
Recombinant Antimicrobial Peptide OaBac5mini Alleviates Inflammation in Pullorum Disease Chicks by Modulating TLR4/MyD88/NF-κB Pathway
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Recombinant Expression of IFN_OaBac5mini
2.2. Real-Time Cell Assay
2.3. Total RNA Extraction and qRT-PCR
2.4. Animal Experiment Design
2.5. Organ Index Determination
2.6. Organ Bacterial Load Detection
2.7. Histopathology Examination
2.8. Western Blot Assays
2.9. Data statistical Analysis
3. Results
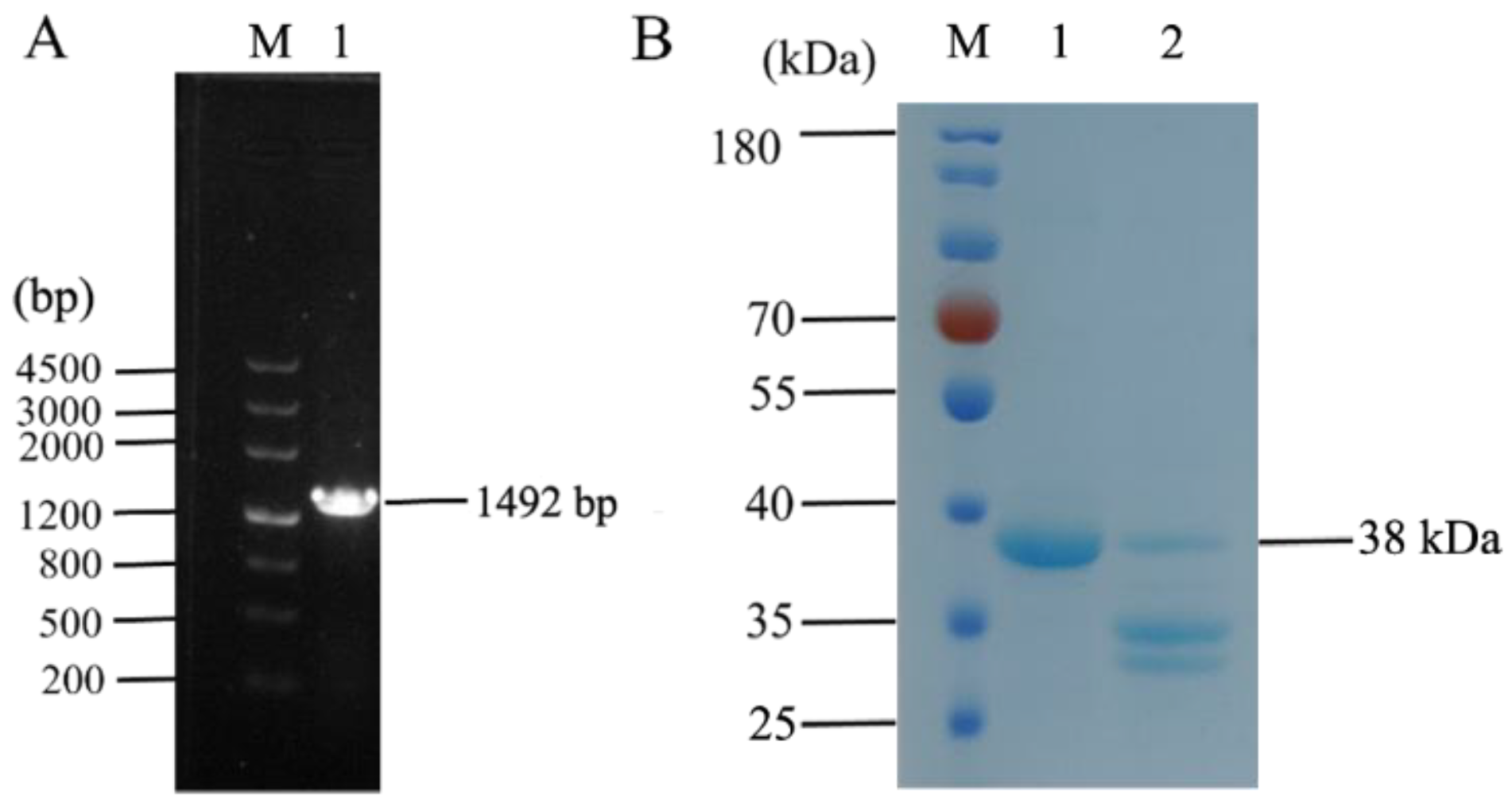
3.1. Recombinant Expression of Fusion Protein IFN_OaBac5mini
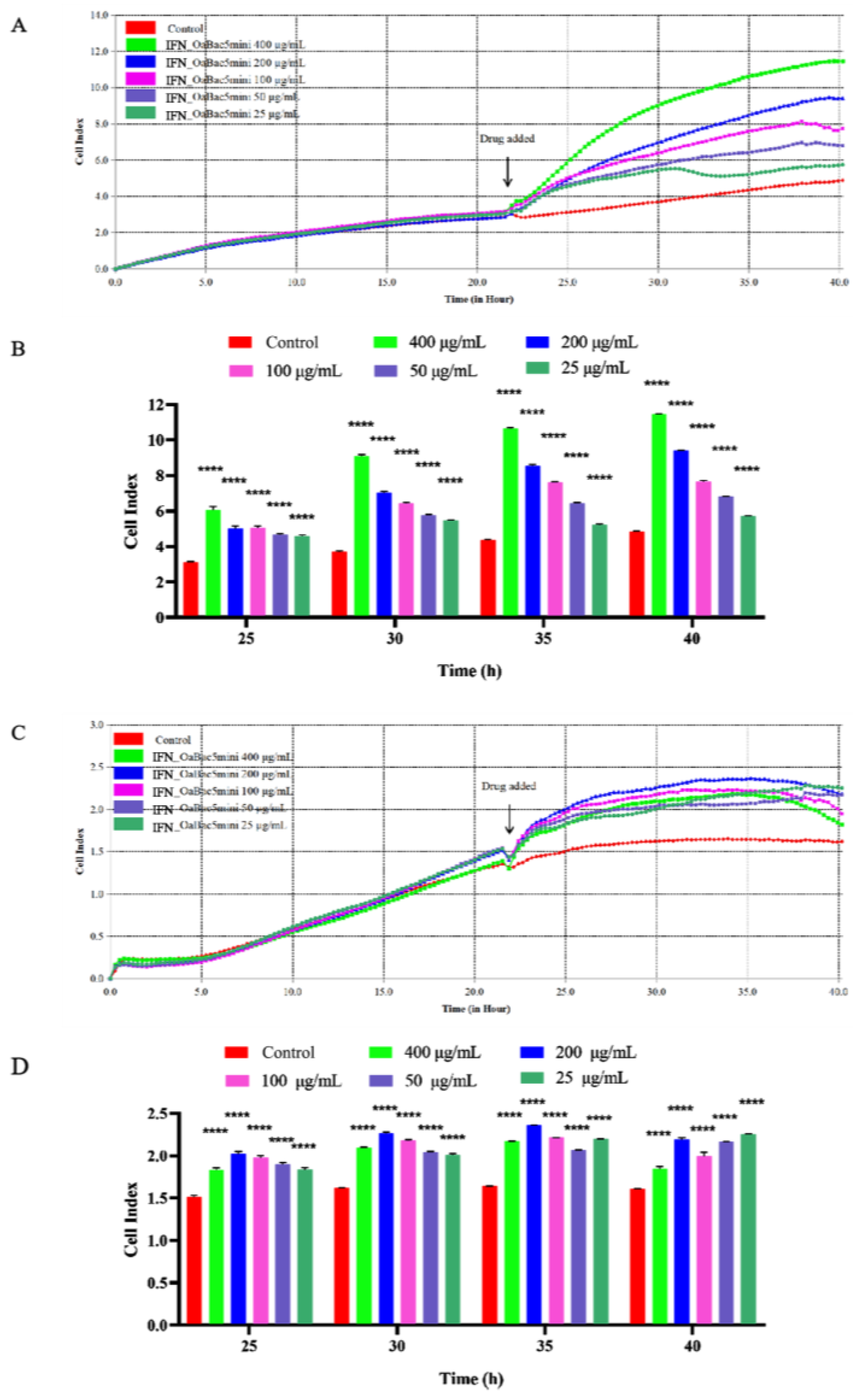
3.2. Recombinant IFN_OaBac5mini Exhibited no Obvious Cytotoxicity
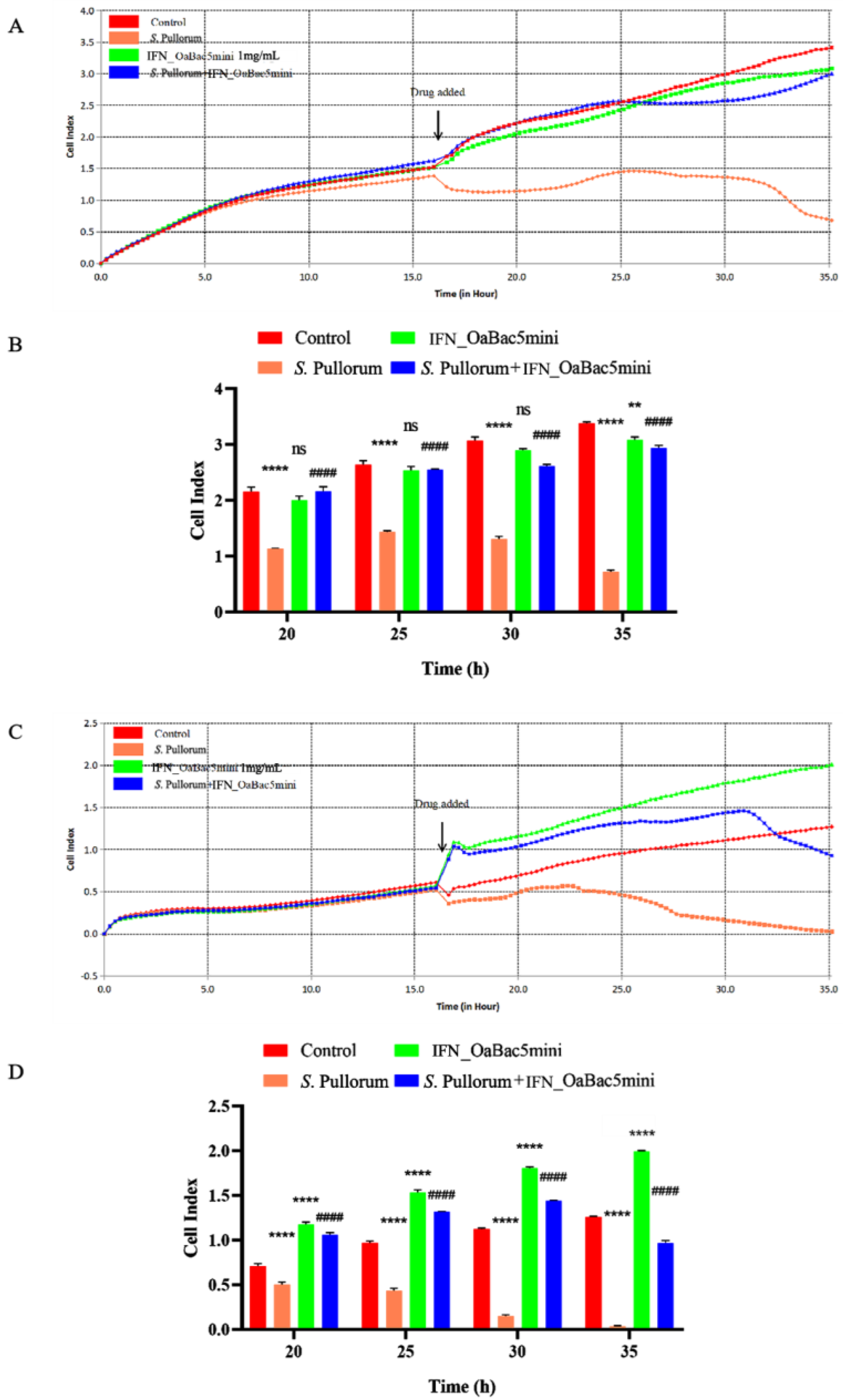
3.3. Recombinant OaBac5mini Alleviated the Drop in the Cell Index of S. Pullorum-Infected Cells
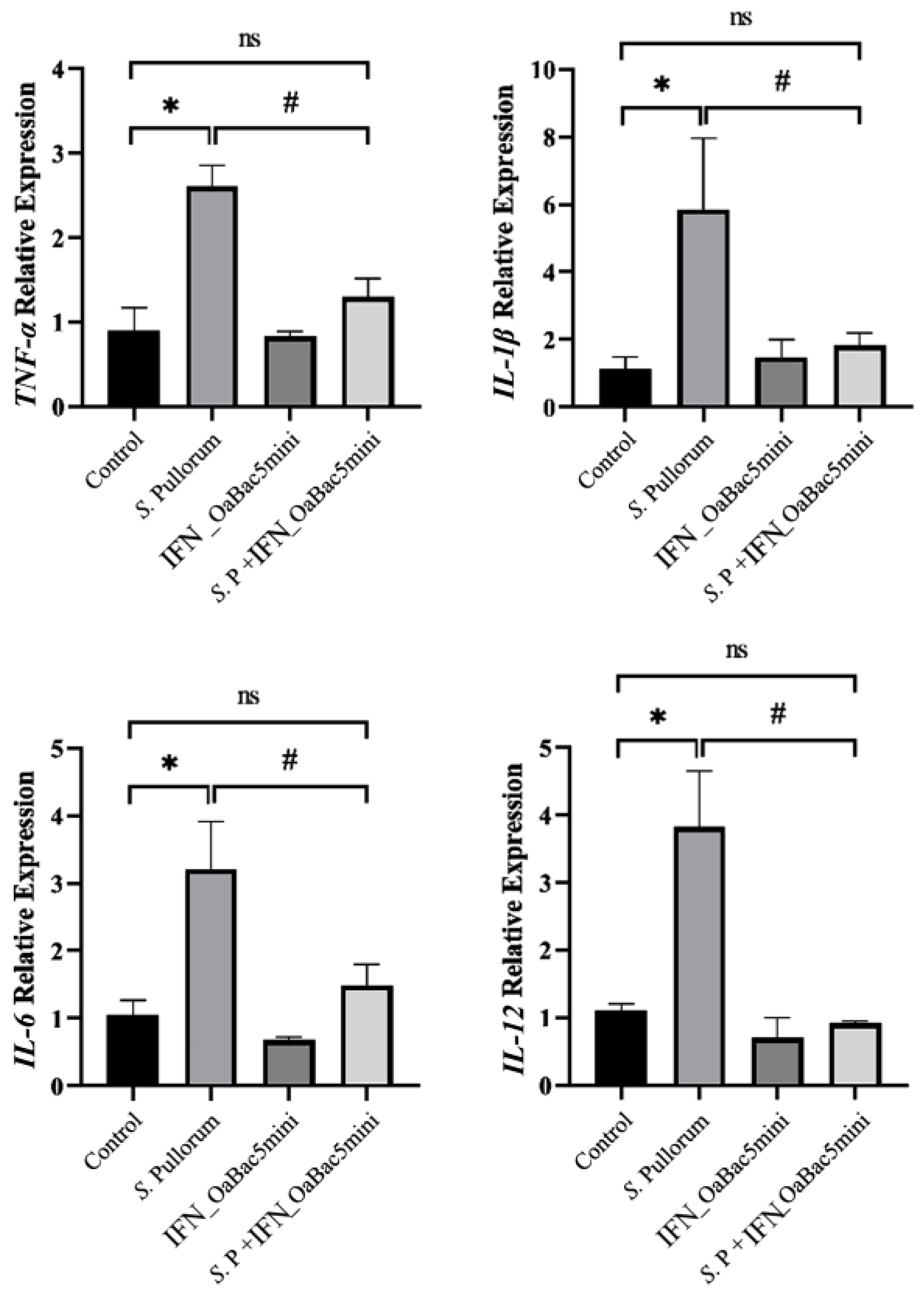
3.4. Recombinant OaBac5mini Suppressed the mRNA Expression of Pro-Inflammatory Cytokines Induced by S. Pullorum
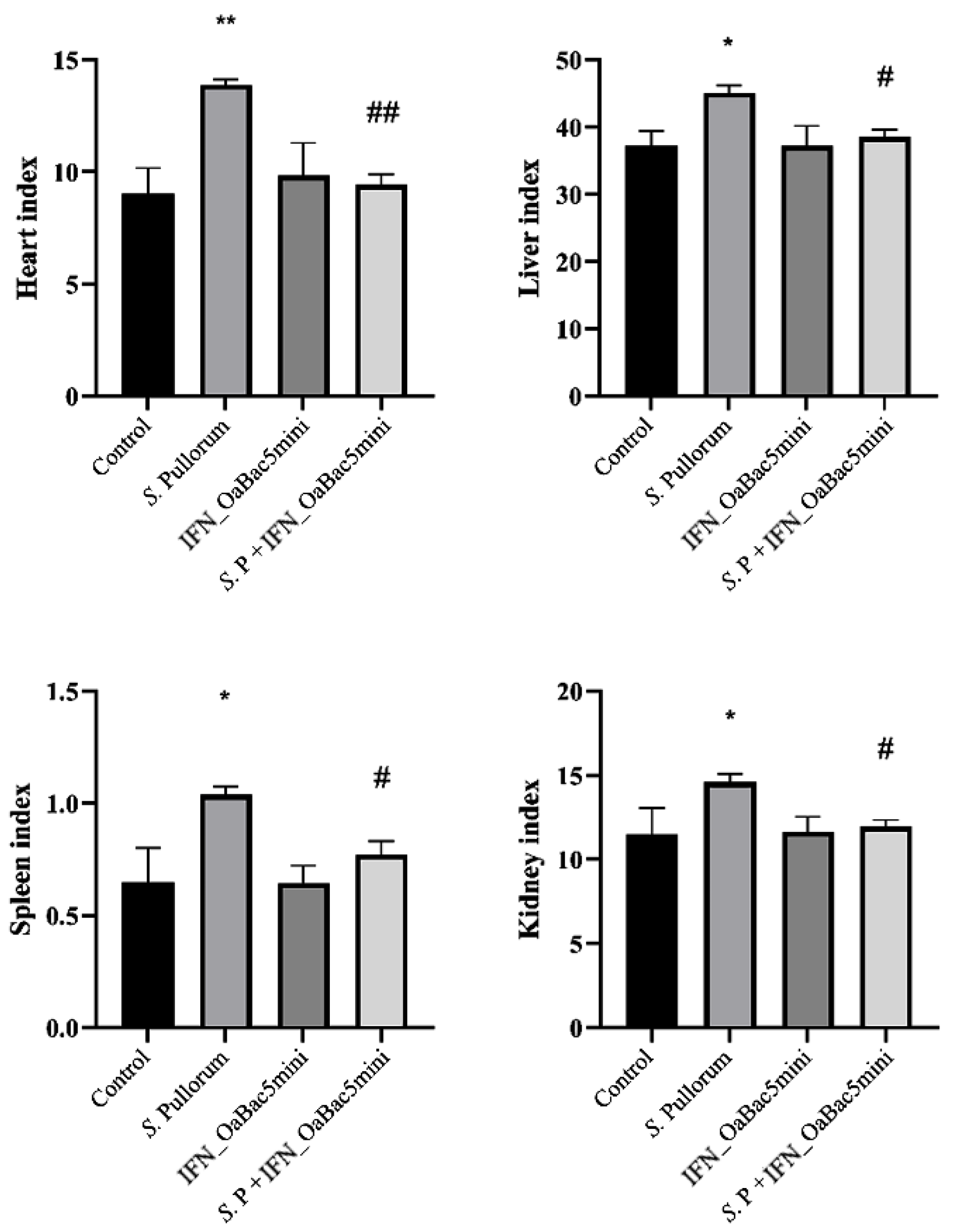
3.5. Recombinant OaBac5mini Attenuated the Increase in Organ Indexes in S. Pullorum-Challenged Chicks
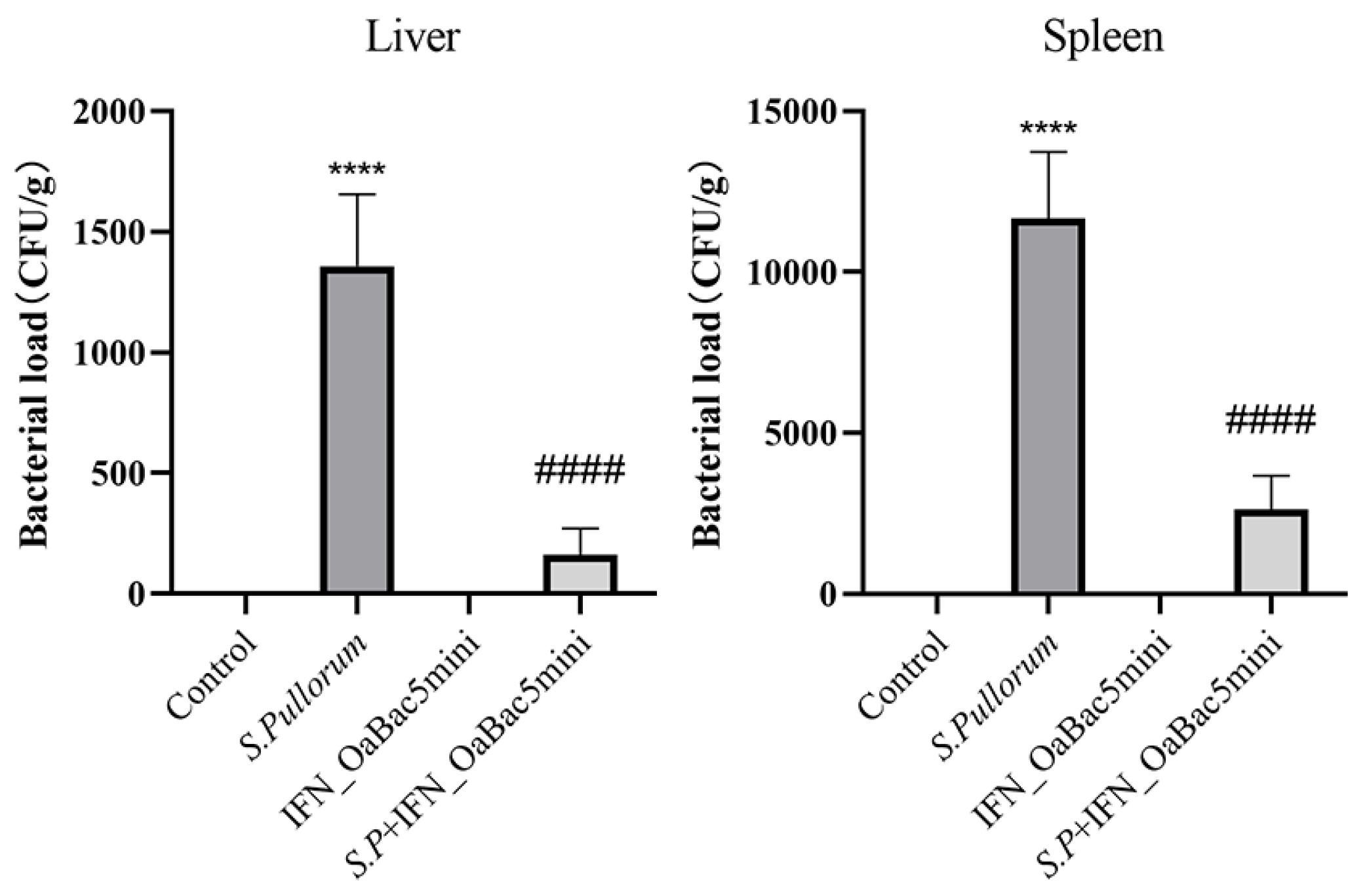
3.6. Recombinant OaBac5mini Decreased Organ Bacterial Loads in S. Pullorum-Challenged Chicks
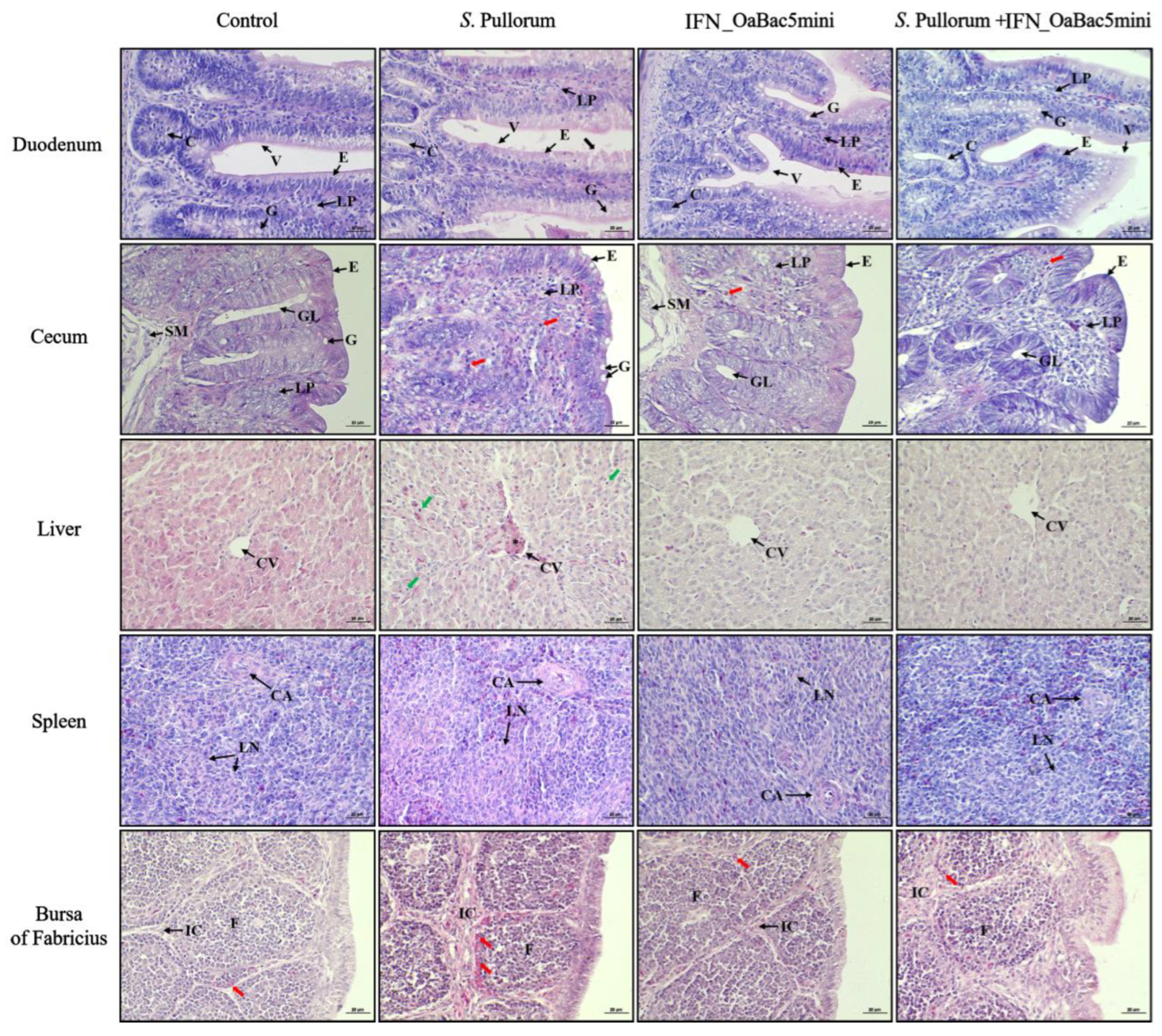
3.7. Recombinant OaBac5mini Ameliorated Histopathological Changes in PD Chicks
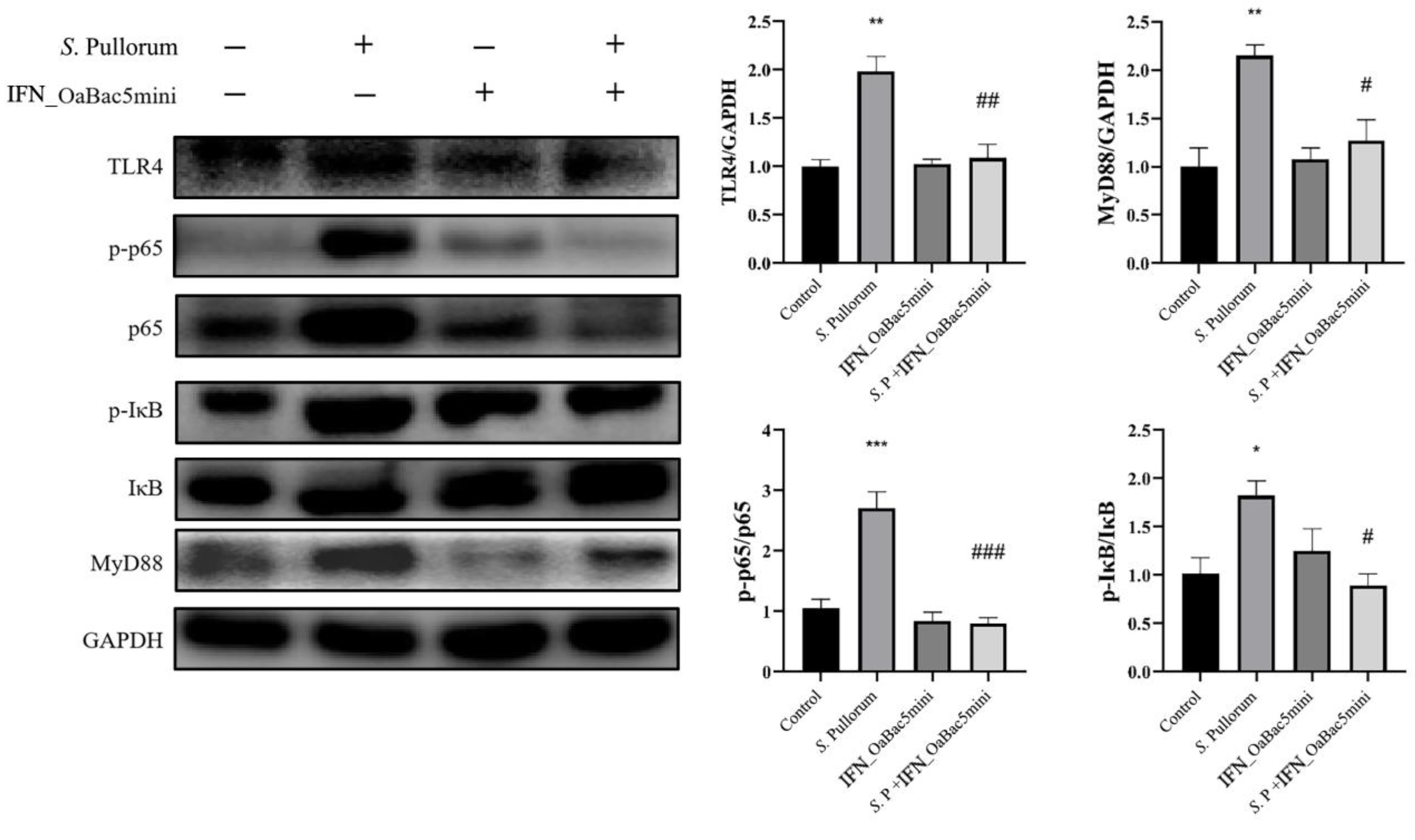
3.8. Recombinant OaBac5mini Modulated the TLR4/MyD88/NF-κB Pathway in PD Chicks
