
Seasonal Variations on Testicular Morphology, Boar Taint, and Meat Quality Traits in Traditional Outdoor Pig Farming

 ,
,  ,
,  , ,
, ,  , ,
, ,  , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
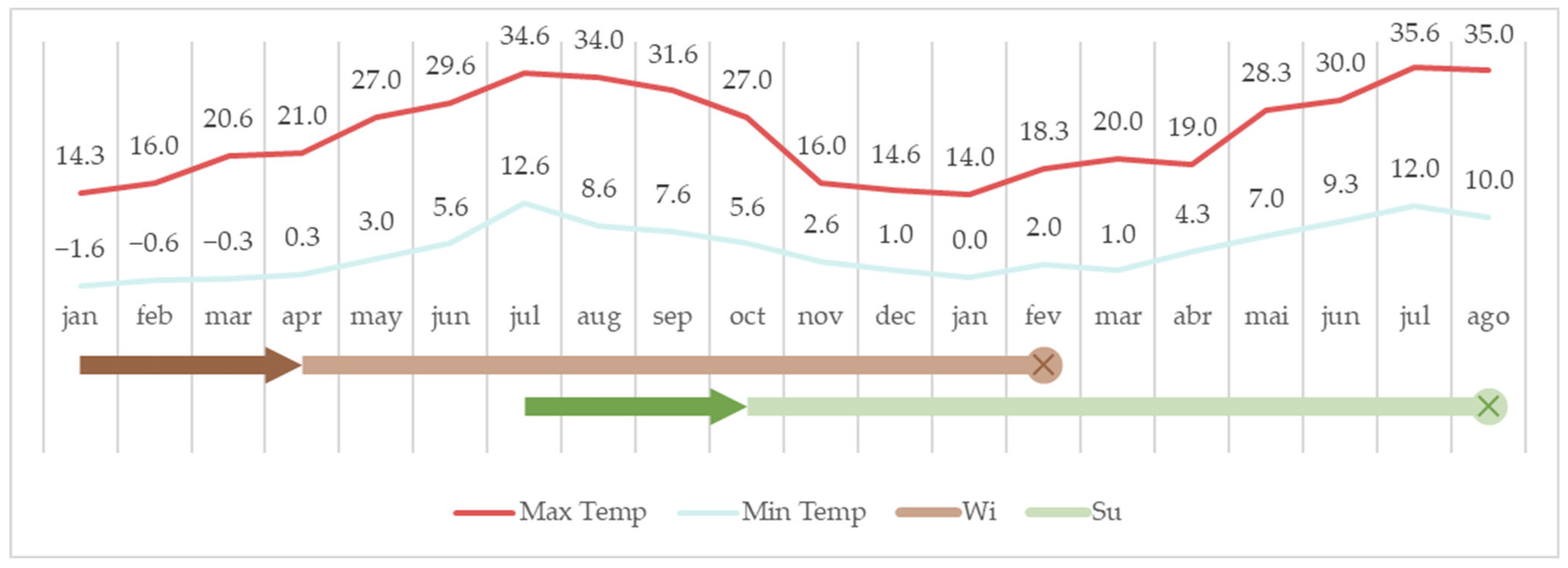
2.1. Animal Husbandry
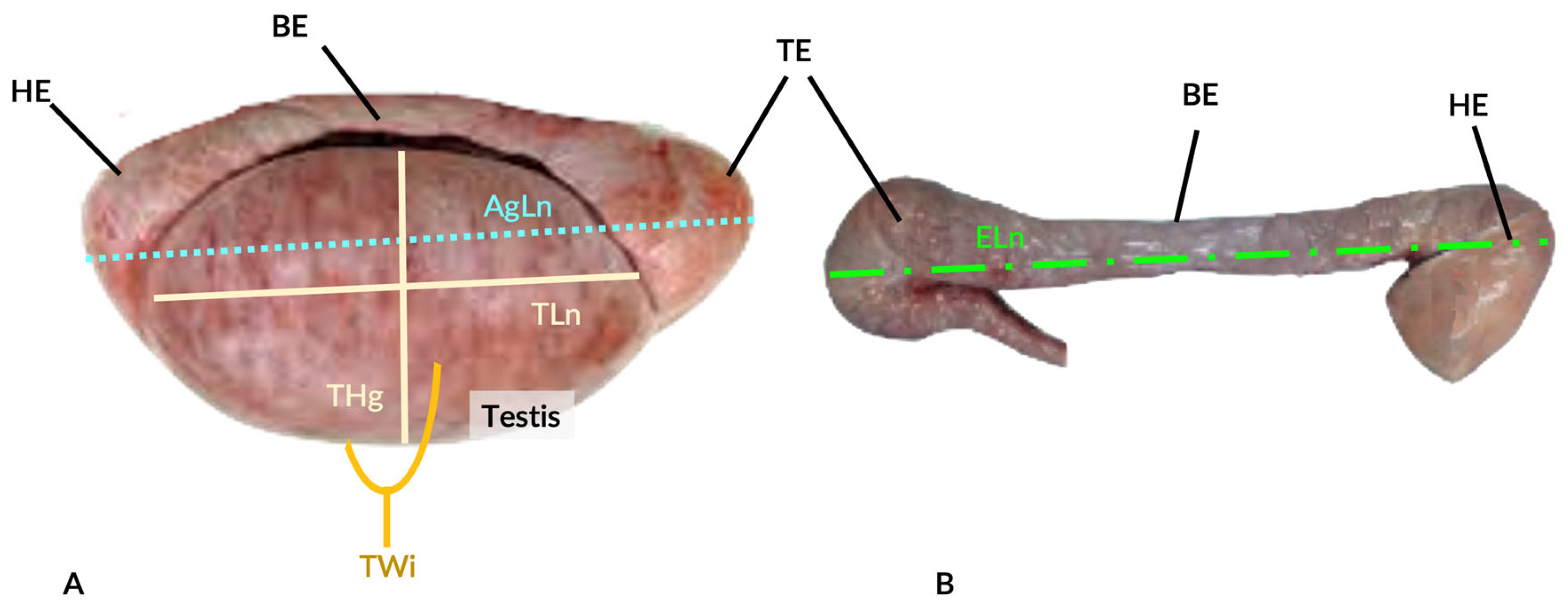
2.2. Reproductive Tract Morphometry
2.3. Physicochemical Analysis
2.4. Sensory Analysis
2.5. Fatty Acid Profile
2.6. Statistical Analysis
3. Results
3.1. Reproductive Tract Morphometry
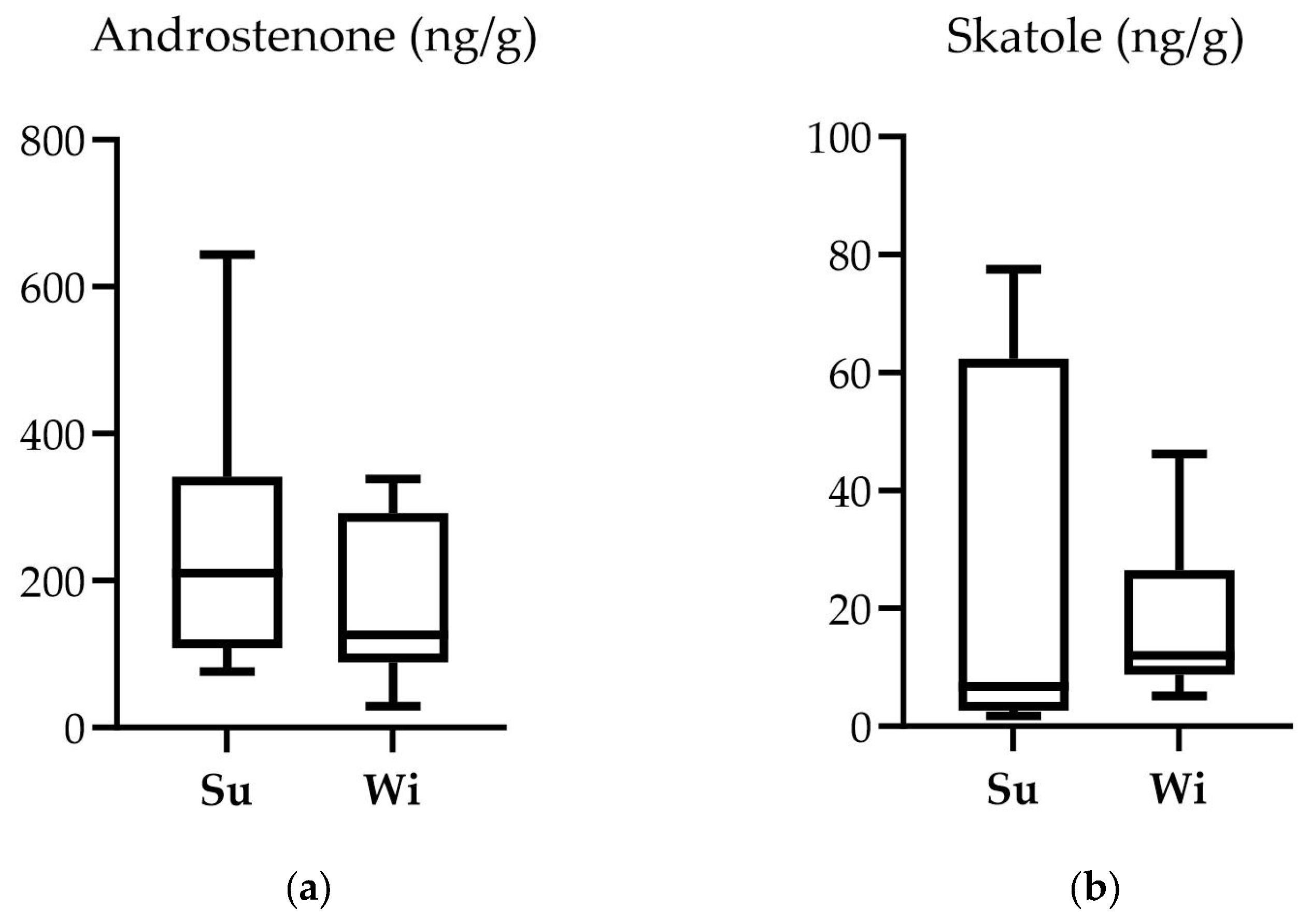
3.2. Physicochemical Analysis
3.3. Sensory Analysis
3.4. Fatty Acid Profile
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gade, P.B. Welfare of Animal Production in Intensive and Organic Systems with Special Reference to Danish Organic Pig Production. Meat Sci. 2002, 62, 353–358. [Google Scholar] [CrossRef] [PubMed]
- Dostálová, A.; Svitáková, A.; Bureš, D.; Vališ, L.; Volek, Z. Effect of an Outdoor Access System on the Growth Performance, Carcass Characteristics, and Longissimus Lumborum Muscle Meat Quality of the Prestice Black-Pied Pig Breed. Animals 2020, 10, 1244. [Google Scholar] [CrossRef] [PubMed]
