, L.; Shuang, Z. Effects of Soy Protein Hydrolysate on Growth Performance, Serum Biochemical Parameters, Hapten and Disease Resistance of Litopenaeus Vannamei Larvae. J. Guangdong Ocean Univ. 2018, 1, 14–21. [Google Scholar]
- He, Y.; Chi, S.; Zhang, H.; Tan, B.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S. Effect of Yeast Culture on the Structure of Intestinal Microflora in Litopenaeus Vannamei. J. Guangdong Ocean Univ. 2017, 37, 21–27. [Google Scholar]
- Ye, G.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, H.; Tan, B.; Zhang, S. Dietary Replacement of FishMeal with Peanut Meal in Juvenile Hybrid Grouper (Epinephelus Fuscoguttatus ♀ × Epinephelus Lanceolatus ♂): Growth Performance, Immune Response and Intestinal Microbiota. Aquac. Rep. 2020, 17, 100327. [Google Scholar] [CrossRef]
- Zheng, Y.; Hou, C.; Yan, Z.; Chen, J.; Wang, H.; Tan, B.; Zhang, S. Effects of Dietary Zymosan-A on the Growth Performance and Intestinal Morphology, Digestive Capacity, and Microbial Community in Litopenaeus Vannamei. Front. Mar. Sci. 2022, 9, 439. [Google Scholar] [CrossRef]
- Ray, G.; Liang, D.; Yang, Q.; Tan, B.; Dong, X.; Chi, S.; Liu, H.; Zhang, S.; Li, R. Effects of Replacing Fishmeal with Dietary Soybean Protein Concentrate (SPC) on Growth, Serum Biochemical Indices, and Antioxidative Functions for Juvenile Shrimp Litopenaeus Vannamei. Aquaculture 2020, 516, 734630. [Google Scholar] [CrossRef]
- Yousif, R.; Khan, M.; Zehra, S. Effect of Replacing Fishmeal with Soybean Meal on Growth, Feed Conversion and Carcass Composition of Fingerling Oreochromis Niloticus (Nile Tilapia). Aust. J. Agric. Vet. Sci. 2021, 22, 85–92. [Google Scholar]
- Liang, X.; Han, J.; Xue, M.; Yu, H.; Huang, H.; Wu, X.; Zheng, Y.; Qin, Y.; Liang, X. Growth and Feed Intake Regulation Responses to Anorexia, Adaptation and Fasting in Japanese Seabss, Lateolabrax Japonicas when Fishmeal Is Totally Replaced by Plant Protein. Aquaculture 2019, 498, 528–538. [Google Scholar] [CrossRef]
- Shen, J.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Effects of Replacement of Fishmeal with Cottonseed Protein Concentrate on the Growth, Intestinal Microflora, Haematological and Antioxidant Indices of Juvenile Golden Pompano (Trachinotus Ovatus). Aquac. Nutr. 2020, 26, 1119–1130. [Google Scholar] [CrossRef]
- Song, Z.; Li, H.; Wang, J.; Li, P.; Sun, Y.; Zhang, L. Effects of Fishmeal Replacement with Soy Protein Hydrolysates on Growth Performance, Blood Biochemistry, Gastrointestinal Digestion and Muscle Composition of Juvenile Starry Flounder (Platichthys Stellatus). Aquaculture 2014, 426, 96–104. [Google Scholar] [CrossRef]
