
Tissue-Specific Modulation of Gluco- and Growth-Regulatory Factor Abundance by Nesfatin-1 and Nesfatin-1-like Peptide in Goldfish

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal and Ethics
2.2. Peptides and Reagents
2.3. In Vivo Studies
2.4. In Vitro Studies
2.5. Total RNA Extraction, cDNA Synthesis, and qPCR
2.6. Statistical Analysis
3. Results
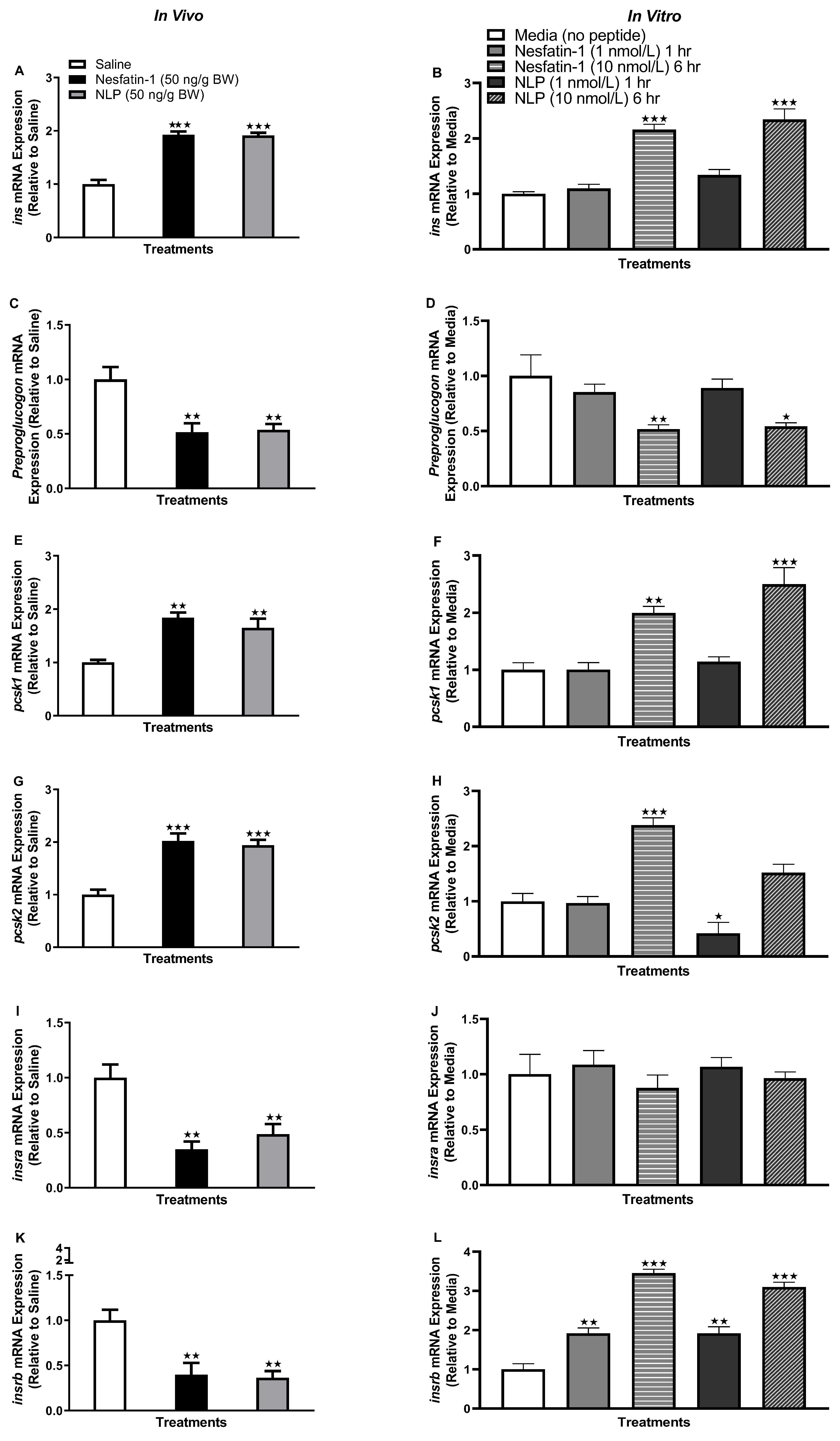
3.1. Nesfatin-1 and Nlp Stimulated Insulin mRNA Abundance in Goldfish Adipose Tissue
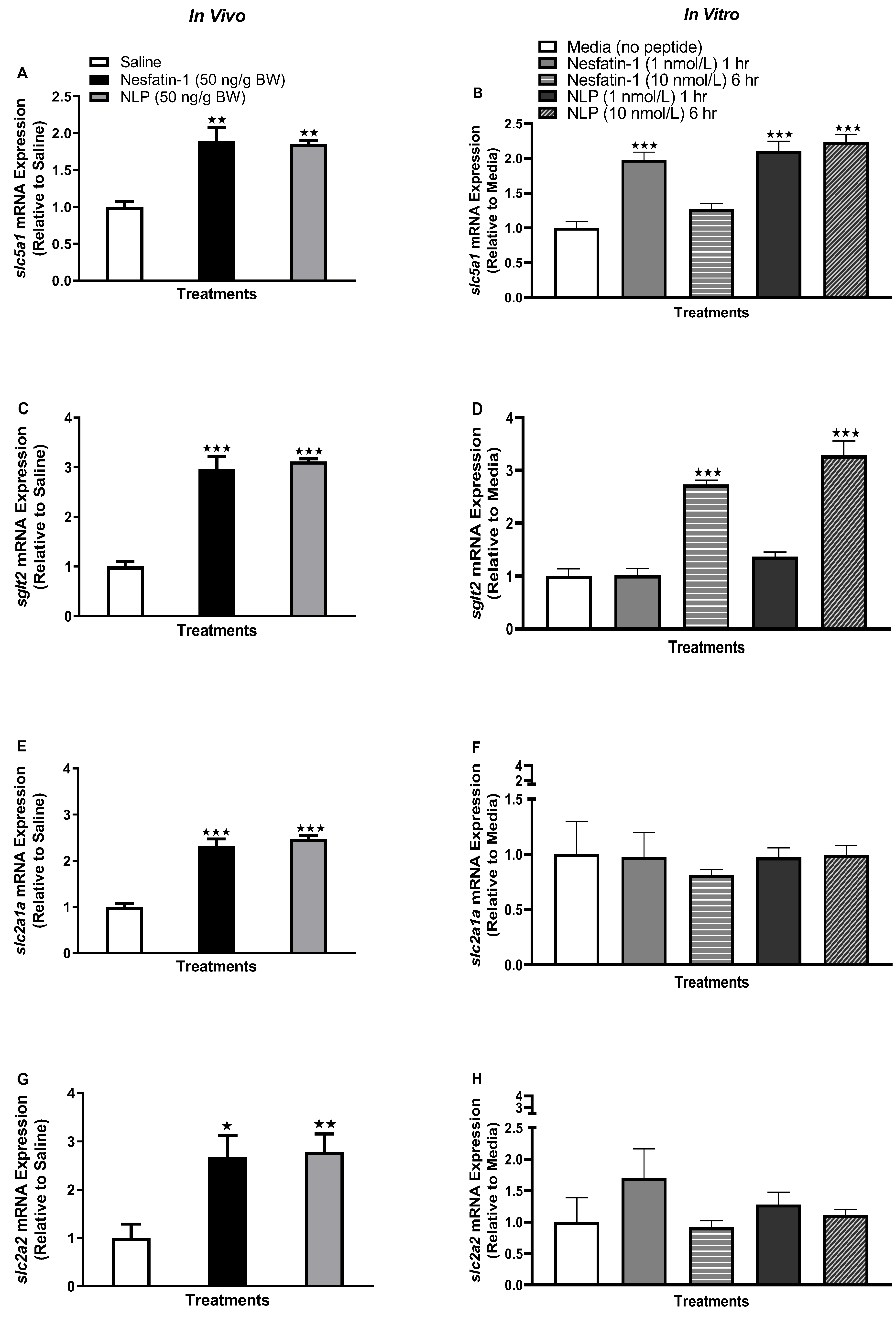
3.2. Nesfatin-1 and Nlp Stimulated Glucose Transporter Transcript Abundance in Goldfish Adipose Tissue
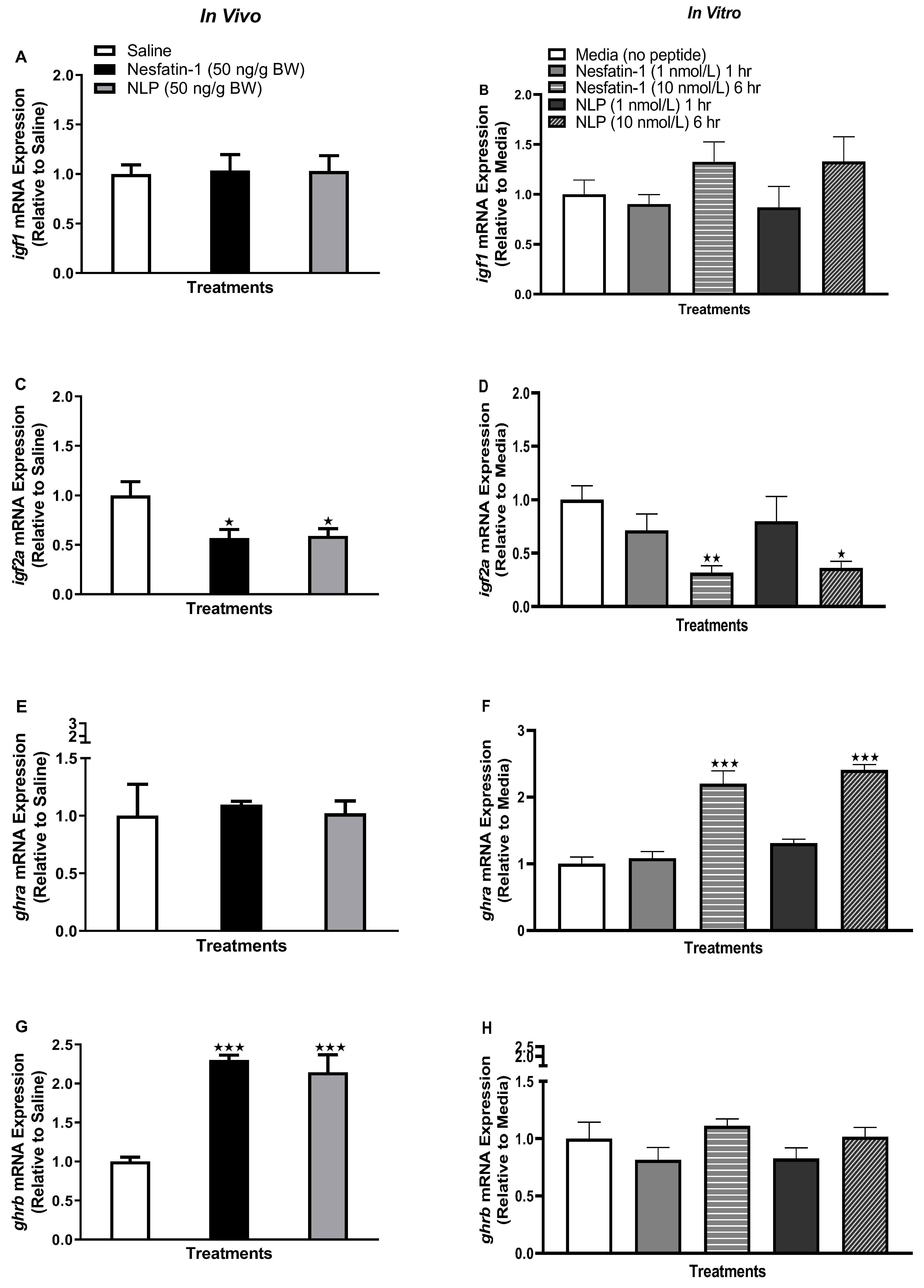
3.3. Nesfatin-1 and Nlp Modulated an ghr-igf System mRNA Abundance in Goldfish Adipose Tissue
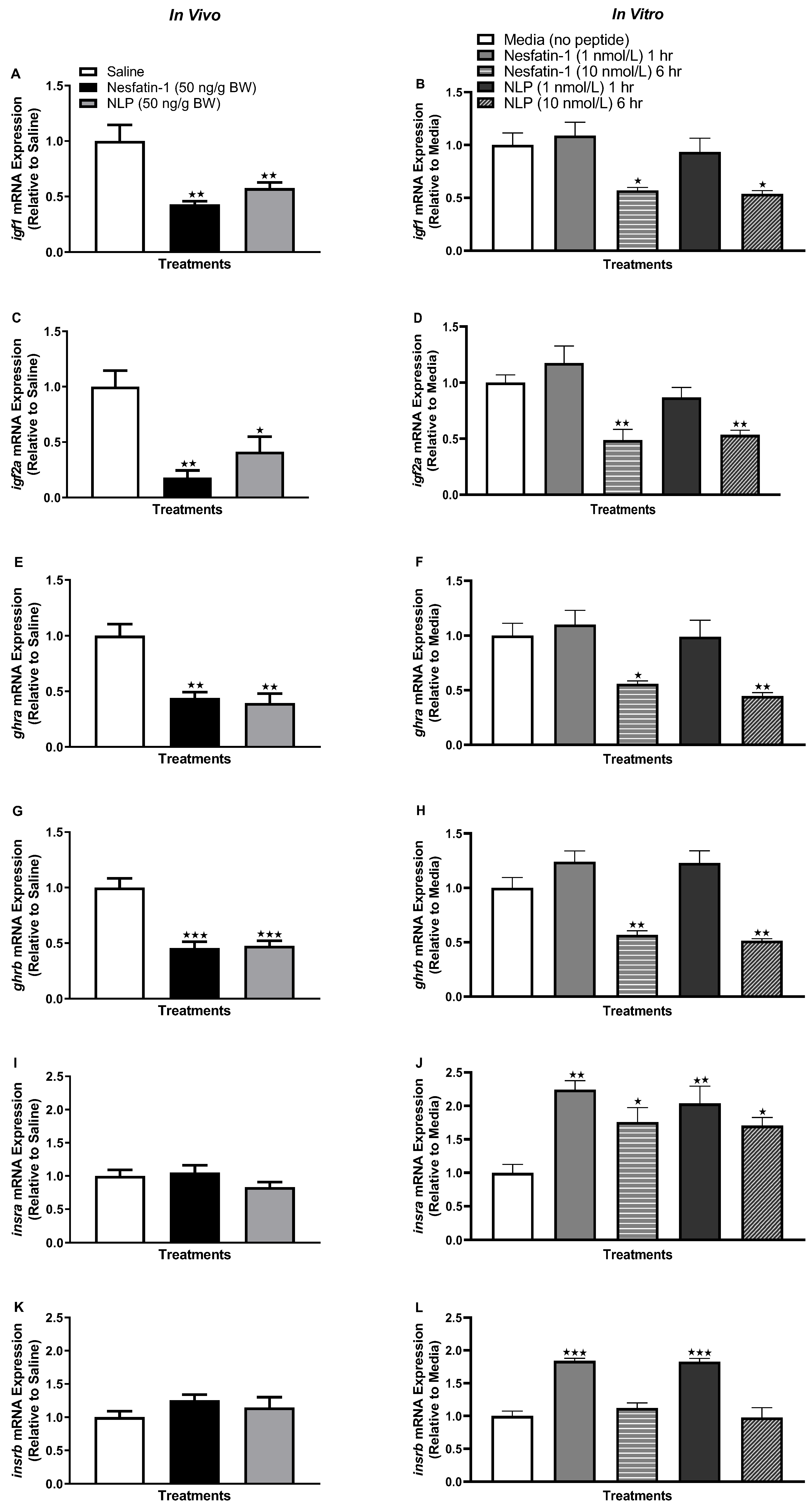
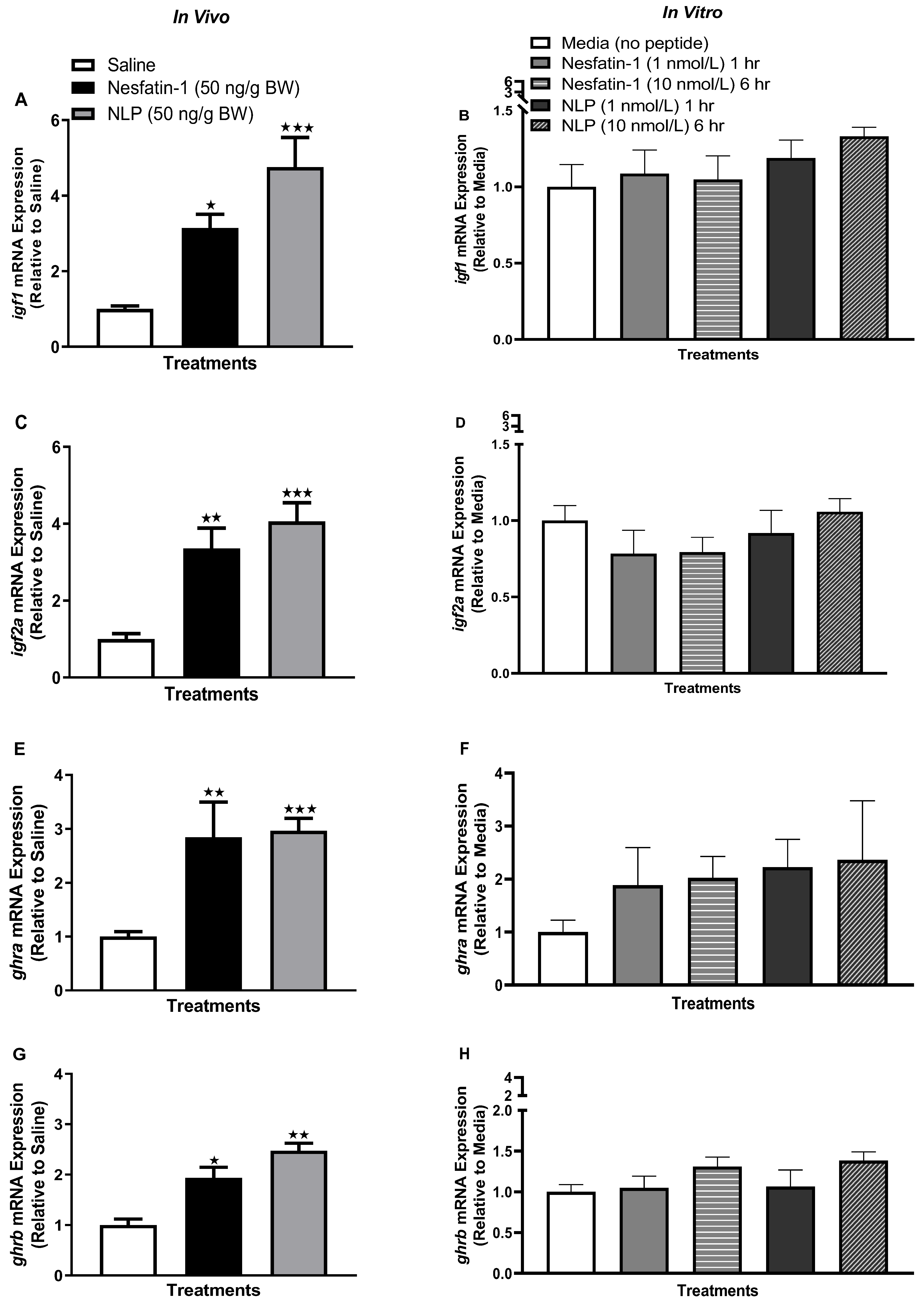
3.4. Nesfatin-1 and Nlp Modulated ghr-igf System in Goldfish Liver
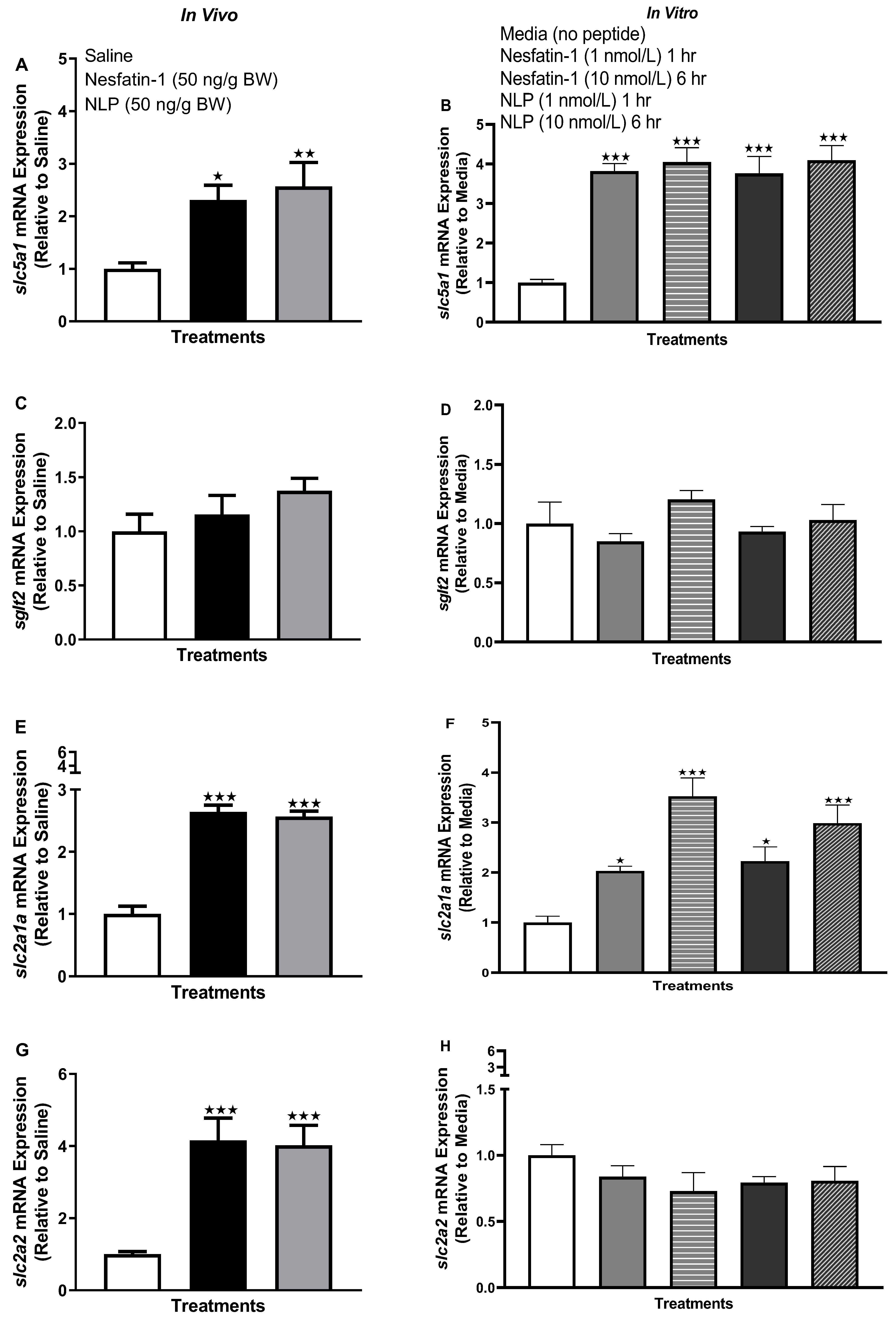
3.5. Nesfatin-1 and Nlp Differently Modulate the mRNA Abundance of Glucose Transporters in Goldfish Liver
3.6. IP Injection, Not In Vitro Treatment with Nesfatin-1 or Nlp, Upregulated the mRNA Abundance of ghr-igf System in the Muscle of Goldfish
3.7. Nesfatin-1 and Nlp Upregulated the Transcript Abundance of Glucose Transporters in Goldfish Muscle
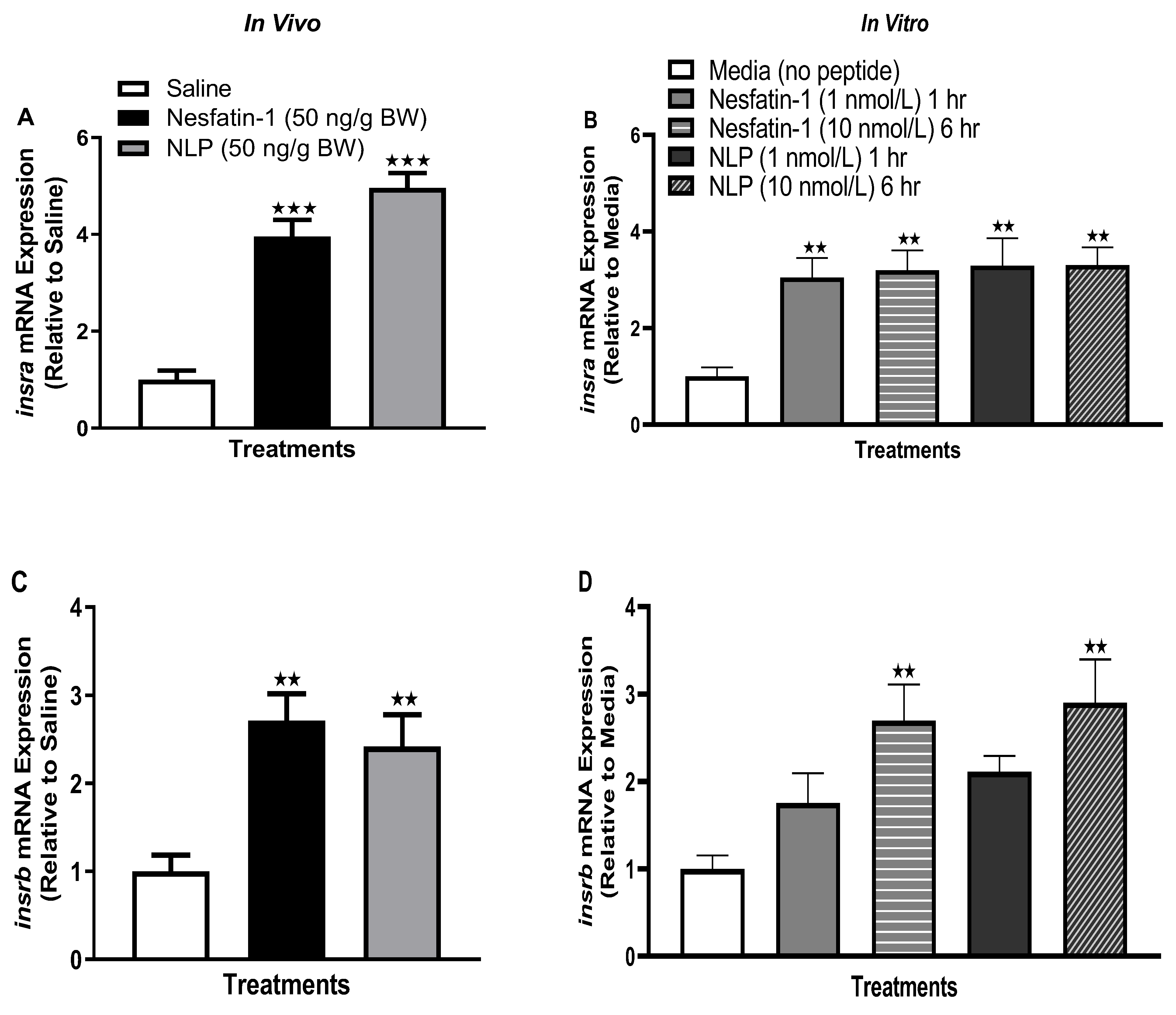
3.8. Nesfatin-1 and Nlp Upregulated the Insulin Receptor Transcript Abundance in Goldfish Muscle