- Alonso, M.E.; González-Montaña, J.R.; Lomillos, J.M. Consumers’ Concerns and Perceptions of Farm Animal Welfare. Animals 2020, 10, 385. [Google Scholar] [CrossRef] [PubMed]
- García-Gudiño, J.; Blanco-Penedo, I.; Gispert, M.; Brun, A.; Perea, J.; Font-i-Furnols, M. Understanding Consumers’ Perceptions towards Iberian Pig Production and Animal Welfare. Meat Sci. 2021, 172, 108317. [Google Scholar] [CrossRef] [PubMed]
- Szyndler-Nędza, M.; Światkiewicz, M.; Migdał, Ł.; Migdał, W. The Quality and Health-Promoting Value of Meat from Pigs of the Native Breed as the Effect of Extensive Feeding with Acorns. Animals 2021, 11, 789. [Google Scholar] [CrossRef] [PubMed]
- Sarmiento-García, A.; Vieira-Aller, C. Improving Fatty Acid Profile in Native Breed Pigs Using Dietary Strategies: A Review. Animals 2023, 13, 1696. [Google Scholar] [CrossRef]
- Čandek-Potokar, M.; Fontanesi, L.; Lebret, B.; Gil, J.M.; Ovilo, C.; Nieto, R.; Fernandez, A.; Pugliese, C.; Oliver, M.