- Wang, J.; Clark, G.; Ju, M.; Castillo, S.; Gatlin, D.M. Effects of Replacing Menhaden Fishmeal with Cottonseed Flour on Growth Performance, Feed Utilization and Body Composition of Juvenile Red Drum Sciaenops Ocellatus. Aquaculture 2020, 523, 735217. [Google Scholar] [CrossRef]
- Xue, M. Cottonseed Protein Concentrate in the Processing Technology and Its Application in Aquatic Feed Nutritional Value. Feed Ind. 2021, 42, 633. [Google Scholar] [CrossRef]
- Chen, G.; Yin, B.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S. Effects of Fishmeal Replacement with Cottonseed Protein Concentrate on Growth, Digestive Proteinase, Intestinal Morphology and Microflora in Pearl Gentian Grouper (♀Epinephelus Fuscoguttatus × ♂Epinephelus Lanceolatu). Aquac. Res. 2020, 51, 2870–2884. [Google Scholar] [CrossRef]
- Ye, G.; Dong, X.; Yang, Q.; Chi, S.; Liu, H.; Zhang, H.; Tan, B.; Zhang, S. Low-Gossypol Cottonseed Protein Concentrate Used as a Replacement of FishMeal for Juvenile Hybrid Grouper (Epinephelus Fuscoguttatus ♀ × Epinephelus Lanceolatus ♂): Effects on Growth Performance, Immune Responses and Intestinal Microbiota. Aquaculture 2020, 524, 735309. [Google Scholar] [CrossRef]
- Zhao, W.; Liu, Z.; Niu, J. Growth Performance, Intestinal Histomorphology, Body Composition, Hematological and Antioxidant Parameters of Oncorhynchus Mykiss Were Not Detrimentally Affected by Replacement of FishMeal with Concentrated Dephenolization Cottonseed Protein. Aquac. Rep. 2021, 19, 100557. [Google Scholar] [CrossRef]
- Yin, B.; Liu, H.; Tan, B.; Dong, X.; Chi, S.; Yang, Q.; Zhang, S.; Chen, L. Cottonseed Protein Concentrate (CPC) Suppresses Immune Function in Different Intestinal Segments of Hybrid Grouper ♀Epinephelus Fuscoguttatus × ♂Epinephelus Lanceolatu via TLR-2/MyD88 Signaling Pathways. Fish Shellfish Immunol. 2018, 81, 318–328. [Google Scholar] [CrossRef]
- Wang, H.; Hu, X.; Zheng, Y.; Chen, J.; Tan, B.; Shi, L.; Zhang, S. Effects of Replacing FishMeal with Cottonseed Protein Concentrate on the Growth, Immune Responses, Digestive Ability and Intestinal Microbial Flora in Litopenaeus Vannamei. Fish Shellfish Immunol. 2022, 128, 91–100. [Google Scholar] [CrossRef]
- Jiang, S.; Chen, Z.; Zhou, F.; Yang, Q.; Huang, J.; Yang, L.; Li, Y.; Jiang, S. Study on Partial Replacement of FishMeal with Concentrated Dephenolized Cottonseed Protein in Feed of Penaeus Monodon. Aquac. Res. 2021, 52, 3871–3881. [Google Scholar] [CrossRef]
- Wan, M.; Yin, P.; Fang, W.; Xie, S.; Chen, S.J.; Tian, L.X.; Niu, J. The Effect of Replacement of Fishmeal by Concentrated Dephenolization Cottonseed Protein on the Growth, Body Composition, Haemolymph Indexes and Haematological Enzyme Activities of the Pacific White Shrimp (Litopenaeus vannamei). Aquac. Nutr. 2018, 24, 1845–1854. [Google Scholar] [CrossRef]