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gonzalez, R.; Kerbel, B.; Chun, A.; Unniappan, S. Molecular, Cellular and Physiological Evidences for the Anorexigenic Actions of Nesfatin-1 in Goldfish. PLoS ONE 2010, 5, e15201. [Google Scholar] [CrossRef]
- Blanco, A.M.; Velasco, C.; Bertucci, J.I.; Soengas, J.L.; Unniappan, S. Nesfatin-1 Regulates Feeding, Glucosensing and Lipid Metabolism in Rainbow Trout. Front. Endocrinol. 2018, 9, 484. [Google Scholar] [CrossRef] [PubMed]
- Blanco, A.M.; Pemberton, J.G.; Gonzalez, R.; Hatef, A.; Pham, V.; Chang, J.P.; Unniappan, S. Nesfatin-1 Is an Inhibitor of the Growth Hormone-Insulin-like Growth Factor Axis in Goldfish (Carassius Auratus). J. Neuroendocrinol. 2021, 33, e13010. [Google Scholar] [CrossRef]
- Rajeswari, J.J.; Unniappan, S. Nesfatin-1 Suppresses Fish Reproductive Axis and Gonadal Steroidogenesis. Reproduction 2020, 160, 445–454. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Shepperd, E.; Thiruppugazh, V.; Lohan, S.; Grey, C.L.; Chang, J.P.; Unniappan, S. Nesfatin-1 Regulates the Hypothalamo-Pituitary-Ovarian Axis of Fish. Biol. Reprod. 2012, 87, 1–11. [Google Scholar] [CrossRef]
- Nair, N.; Gerger, C.; Hatef, A.; Weber, L.P.; Unniappan, S. Ultrasonography Reveals in Vivo Dose-Dependent Inhibition of End Systolic and Diastolic Volumes, Heart Rate and Cardiac Output by Nesfatin-1 in Zebrafish. Gen. Comp. Endocrinol. 2016, 234, 142–150. [Google Scholar] [CrossRef]
- Sundarrajan, L.; Blanco, A.M.; Bertucci, J.I.; Ramesh, N.; Canosa, L.F.; Unniappan, S. Nesfatin-1-Like Peptide Encoded in Nucleobindin-1 in Goldfish Is a Novel Anorexigen Modulated by Sex Steroids, Macronutrients and Daily Rhythm. Sci. Rep. 2016, 6, 28377. [Google Scholar] [CrossRef] [PubMed]