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hu, C.; Dou, W.; Zhao, G. Enzyme immunosensor based on gold nanoparticles electroposition and Streptavidin-biotin system for detection of S. pullorum & S. gallinarum. Electrochim. Acta 2014, 117, 239–245. [Google Scholar] [CrossRef]
- Zhou, X.; Kang, X.; Zhou, K.; Yue, M. A global dataset for prevalence of Salmonella gallinarum between 1945 and 2021. Sci. Data 2022, 9, 495. [Google Scholar] [CrossRef] [PubMed]
- Yin, J.; Cheng, Z.; Xu, Z.; Zhi, L.; Zhang, Y.; Yuan, X.; Pan, P.; Sun, W.; Yu, T.; Liu, T. Contribution of prgH gene for Salmonella Pullorum to virulence and the expression of NLRP3, Caspase-1 and IL-1β in chickens. Microb. Pathog. 2022, 171, 105744. [Google Scholar] [CrossRef]
- Cui, K.; Li, P.; Huang, J.; Lin, F.; Li, R.; Cao, D.; Hao, G.; Sun, S. Salmonella Phage CKT1 Effectively Controls the Vertical Transmission of Salmonella Pullorum in Adult Broiler Breeders. Biology 2023, 12, 312. [Google Scholar] [CrossRef] [PubMed]
- Tariq, S.; Samad, A.; Hamza, M.; Ahmer, A.; Muazzam, A.; Ahmad, S.; Amhabj, A. Salmonella in Poultry; An Overview. Int. J. Multidiscip. Sci. Arts 2022, 1, 80–84. [Google Scholar] [CrossRef]
- Gantois, I.; Ducatelle, R.; Pasmans, F.; Haesebrouck, F.; Gast, R.; Humphrey, T.J.; Van Immerseel, F. Mechanisms of egg contamination by Salmonella Enteritidis. FEMS Microbiol. Rev. 2009, 33, 718–738. [Google Scholar] [CrossRef]
- Ricke, S.C.; Atungulu, G.G.; Rainwater, C.E.; Park, S.H. Food and Feed Safety Systems and Analysis; Academic Press: San Diego, CA, USA, 2018; pp. 153–171. [Google Scholar] [CrossRef]