- Hou, C.; Song, S.; Zhu, L.; Shi, L.; Tan, B.; Zhang, S. Growth, Immunity, and Transcriptome Response to Dietary Wnt/β-Catenin Pathway Activator TWS119 in Penaeus Vannamei. Front. Mar. Sci. 2021, 8, 685429. [Google Scholar] [CrossRef]
- Yin, X.; Zhuang, X.; Liao, M.; Huang, L.; Cui, Q.; Liu, C.; Dong, W.; Wang, F.; Liu, Y.; Wang, W. Transcriptome Analysis of Pacific White Shrimp (Litopenaeus Vannamei) Hepatopancreas Challenged by Vibrio Alginolyticus Reveals Lipid Metabolic Disturbance. Fish Shellfish Immunol. 2022, 123, 238–247. [Google Scholar] [CrossRef] [PubMed]
- Trapnell, C.; Williams, B.; Pertea, G.; Mortazavi, A.; Kwan, G.; Van Baren, M.; Salzberg, S.; Wold, B.; Pachter, L. Transcript Assembly and Quantification by RNA-Seq Reveals Unannotated Transcripts and Isoform Switching during Cell Differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xiao, B.; Liao, X.; Wang, H.; He, J.; Li, C. BigPEN, an Antimicrobial Peptide of Penaeidin Family from Shrimp Litopenaeus Vannamei with Membrane Permeable and DNA Binding Activity. Fish Shellfish Immunol. Rep. 2021, 2, 100034. [Google Scholar] [CrossRef] [PubMed]
- Batista, S.; Medina, A.; Pires, M.; Moriñigo, M.; Sansuwan, K.; Fernandes, J.; Valente, L.; Ozório, R. Innate Immune Response, Intestinal Morphology and Microbiota Changes in Senegalese Sole Fed Plant Protein Diets with Probiotics or Autolysed Yeast. Appl. Microbiol. Biotechnol. 2016, 100, 7223–7238. [Google Scholar] [CrossRef]
- Roy, S.; Baruah, K.; Bossier, P.; Vanrompay, D.; Norouzitallab, P. Induction of Transgenerational Innate Immune Memory against Vibrio Infections in a Brine Shrimp (Artemia Franciscana) Model. Aquaculture 2022, 557, 738309. [Google Scholar] [CrossRef]
- Ruvalcaba-Márquez, J.; Álvarez-Ruíz, P.; Zenteno-Savín, T.; Martínez-Antonio, E.; Goytortúa-Bores, E.; Casillas-Hernández, R.; Mejía-Ruíz, H.; Magallón-Barajas, F. Performance, Immune Response, and Oxidative Stress Parameters of Litopenaeus Vannamei Fed Diets Containing Varying Carbohydrate/Protein, Lipid/Protein, and Energy/Protein Ratios. Aquac. Rep. 2021, 21, 100771. [Google Scholar] [CrossRef]
- Zhu, X.; Deng, Q.; Guo, H.; Li, G.; Zhu, C. Effects of Hydrolyzed Tannin on Blood and Blood Cell Immune Indexes of Vibrio Parahaemolyticus Infected Litopenaeus Vannamei. J. Guangdong Ocean Univ. 2021, 41, 12–19. [Google Scholar]
- Abdel-Latif, H.; Yilmaz, E.; Dawood, M.; Ringø, E.; Ahmadifar, E.; Yilmaz, S. Shrimp Vibriosis and Possible Control Measures Using Probiotics, Postbiotics, Prebiotics, and Synbiotics: A Review. Aquaculture 2022, 551, 737951. [Google Scholar] [CrossRef]
- Vazirzadeh, A.; Marhamati, A.; Rabiee, R.; Faggio, C. Immunomodulation, Antioxidant Enhancement and Immune Genes Up-Regulation in Rainbow Trout (Oncorhynchus Mykiss) Fed on Seaweeds Included Diets. Fish Shellfish Immunol. 2020, 106, 852–858. [Google Scholar] [CrossRef]