-A.; Bozzi, R. Introductory Chapter: Concept and Ambition of Project TREASURE. In European Local Pig Breeds-Diversity and Performance: A Study of Project TREASURE; IntechOpen: London, UK, 2019; pp. 3–9. ISBN 9781789854077. [Google Scholar]
- Edwards, S.A. Product Quality Attributes Associated with Outdoor Pig Production. Livest. Prod. Sci. 2005, 94, 5–14. [Google Scholar] [CrossRef]
- Nevrkla, P.; Kapelański, W.; Václavková, E.; Hadaš, Z.; Cebulska, A.; Horký, P. Meat Quality and Fatty Acid Profile of Pork and Backfat from an Indigenous Breed and a Commercial Hybrid of Pigs. Ann. Anim. Sci. 2017, 17, 1215–1227. [Google Scholar] [CrossRef]
- Leite, A.; Vasconcelos, L.; Ferreira, I.; Sarmiento-García, A.; Domínguez, R.; Santos, E.M.; Campagnol, P.C.B.; Rodrigues, S.; Lorenzo, J.M.; Teixeira, A. Chemical, Physicochemical and Sensorial Characterization of Nitrite-Free Dry-Cured Bísaro Shoulders. Foods 2022, 11, 3079. [Google Scholar] [CrossRef]
- Bedia, M.; Serrano, R.; Bañón, S. Use of Rustic Pigs to Manufacture Pork Salami Free of Colour and Taste Enhancers. Ital. J. Food Sci. 2012, 24, 223–230. [Google Scholar]