- Eng, S.K.; Pusparajah, P.; Ab Mutalib, N.S.; Ser, H.L.; Chan, K.G.; Lee, L.H. Salmonella: A review on pathogenesis, epidemiology and antibiotic resistance. Front. Life Sci. 2015, 8, 284–293. [Google Scholar] [CrossRef]
- Michael, G.B.; Schwarz, S. Antimicrobial resistance in zoonotic nontyphoidal Salmonella: An alarming trend? Clin. Microbiol. Infect. 2016, 22, 968–974. [Google Scholar] [CrossRef]
- AccessScience Editors. U.S. Bans Antibiotics Use for Enhancing Growth in Livestock. 2017. Available online: https://www.accessscience.com/content/briefing/aBR0125171 (accessed on 29 April 2023). [CrossRef]
- Tian, M.; He, X.; Feng, Y.; Wang, W.; Chen, H.; Gong, M.; Liu, D.; Clarke, J.L.; van Eerde, A. Pollution by Antibiotics and Antimicrobial Resistance in LiveStock and Poultry Manure in China, and Countermeasures. Antibiotics 2021, 10, 539. [Google Scholar] [CrossRef]
- Gan, B.H.; Gaynord, J.; Rowe, S.M.; Deingruber, T.; Spring, D.R. The multifaceted nature of antimicrobial peptides: Current synthetic chemistry approaches and future directions. Chem. Soc. Rev. 2021, 50, 7820–7880. [Google Scholar] [CrossRef]
- Mishra, A.K.; Choi, J.; Moon, E.; Baek, K.H. Tryptophan-Rich and Proline-Rich Antimicrobial Peptides. Molecules 2018, 23, 815. [Google Scholar] [CrossRef] [PubMed]
- Mardirossian, M.; Barrière, Q.; Timchenko, T.; Müller, C.; Pacor, S.; Mergaert, P.; Scocchi, M.; Wilson, D.N. Fragments of the Nonlytic Proline-Rich Antimicrobial Peptide Bac5 Kill Escherichia coli Cells by Inhibiting Protein Synthesis. Antimicrob. Agents Chemother. 2018, 62, e00534-18. [Google Scholar] [CrossRef] [PubMed]