- Ahmadifar, E.; Sadegh, T.; Dawood, M.; Dadar, M.; Sheikhzadeh, N. The Effects of Dietary Pediococcus Pentosaceus on Growth Performance, Hemato-Immunological Parameters and Digestive Enzyme Activities of Common Carp (Cyprinus Carpio). Aquaculture 2020, 516, 734656. [Google Scholar] [CrossRef]
- Bogusz, D.; Appleby, C.; Landsmann, J.; Dennis, E.; Trinick, M.; Peacock, W. Functioning Haemoglobin Genes in Non-Nodulating Plants. Nature 1988, 331, 178–180. [Google Scholar] [CrossRef]
- Czelusniak, J.; Goodman, M.; Hewett Emmett, D.; Weiss, M.; Venta, P.; Tashian, E. Phylogenetic Origins and Adaptive Evolution of Avian and Mammalian Haemoglobin Genes. Nature 1982, 298, 297–300. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.; Yao, W.; Yang, H.; Tan, S.; Leng, X.; Li, X. Dietary Effects of Clostridium Autoethanogenum Protein Substituting FishMeal on Growth, Intestinal Histology and Immunity of Pacific White Shrimp (Litopenaeus Vannamei) Based on Transcriptome Analysis. Fish Shellfish Immunol. 2021, 119, 635–644. [Google Scholar] [CrossRef]
- Zhang, W.; Liu, K.; Tan, B.; Liu, H.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S.; Wang, H. Transcriptome, Enzyme Activity and Histopathology Analysis Reveal the Effects of Dietary Carbohydrate on Glycometabolism in Juvenile Largemouth Bass, Micropterus Salmoides. Aquaculture 2019, 504, 39–51. [Google Scholar] [CrossRef]
- Zhang, S.; Zhu, L.; Hou, C.; Yuan, H.; Yang, S.; Dehwah, M.; Shi, L. GSK3β Plays a Negative Role During White Spot Syndrome Virus (WSSV) Infection by Regulating NF-ΚB Activity in Shrimp Litopenaeus Vannamei. Front. Immunol. 2020, 11, 607543. [Google Scholar] [CrossRef]
- Li, C.; Wang, S.; He, J. The Two NF-ΚB Pathways Regulating Bacterial and WSSV Infection of Shrimp. Front. Immunol. 2019, 10, 1785. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, W.; Lin, X.; Wang, C.; Ke, J.; Wang, L.; Liu, H. Transcriptome of White Shrimp Litopenaeus Vannamei Induced with Rapamycin Reveals the Role of Autophagy in Shrimp Immunity. Fish Shellfish Immunol. 2019, 86, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Tassanakajon, A.; Somboonwiwat, K. Antimicrobial Peptides from the Black Tiger Shrimp Penaeus Monodon—A Review. Dis. Asian Aquac. VII 2011, 229–240. [Google Scholar]
- Li, C.; Chen, Y.; Zhang, S.; Lü, L.; Chen, Y.; Chai, J.; Weng, S.; Chen, Y.; He, J.; Xu, X. Identification, Characterization, and Function Analysis of the Cactus Gene from Litopenaeus Vannamei. PLoS ONE 2012, 7, e49711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, M.; Li, S.; Jin, S.; Li, X.; Xiang, J.; Li, F. A Novel TRIM9 Protein Promotes NF-ΚB Activation through Interacting with LvIMD in Shrimp during WSSV Infection. Front. Immunol. 2022, 13, 526. [Google Scholar] [CrossRef]