- Rajeswari, J.J.; Hatef, A.; Unniappan, S. Nesfatin-1-Like Peptide Suppresses Hypothalamo-Pituitary-Gonadal MRNAs, Gonadal Steroidogenesis and Oocyte Maturation in Fish. Biol. Reprod. 2020, 103, 802–816. [Google Scholar] [CrossRef]
- Pham, V.; Pemberton, J.G.; Chang, J.P.; Blanco, A.M.; Nasri, A.; Unniappan, S. Nesfatin-1 Stimulates the Hypothalamus-Pituitary-Interrenal Axis Hormones in Goldfish. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2021, 321, R603–R613. [Google Scholar] [CrossRef]
- Blanco, A.M.; Bertucci, J.I.; Unniappan, S. Goldfish Adipocytes Are Pancreatic Beta Cell-like, Glucose-Responsive Insulin-Producing Cells. J. Cell. Physiol. 2020, 235, 6875–6886. [Google Scholar] [CrossRef]
- Bottalico, L.N.; Korlyakova, J.; Weljie, A.M.; Habibi, H.R. Seasonally Related Disruption of Metabolism by Environmental Contaminants in Male Goldfish (Carassius Auratus). Front. Toxicol. 2021, 3, 750870. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez, R.; Unniappan, S. Molecular Characterization, Appetite Regulatory Effects and Feeding Related Changes of Peptide YY in Goldfish. Gen. Comp. Endocrinol. 2010, 166, 273–279. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Bretaño, A.; Callejo, M.; Montero, M.; Alonso-Gómez, Á.L.; Delgado, M.J.; Isorna, E. Performing a Hepatic Timing Signal: Glucocorticoids Induce Gper1a and Gper1b Expression and Repress Gclock1a and Gbmal1a in the Liver of Goldfish. J Comp. Physiol. B 2016, 186, 73–82. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Vélez, E.J.; Unniappan, S. Nesfatin-1 and Nesfatin-1-like Peptide Suppress Growth Hormone Synthesis via the AC/PKA/CREB Pathway in Mammalian Somatotrophs. Sci. Rep. 2020, 10, 16686. [Google Scholar] [CrossRef] [PubMed]
- Gawli, K.; Ramesh, N.; Unniappan, S. Nesfatin-1-like Peptide Is a Novel Metabolic Factor That Suppresses Feeding, and Regulates Whole-Body Energy Homeostasis in Male Wistar Rats. PLoS ONE 2017, 12, e0178329. [Google Scholar] [CrossRef] [PubMed]
- Sundarrajan, L.; Jayakumar Rajeswari, J.; Weber, L.P.; Unniappan, S. Nesfatin-1-like Peptide Is a Negative Regulator of Cardiovascular Functions in Zebrafish and Goldfish. Gen. Comp. Endocrinol. 2021, 313, 113892. [Google Scholar] [CrossRef]
- Ramesh, N.; Mohan, H.; Unniappan, S. Nucleobindin-1 Encodes a Nesfatin-1-like Peptide That Stimulates Insulin Secretion. Gen. Comp. Endocrinol. 2015, 216, 182–189. [Google Scholar] [CrossRef]
- Yang, Y.; Zhang, B.; Nakata, M.; Nakae, J.; Mori, M.; Yada, T. Islet β-Cell-Produced NUCB2/Nesfatin-1 Maintains Insulin Secretion and Glycemia along with Suppressing UCP-2 in β-Cells. J. Physiol. Sci. 2019, 69, 733–739. [Google Scholar] [CrossRef]
- Matta, R.A.; El-Hini, S.H.; Salama, A.M.S.E.; Moaness, H.M. Serum Nesfatin-1 Is a Biomarker of Pre-Diabetes and Interplays with Cardiovascular Risk Factors. Egypt. J. Intern. Med. 2022, 34, 15. [Google Scholar] [CrossRef]