- Anderson, R.C.; Yu, P.L. Isolation and characterisation of proline/arginine-rich cathelicidin peptides from ovine neutrophils. Biochem. Biophys. Res. Commun. 2003, 312, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Shen, S.; Sun, Y.; Ren, F.; Blair, J.M.A.; Siasat, P.; Fan, S.; Hu, J.; He, J. Characteristics of antimicrobial peptide OaBac5mini and its bactericidal mechanism against Escherichia coli. Front. Vet. Sci. 2023, 10, 1123054. [Google Scholar] [CrossRef]
- Anderson, R.C.; Hancock, R.E.; Yu, P.L. Antimicrobial activity and bacterial-membrane interaction of ovine-derived cathelicidins. Antimicrob. Agents Chemother. 2004, 48, 673–676. [Google Scholar] [CrossRef]
- Li, Y. Recombinant production of antimicrobial peptides in Escherichia coli: A review. Protein Expr. Purif. 2011, 80, 260–267. [Google Scholar] [CrossRef]
- Chen, P.; Jin, D.; Yang, S.; Yu, X.; Yi, G.; Hu, S.; Sun, Y.; Hu, Y.; Cui, J.; Rang, J.; et al. Aeromonas veronii infection remarkably increases expression of lysozymes in grass carp (Ctenopharyngodon idellus) and injection of lysozyme expression cassette along with QCDC adjuvant significantly upregulates immune factors and decreases cumulative mortality. Microb. Pathog. 2022, 169, 105646. [Google Scholar] [CrossRef]
- Martinez-Serra, J.; Gutierrez, A.; Muñoz-Capó, S.; Navarro-Palou, M.; Ros, T.; Amat, J.C.; Lopez, B.; Marcus, T.F.; Fueyo, L.; Suquia, A.G.; et al. xCELLigence system for real-time label-free monitoring of growth and viability of cell lines from hematological malignancies. Onco. Targets Ther. 2014, 7, 985–994. [Google Scholar] [CrossRef]