- Ernst, M.; Dunn, L.; Rice, N. The PEST-like Sequence of I Kappa B Alpha Is Responsible for Inhibition of DNA Binding but Not for Cytoplasmic Retention of c-Rel or RelA Homodimers. Mol. Cell. Biol. 1995, 15, 872–882. [Google Scholar] [CrossRef] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredient | Group | |||
|---|---|---|---|---|
| FM | CPC15 | CPC30 | CPC45 | |
| Fishmeal | 25 | 21.25 | 17.5 | 13.75 |
| Soybean meal | 25 | 25 | 25 | 25 |
| Peanut meal | 8.92 | 8.92 | 8.92 | 8.92 |
| Flour | 18 | 18 | 18 | 18 |
| Brewer’s yeast | 2.84 | 2.84 | 2.84 | 2.84 |
| Shrimp head powder | 5.53 | 5.53 | 5.53 | 5.53 |
| Cottonseed protein concentrate | 0 | 4.39 | 8.77 | 13.16 |
| Fish oil | 2 | 2 | 2 | 2 |
| Soybean oil | 2.21 | 2.46 | 2.72 | 2.98 |
| Choline | 0.3 | 0.3 | 0.3 | 0.3 |
| Soy lecithin | 1 | 1 | 1 | 1 |
| Vitamin and mineral premix a | 1 | 1 | 1 | 1 |
| Choline chloride | 1.5 | 1.5 | 1.5 | 1.5 |
| Vitamin C | 0.1 | 0.1 | 0.1 | 0.1 |
| Microcrystalline cellulose | 6.55 | 5.66 | 4.77 | 3.87 |
| Antioxidant | 0.05 | 0.05 | 0.05 | 0.05 |
| Total | 100 | 100 | 100 | 100 |
| Proximate composition | ||||
| Crude protein | 38.11 | 38.47 | 38.21 | 38.37 |
| Ether extract | 6.74 | 6.73 | 6.78 | 6.76 |
| Moisture | 7.52 | 7.61 | 7.59 | 7.46 |
| Ash | 10.87 | 11.80 | 11.48 | 10.80 |
| Item | FM | CPC15 | CPC30 | CPC45 |
|---|---|---|---|---|
| Asp | 3.51 | 3.53 | 3.6 | 3.71 |
| Thr | 1.48 | 1.43 | 1.44 | 1.34 |
| Ser | 1.65 | 1.59 | 1.64 | 1.64 |
| Glu | 6.45 | 6.68 | 6.94 | 7.1 |
| Gly | 1.92 | 1.89 | 1.84 | 1.88 |
| Ala | 1.87 | 1.83 | 1.76 | 1.83 |
| Cys | 0.42 | 0.45 | 0.43 | 0.45 |
| Val | 1.63 | 1.65 | 1.64 | 1.61 |
| Met | 0.59 | 0.57 | 0.54 | 0.46 |
| Ile | 1.44 | 1.45 | 1.44 | 1.45 |
| Leu | 2.59 | 2.58 | 2.57 | 2.57 |
| Tyr | 0.98 | 0.98 | 1.06 | 0.96 |
| Phe | 1.64 | 1.72 | 1.78 | 1.53 |
| Lys | 2.29 | 2.2 | 2.14 | 2.12 |
| His | 0.97 | 0.97 | 1.02 | 1.11 |
| Arg | 2.38 | 2.57 | 2.79 | 3.12 |
| Pro | 1.82 | 1.82 | 1.86 | 1.77 |
| Total amino acids | 33.63 | 33.91 | 34.49 | 34.65 |
| Gene ID No. | Primer Name | Sequence (5′-3′) |
|---|---|---|
| GU136229 | EF1α-F | GAAGTAGCCGCCCTGGTTG |
| EF1α-R | CGGTTAGCCTTGGGGTTGAG | |
| DQ005531 | SOD-F | CTTTGCCACCCCTCAAGTATG |
| SOD-R | TGCCTCCGCCTCAACCA | |
| XM027381766 | PO-F | AAGCCAGGCAGCAACCAC |
| PO-R | CAGAAGTTGAAACCCGTGGC | |
| AF425673 | LZM-F | TATTCTGCCTGGGTGGCTTAC |
| LZM-R | CAGAGTTGGAACCGTGAGACC | |
| JX162772 | CAT-F | ACAAGACGGACCAAGGCATC |
| CAT-R | ACTTATATCCTCAGTGACTGGCATG | |
| KR676449 | ACP-F | TCAAGACCTGGAAGAATGGGACT |
| ACP-R | CCTCTCTGAACCTCCTCCTGTAAC | |
| XM027353927 | AKP-F | TACGGCTACCGAAGGGACG |
| AKP-R | AGTCGGTGTTGTTCGTGTTGG | |
| XM027365017 | AST-F | AGATGGACCAGCGTGTGATTAC |