- Zhai, T.; Li, S.-Z.; Fan, X.-T.; Tian, Z.; Lu, X.-Q.; Dong, J. Circulating Nesfatin-1 Levels and Type 2 Diabetes: A Systematic Review and Meta-Analysis. J. Diabetes Res. 2017, 2017, 7687098. [Google Scholar] [CrossRef]
- Durst, M.; Könczöl, K.; Ocskay, K.; Sípos, K.; Várnai, P.; Szilvásy-Szabó, A.; Fekete, C.; Tóth, Z.E. Hypothalamic Nesfatin-1 Resistance May Underlie the Development of Type 2 Diabetes Mellitus in Maternally Undernourished Non-Obese Rats. Front. Neurosci. 2022, 16, 828571. [Google Scholar] [CrossRef] [PubMed]
- Schalla, M.A.; Stengel, A. Current Understanding of the Role of Nesfatin-1. J. Endocr. Soc. 2018, 2, 1188–1206. [Google Scholar] [CrossRef] [PubMed]
- Bachle, L.A.; Smith, D.D.; Petzel, D. Isolation and Characterization of Insulin from the Brockmann Body of Dissostichus Mawsoni, an Antarctic Teleost Fish. J. Pept. Res. 2000, 56, 47–54. [Google Scholar] [CrossRef] [PubMed]
- Youson, J.H.; Al-Mahrouki, A.A.; Amemiya, Y.; Graham, L.C.; Montpetit, C.J.; Irwin, D.M. The Fish Endocrine Pancreas: Review, New Data, and Future Research Directions in Ontogeny and Phylogeny. Gen. Comp. Endocrinol. 2006, 148, 105–115. [Google Scholar] [CrossRef]
- Conde-Sieira, M.; Soengas, J.L. Nutrient Sensing Systems in Fish: Impact on Food Intake Regulation and Energy Homeostasis. Front. Neurosci. 2017, 10, 603. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.L.I.; Kostyniuk, D.J.; Mennigen, J.A.; Weber, J.-M. Unexpected Effect of Insulin on Glucose Disposal Explains Glucose Intolerance of Rainbow Trout. Am. J. Physiol. Regul. Integr. Comp. Physiol. 2019, 316, R387–R394. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Soengas, J.L.; Moon, T.W. Glucose Metabolism in Fish: A Review. J. Comp. Physiol. B 2012, 182, 1015–1045. [Google Scholar] [CrossRef]