- Ngoc Le, H.T.; Kim, J.; Park, J.; Cho, S. A Review of Electrical Impedance Characterization of Cells for Label-Free and Real-Time Assays. Bio. Chip. J. 2019, 13, 295–305. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Hassan, M.; Flanagan, T.W.; Kharouf, N.; Bertsch, C.; Mancino, D.; Haikel, Y. Antimicrobial Proteins: Structure, Molecular Action, and Therapeutic Potential. Pharmaceutics 2022, 15, 72. [Google Scholar] [CrossRef] [PubMed]
- Hoelscher, M.P.; Forner, J.; Calderone, S.; Krämer, C.; Taylor, Z.; Loiacono, F.V.; Agrawal, S.; Karcher, D.; Moratti, F.; Kroop, X.; et al. Expression strategies for the efficient synthesis of antimicrobial peptides in plastids. Nat. Commun. 2022, 13, 5856. [Google Scholar] [CrossRef] [PubMed]
- Lv, Q.; Ran, X.; Qiu, H.; Zhao, S.; Hu, Z.; Wang, J.; Ni, H.; Wen, X. Seroprevalence of pullorum disease in chicken across mainland China from 1982 to 2020: A systematic review and meta-analysis. Res. Vet. Sci. 2022, 152, 156–166. [Google Scholar] [CrossRef]
- Wigley, P.; Berchieri, A., Jr.; Page, K.L.; Smith, A.L.; Barrow, P.A. Salmonella enterica serovar Pullorum persists in splenic macrophages and in the reproductive tract during persistent, disease-free carriage in chickens. Infect. Immun. 2001, 69, 7873–7879. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.G.; Wu, S.; Xia, Y.; Sun, J. Salmonella-infected crypt-derived intestinal organoid culture system for host-bacterial interactions. Physiol. Rep. 2014, 2, e12147. [Google Scholar] [CrossRef]