| AST-R | TCGCCTTGGTCACGCTGT | |
| XM027364929 | ALT-F | TCTCCGCATCGGCTCCA |
| ALT-R | CCACAGGTCTTTGGGTCGTAG | |
| ROT83547.1 | phc1-F | CGAAGGTCGCTGGAATGC |
| phc1-R | ATTCCGTGGACTTGCGTTC | |
| ROT81694.1 | Lac2-F | CAGATGGTAACCCCGTTGTGC |
| Lac2-R | GAAGGTTCCGACTTTGGTGTTTT | |
| ROT80984.1 | Trilip-F | CGGCTTTATGGAGGGGATG |
| Trilip-R | ACTCGTTCGTCACAGGCATAAG | |
| ROT69062.1 | Sp2-F | GCGTGGAAGGGAAGTGGG |
| Sp2-R | TTCGGCTCACGGGCTTCT | |
| ROT66027.1 | GTP-F | TGCTGTCCAACACCATCTTCAC |
| GTP-R | ACATCGTTCGCCGTTTCCT | |
| ROT63397.1 | NFBI-F | ACTCCTTGCCGCTGACCTTAC |
| NFBI-R | GTGCCGTCCGACCACTCTT | |
| ROT85909.1 | Brp3-F | GCGAGAGGTCACAGGAGAGC |
| Brp3-R | CACTCGTTCTTCAGCCCTTG | |
| MSTRG.2558.1 | 2558F | CCGTGTCAACCAAACCAGAAG |
| 2558R | CATTCAGACAAATCACACTCCACA | |
| ROT75114.1 | SC28-F | CGAGAAGGTGGGCGAACTGA |
| SC28-R | CAGGTTGATGCCGATGGAGC | |
| MSTRG.13279.1 | 13279F | GTCCACGGTAGTCAAGCAAGC |
| 13279R | TCTCTCTACCAGATGCTGTGAAAGT | |
| MSTRG.23082.1 | 23082F | GAATGGACGGTGAAAACACTTATG |
| 23082R | TGCCTGGATGGGTAGGTGTG | |
| ROT81705.1 | Lys-F | TACTGGTGCGGAAGCGACTA |
| Lys-R | GTAAGCCACCCAGGCAGAATA | |
| ROT77417.1 | E75-F | GACCAAGCATCACCCCGAGA |
| E75-R | GCGGTGTGCTCAGTCATCTTGTA | |
| ROT70303.1 | Naa-F | CTACAACTCCTTCCGCCACAA |
| Naa-R | ATGGCGAAGGTGGTGAAGC | |
| ROT67869.1 | GABA-F | CATTGGCTCCCCTCTGGTC |
| GABA-R | CGAAGGTAGTCGGGGAACAA | |
| ROT62527.1 | 62527F | CGAACGCAAAGGCAGCAC |
| 62527R | CGACCCTTGTTGCTTCCTCC | |
| LOC113823636 | Cactus-F | GCGGACGAAGACCTTGACT |
| Cactus-R | GGCTGATGGTCATTCACTTGC | |
| DQ206401.1 | PEN2-F | GACGGAGAAGACAATGGAAACC |
| PEN2-R | ATCTTTAGCGATGGATAGACGAA | |
| DQ206403.1 | Pen3-F | TACAACGGTTGCCCTGTCTCA |
| Pen3-R | ACCGGAATATCCCTTTCCCAC | |
| DQ206402.1 | Pen4-F | GGTGCGATGTATGCTACGGAA |
| Pen4-R | CATCGTCTTCTCCATCAACCA | |
| AF430071.1 | Cru1-F | GTAGGTGTTGGTGGTGGTTTC |
| Cru1-R | CTCGCAGCAGTAGGCTTGAC | |
| AY465833.1 | Cru3-F | TCCACAATGGTCAGCGTCAAG |
| Cru3-R | CTGTCCGACAAGCAGTTCCTC | |
| EW713395 | ALF1-F | TTACTTCAATGGCAGGATGTGG |
| ALF1-R | GTCCTCCGTGATGAGATTACTCTG | |
| EW713396 | ALF2-F | GGCCATTGCGAACAAACTCAC |
| ALF2-R | GTCCATCCTGGGCACCACAT | |
| ABB22831 | ALF3-F | CTCCGTGTTGACAAGCCTGG |
| ALF3-R | GCAGCTCCGTCTCCTCGTTC |
| Parameter | Group | |||
|---|---|---|---|---|
| FM | CPC15 | CPC30 | CPC45 | |
| ACP (IU/g) | 92.95 ± 0.92 a | 108.60 ± 3.17 b | 140.90 ± 2.40 c | 90.53 ± 1.72 a |
| AKP (IU/g) | 66.67 ± 1.32 a | 120.34 ± 4.52 b | 106.27 ± 3.78 b | 120.66 ± 2.77 b |
| SOD (U/mL) | 96.29 ± 0.54 a | 145.59 ± 3.28 b | 141.40 ± 4.23 b | 137.94 ± 3.24 b |
| PO (U/L) | 34.31 ± 0.64 | 34.30 ± 2.46 | 38.80 ± 2.12 | 36.14 ± 0.78 |
| LZM (U/L) | 2.92 ± 0.04 a | 2.92 ± 0.12 a | 4.90 ± 0.10 b | 4.26 ± 0.08 ab |