- Migrenne, S.; Marsollier, N.; Cruciani-Guglielmacci, C.; Magnan, C. Importance of the Gut-Brain Axis in the Control of Glucose Homeostasis. Curr. Opin. Pharmacol. 2006, 6, 592–597. [Google Scholar] [CrossRef]
- Blouet, C.; Schwartz, G.J. Hypothalamic Nutrient Sensing in the Control of Energy Homeostasis. Behav. Brain Res. 2010, 209, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Migrenne, S.; Le Foll, C.; Levin, B.E.; Magnan, C. Brain Lipid Sensing and Nervous Control of Energy Balance. Diabetes Metab. 2011, 37, 83–88. [Google Scholar] [CrossRef] [PubMed]
- Andoh, T. Amino Acids Are More Important Insulinotropins than Glucose in a Teleost Fish, Barfin Flounder (Verasper Moseri). Gen. Comp. Endocrinol. 2007, 151, 308–317. [Google Scholar] [CrossRef]
- Navarro, I.; Rojas, P.; Capilla, E.; Albalat, A.; Castillo, J.; Montserrat, N.; Codina, M.; Gutiérrez, J. Insights into Insulin and Glucagon Responses in Fish. Fish Physiol. Biochem. 2002, 27, 205–216. [Google Scholar] [CrossRef]
- Gonzalez, R.; Reingold, B.K.; Gao, X.; Gaidhu, M.P.; Tsushima, R.G.; Unniappan, S. Nesfatin-1 Exerts a Direct, Glucose-Dependent Insulinotropic Action on Mouse Islet β- and MIN6 Cells. J. Endocrinol. 2011, 208, R9–R16. [Google Scholar] [CrossRef]
- Tagawa, N.; Ogura, H.; Miyawaki, H.; Asakawa, A.; Kato, I. Intraperitoneal Administration of Nesfatin-1 Stimulates Glucagon-like Peptide-1 Secretion in Fasted Mice. Mol. Med. Rep. 2023, 27, 1–9. [Google Scholar] [CrossRef]