- Foster, N.; Tang, Y.; Berchieri, A.; Geng, S.; Jiao, X.; Barrow, P. Revisiting Persistent Salmonella Infection and the Carrier State: What Do We Know? Pathogens 2021, 10, 1299. [Google Scholar] [CrossRef]
- Setta, A.M.; Barrow, P.A.; Kaiser, P.; Jones, M.A. Early immune dynamics following infection with Salmonella enterica serovars Enteritidis, Infantis, Pullorum and Gallinarum: Cytokine and chemokine gene expression profile and cellular changes of chicken cecal tonsils. Comp. Immunol. Microbiol. Infect. Dis. 2012, 35, 397–410. [Google Scholar] [CrossRef]
- Duan, T.; Du, Y.; Xing, C.; Wang, H.Y.; Wang, R.F. Toll-Like Receptor Signaling and Its Role in Cell-Mediated Immunity. Front. Immunol. 2022, 13, 812774. [Google Scholar] [CrossRef]
- Yu, F.; Chen, J.; Lin, J.; Zhong, Z.; Lu, Y.; Zeng, X.; Lei, X. TLR4 involved in immune response against Vibrio Parahaemolyticus by MyD88-dependent pathway in Crassostrea hongkongensis. Fish Shellfish. Immunol. 2023, 134, 108591. [Google Scholar] [CrossRef]
- Zhang, Q.; Feng, A.; Zeng, M.; Zhang, B.; Shi, J.; Lv, Y.; Cao, B.; Zhao, C.; Wang, M.; Ding, Y.; et al. Chrysosplenol D protects mice against LPS-induced acute lung injury by inhibiting oxidative stress, inflammation, and apoptosis via TLR4-MAPKs/NF-κB signaling pathways. Innate Immun. 2021, 27, 514–524. [Google Scholar] [CrossRef]
- Li, P.; Xia, P.; Wen, J.; Zheng, M.; Chen, J.; Zhao, J.; Jiang, R.; Liu, R.; Zhao, G. Up-regulation of the MyD88-dependent pathway of TLR signaling in spleen and caecum of young chickens infected with Salmonella serovar Pullorum. Vet. Microbiol. 2010, 143, 346–351. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Lei, Y.; Chen, L.; Zhou, H.; Liu, H.; Jiang, J.; Yang, Y.; Wu, B. Phosphorylation of NF-κBp65 drives inflammation-mediated hepatocellular carcinogenesis and is a novel therapeutic target. J. Exp. Clin. Cancer Res. CR. 2021, 40, 253. [Google Scholar] [CrossRef] [PubMed]
- Veloso, P.; Fernández, A.; Astorga, J.; González-Quintanilla, D.; Castro, A.; Escobar, A.; Hoare, A.; Hernández, M. Lipopolysaccharide from Porphyromonas gingivalis, but Not from Porphyromonas endodontalis, Induces Macrophage M1 Profile. Int. J. Mol. Sci. 2022, 23, 10011. [Google Scholar] [CrossRef]
- Gong, T.; Fu, J.; Shi, L.; Chen, X.; Zong, X. Antimicrobial Peptides in Gut Health: A Review. Front. Nutr. 2021, 8, 751010. [Google Scholar] [CrossRef] [PubMed]
- Sun, Y.; Chan, J.; Bose, K.; Tam, C. Simultaneous control of infection and inflammation with keratin-derived antibacterial peptides targeting TLRs and co-receptors. Sci. Transl. Med. 2023, 15, eade2909. [Google Scholar] [CrossRef] [PubMed]
- Shin, M.K.; Lee, B.; Kim, S.T.; Yoo, J.S.; Sung, J.S. Designing a Novel Functional Peptide with Dual Antimicrobial and Anti-inflammatory Activities via in Silico Methods. Front. Immunol. 2022, 13, 821070. [Google Scholar] [CrossRef] [PubMed]
- Niyonsaba, F.; Iwabuchi, K.; Someya, A.; Hirata, M.; Matsuda, H.; Ogawa, H.; Nagaoka, I. A cathelicidin family of human antibacterial peptide LL-37 induces mast cell chemotaxis. Immunology 2002, 106, 20–26. [Google Scholar] [CrossRef] [PubMed]