| CAT (U/mL) | 40.81 ± 1.21 a | 58.08 ± 1.32 b | 52.24 ± 0.38 b | 53.56 ± 0.47 b |
| AST (mU/g) | 44.75 ± 0.07 b | 27.35 ± 0.16 a | 29.16 ± 0.09 a | 44.41 ± 1.41 b |
| ALT (mU/g) | 74.78 ± 2.34 b | 72.75 ± 2.67 b | 58.59 ± 1.57 a | 88.91 ± 4.35 c |
| MDA (nmol/mL) | 10.70 ± 0.05 c | 6.36 ± 0.07 a | 8.04 ± 0.15 b | 7.93 ± 0.32 b |
| Item | Raw Data (bp) | Clean Data (bp) | GC Percentage (bp) | Q20% (bp) | Q30% (bp) | Effective Reads (bp) | Total Mapped (bp) | Exon (bp) | Intron (bp) | Intergenic (bp) |
|---|---|---|---|---|---|---|---|---|---|---|
| FM-1 | 7,750,691,400 | 7,667,795,876 | 3,561,592,079 (45.95%) | 7,500,430,776 (97.82%) | 7,185,399,849 (93.71%) | 51,526,760 | 41,699,138 (85.49%) | 26,675,562 (63.97%) | 3,204,123 (7.68%) | 11,819,453 (28.34%) |
| FM-2 | 6,725,523,000 | 6,642,429,430 | 3,134,589,163 (46.61%) | 6,481,225,567 (97.57%) | 6,181,988,334 (93.07%) | 44,694,126 | 37,060,603 (86.07%) | 23,845,520 (64.34%) | 2,734,914 (7.38%) | 10,480,169 (28.28%) |
| FM-3 | 6,593,927,100 | 6,513,279,775 | 3,068,267,760 (46.53%) | 6,358,061,273 (97.62%) | 6,073,534,860 (93.25%) | 43,825,148 | 35,434,954 (85.21%) | 22,983,166 (64.86%) | 2,730,666 (7.71%) | 9,721,122 (27.43%) |
| CPC15-1 | 7,156,347,000 | 7,072,502,810 | 3,253,038,517 (45.46%) | 6,906,298,032 (97.65%) | 6,590,590,898 (93.19%) | 47,571,384 | 38,572,370 (85.00%) | 24,381,187 (63.21%) | 2,984,675 (7.74%) | 11,206,508 (29.05%) |
| CPC15-2 | 6,818,080,800 | 6,736,566,160 | 3,103,084,796 (45.51%) | 6,573,194,948 (97.57%) | 6,263,533,549 (92.98%) | 45,334,742 | 36,034,745 (84.43%) | 22,764,691 (63.17%) | 2,787,220 (7.73%) | 10,482,834 (29.09%) |
| CPC15-3 | 7,049,986,800 | 6,974,039,570 | 3,180,898,090 (45.12%) | 6,808,099,463 (97.62%) | 6,497,966,411 (93.17%) | 46,870,694 | 37,195,349 (84.58%) | 23,388,113 (62.88%) | 2,996,288 (8.06%) | 10,810,948 (29.07%) |
| CPC30-1 | 8,211,688,800 | 8,120,317,400 | 3,765,592,526 (45.86%) | 7,933,918,636 (97.70%) | 7,581,072,924 (93.36%) | 54,588,588 | 44,193,568 (85.59%) | 28,414,814 (64.30%) | 3,391,737 (7.67%) | 12,387,017 (28.03%) |
| CPC30-2 | 7,362,995,400 | 7,282,017,926 | 3,402,468,398 (46.21%) | 7,106,052,798 (97.58%) | 6,780,633,929 (93.11%) | 48,933,996 | 40,421,630 (85.86%) | 25,501,372 (63.09%) | 3,078,600 (7.62%) | 11,841,658 (29.30%) |
| CPC30-3 | 7,383,816,300 | 7,309,068,344 | 3,356,616,410 (45.46%) | 7,142,935,453 (97.73%) | 6,828,866,924 (93.43%) | 49,102,968 | 39,634,331 (85.22%) | 25,509,188 (64.36%) | 3,152,083 (7.95%) | 10,973,060 (27.69%) |
| CPC45-1 | 7,461,683,700 | 7,381,254,347 | 3,456,162,821 (46.32%) | 7,212,569,018 (97.71%) | 6,896,396,008 (93.43%) | 49,604,658 | 40,140,871 (85.62%) | 24,365,099 (64.84%) | 2,849,807 (7.58%) | 10,361,522 (27.57%) |