- Ramesh, N.; Mortazavi, S.; Unniappan, S. Nesfatin-1 Stimulates Glucagon-like Peptide-1 and Glucose-Dependent Insulinotropic Polypeptide Secretion from STC-1 Cells in Vitro. Biochem. Biophys. Res. Commun. 2015, 462, 124–130. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Gao, L.; Tang, H.; Yin, Y.; Xiang, X.; Li, Y.; Zhao, J.; Mulholland, M.; Zhang, W. Peripheral Effects of Nesfatin-1 on Glucose Homeostasis. PLoS ONE 2013, 8, e71513. [Google Scholar] [CrossRef] [PubMed]
- Merz, K.E.; Thurmond, D.C. Role of Skeletal Muscle in Insulin Resistance and Glucose Uptake. Compr. Physiol. 2020, 10, 785–809. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Accession No. | Primer Sequence (5′-3′) | Annealing Temperature (°C) | |
|---|---|---|---|---|
| Forward | Reverse | |||
| 18S rRNA | MG830470.1 | GGATGCCCTTAACTGGGTGT | CTAGCGGCGCAATACGAATG | 60 |
| β-actin | LC382464.1 | CAGGGAGTGATGGTTGGCA | AACACGCAGCTCGTTGTAGA | 60 |
| ghra | XM_026270357.1 | CGCCAATGATTCCCAGACG | ATGGGCATGGTTGGGATTACA | 60 |
| ghrb | KT985189.1 | TCCACCAGTGATTCCCAGACG | GGTAGGCATTGCTGGGAGGT | 60 |
| igf1 | GU583648.1 | GGGGGCAGAAACTATCGCAT | GCACGTCCCTGCAAAAATTCA | 60 |
| igf2a | FJ410929.1 | CGTGCCGAAAAACTGTGGAA | CTCCGCACACGAACTGAAGA | 57 |
| slc2a1a | XM_026265785.1 | TGGCCTTCTTTGAGATTGGACC | ACTTTGAAGTAGGTGAAGACGAAGAA | 58 |
| slc2a2 | XM_026206696.1 | TGTGCTGTGGCCATGAC | CCAGGTCCGATCTCAAAGAA | 58 |
| slc5a1 | XM_026274578.1 | GATCGTGACCATGCCAGAG | TTTAGTCCCAGAGCCTGGTT | 58 |