- Mookherjee, N.; Brown, K.L.; Bowdish, D.M.; Doria, S.; Falsafi, R.; Hokamp, K.; Roche, F.M.; Mu, R.; Doho, G.H.; Pistolic, J.; et al. Modulation of the TLR-mediated inflammatory response by the endogenous human host defense peptide LL-37. J. Immunol. Baltim. Md. 1950 2006, 176, 2455–2464. [Google Scholar] [CrossRef] [PubMed]
- Kuroda, K.; Okumura, K.; Isogai, H.; Isogai, E. The Human Cathelicidin Antimicrobial Peptide LL-37 and Mimics are Potential Anticancer Drugs. Front. Oncol. 2015, 5, 144. [Google Scholar] [CrossRef]








Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shen, S.; Ren, F.; He, J.; Wang, J.; Sun, Y.; Hu, J. Recombinant Antimicrobial Peptide OaBac5mini Alleviates Inflammation in Pullorum Disease Chicks by Modulating TLR4/MyD88/NF-κB Pathway. Animals 2023, 13, 1515. https://doi.org/10.3390/ani13091515
Shen S, Ren F, He J, Wang J, Sun Y, Hu J. Recombinant Antimicrobial Peptide OaBac5mini Alleviates Inflammation in Pullorum Disease Chicks by Modulating TLR4/MyD88/NF-κB Pathway. Animals. 2023; 13(9):1515. https://doi.org/10.3390/ani13091515
Chicago/Turabian StyleShen, Shanshan, Fei Ren, Junping He, Jie Wang, Yawei Sun, and Jianhe Hu. 2023. "Recombinant Antimicrobial Peptide OaBac5mini Alleviates Inflammation in Pullorum Disease Chicks by Modulating TLR4/MyD88/NF-κB Pathway" Animals 13, no. 9: 1515. https://doi.org/10.3390/ani13091515
APA StyleShen, S., Ren, F., He, J., Wang, J., Sun, Y., & Hu, J. (2023). Recombinant Antimicrobial Peptide OaBac5mini Alleviates Inflammation in Pullorum Disease Chicks by Modulating TLR4/MyD88/NF-κB Pathway. Animals, 13(9), 1515. https://doi.org/10.3390/ani13091515