| CPC45-2 | 7,334,094,900 | 7,262,493,436 | 3,227,802,467 (44.01%) | 7,111,478,739 (97.92%) | 6,813,670,715 (93.82%) | 48,787,044 | 37,576,428 (83.57%) | 24,275,268 (63.78%) | 3,000,806 (7.88%) | 10,782,327 (28.33%) |
| CPC45-3 | 6,898,534,800 | 6,822,192,973 | 3,192,487,607 (46.28%) | 6,650,991,010 (97.49%) | 6,332,191,861 (92.82%) | 45,841,774 | 38,058,401 (86.38%) | 23,914,820 (65.83%) | 2,895,969 (7.97%) | 9,519,324 (26.20%) |
| Category or Gene ID | Gene Description | Species | Log2FC a | Log2FC b | Log2FC c |
|---|---|---|---|---|---|
| Toxoplasmosis | |||||
| ROT61665.1 | Cactus | Penaeus vannamei | −0.22 | −9.16 | −9.16 |
| ROT61785.1 | Hsc71 | Penaeus vannamei | −0.61 | −0.79 | −0.87 |
| ROT67561.1 | Hsc71 | Penaeus vannamei | −0.84 | −0.89 | −0.87 |
| Pathogenic Escherichia coli infection | |||||
| ROT61665.1 | Cactus | Penaeus vannamei | −0.22 | −9.16 | −9.16 |
| ROT62386.1 | Cycs | Penaeus vannamei | −0.84 | −0.55 | −0.54 |
| ROT67928.1 | Tuba1a | Gecarcinus lateralis | −1.74 | −1.84 | −2.48 |
| ROT62891.1 | Tbb-4 | Penaeus vannamei | 0.77 | 0.84 | 1.72 |
| Insulin resistance | |||||
| ROT61665.1 | Cactus | Penaeus vannamei | −0.22 | −9.16 | −9.16 |
| ROT83272.1 | Slc2a3 | Penaeus vannamei | −1.50 | −1.66 | −1.23 |
| ROT74164.1 | Mlxipl | Penaeus vannamei | −0.69 | −1.16 | −1.01 |
| Prostate cancer | |||||
| ROT61665.1 | Cactus | Penaeus vannamei | −0.22 | −9.16 | −9.16 |
| ROT63374.1 | Hsp90b1 | Penaeus vannamei | −0.92 | −1.21 | −1.22 |
| Toll and IMD pathway | |||||
| ROT61665.1 | Cactus | Penaeus vannamei | −0.22 | −9.16 | −9.16 |
| ROT79438.1 | Prss8 | Penaeus vannamei | −0.92 | −1.22 | −1.21 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Hu, X.; Chen, J.; Yuan, H.; Hu, N.; Tan, B.; Dong, X.; Zhang, S. Transcriptome Analysis Reveals the Immunoregulation of Replacing Fishmeal with Cottonseed Protein Concentrates on Litopenaeus vannamei. Animals 2023, 13, 1185. https://doi.org/10.3390/ani13071185
Wang H, Hu X, Chen J, Yuan H, Hu N, Tan B, Dong X, Zhang S. Transcriptome Analysis Reveals the Immunoregulation of Replacing Fishmeal with Cottonseed Protein Concentrates on Litopenaeus vannamei. Animals. 2023; 13(7):1185. https://doi.org/10.3390/ani13071185
Chicago/Turabian StyleWang, Hongming, Xin Hu, Jian Chen, Hang Yuan, Naijie Hu, Beiping Tan, Xiaohui Dong, and Shuang Zhang. 2023. "Transcriptome Analysis Reveals the Immunoregulation of Replacing Fishmeal with Cottonseed Protein Concentrates on Litopenaeus vannamei" Animals 13, no. 7: 1185. https://doi.org/10.3390/ani13071185
APA StyleWang, H., Hu, X., Chen, J., Yuan, H., Hu, N., Tan, B., Dong, X., & Zhang, S. (2023). Transcriptome Analysis Reveals the Immunoregulation of Replacing Fishmeal with Cottonseed Protein Concentrates on Litopenaeus vannamei. Animals, 13(7), 1185. https://doi.org/10.3390/ani13071185