| sglt 2 | XM_026264357.1 | GCACCTTGTTCACCATGGACAT | ACCACTCTGGGCTGCCTG | 58 |
| preproinsulin | LC387783.1 | ATCACGCCGAGCTGATAAGG | TGGTGAAGTCATTGGCGGTT | 57 |
| preproglucagon | U65528.1 | GCCTGGCTAAAATCCGGACA | CGTGATGAAGCAGTCAGCAG | 60 |
| pcsk1 | XM_026196557.1 | ATCGTGGTCATCCAGGTCAT | CGCACTTCTTTTGGTCCTGC | 58 |
| pcsk2 | NM_001142266.1 | GTCAGAAGCGAGGCTACAGA | CCATCATCCATAATGGCGATGGT | 58 |
| insra | AF218355.1 | GCCCACCTTGAAGGAGATGA | TTCCGAAGTCGCCAATCTTCA | 60 |
| insrb | AF321225.1 | GAGCTGCACCATTGCGTATC | CCACACGTAGGTCTTGCACA | 60 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rajeswari, J.J.; Unniappan, S. Tissue-Specific Modulation of Gluco- and Growth-Regulatory Factor Abundance by Nesfatin-1 and Nesfatin-1-like Peptide in Goldfish. Animals 2023, 13, 1437. https://doi.org/10.3390/ani13091437
Rajeswari JJ, Unniappan S. Tissue-Specific Modulation of Gluco- and Growth-Regulatory Factor Abundance by Nesfatin-1 and Nesfatin-1-like Peptide in Goldfish. Animals. 2023; 13(9):1437. https://doi.org/10.3390/ani13091437
Chicago/Turabian StyleRajeswari, Jithine Jayakumar, and Suraj Unniappan. 2023. "Tissue-Specific Modulation of Gluco- and Growth-Regulatory Factor Abundance by Nesfatin-1 and Nesfatin-1-like Peptide in Goldfish" Animals 13, no. 9: 1437. https://doi.org/10.3390/ani13091437
APA StyleRajeswari, J. J., & Unniappan, S. (2023). Tissue-Specific Modulation of Gluco- and Growth-Regulatory Factor Abundance by Nesfatin-1 and Nesfatin-1-like Peptide in Goldfish. Animals, 13(9), 1437. https://doi.org/10.3390/ani13091437