- Paixão, G.; Botelho-Fontela, S.; Marques, J.; Esteves, A.; Charneca, R.; Payan-Carreira, R. Long-Term Immunocastration Protocols Successfully Reduce Testicles’ Size in Bísaro Pigs. Animals 2021, 11, 632. [Google Scholar] [CrossRef] [PubMed]
- Rey, A.I.; Daza, A.; López-Carrasco, C.; López-Bote, C.J. Feeding Iberian Pigs with Acorns and Grass in Either Free-Range or Confinement Affects the Carcass Characteristics and Fatty Acids and Tocopherols Accumulation in Longissimus Dorsi Muscle and Backfat. Meat Sci. 2006, 73, 66–74. [Google Scholar] [CrossRef]
- Hälli, O.; Peltoniemi, O.A.T.; Tast, A.; Virolainen, J.V.; Munsterhjelm, C.; Valros, A.; Heinonen, M. Photoperiod and Luteinizing Hormone Secretion in Domestic and Wild Pigs. Anim. Reprod. Sci. 2008, 103, 99–106. [Google Scholar] [CrossRef] [PubMed]
- Peltoniemi, A.T.; Heinonenl, M.; Leppdvuori, A.; Low, R.J. Seasonal Effects on Reproduction in the Domestic Sow in Finland-A Herd Record Study. Acta Vet. Scand. 1999, 40, 133–144. [Google Scholar] [CrossRef] [PubMed]
- Peltoniemi, O.A.T.; Virolainen, J.V. Seasonality of Reproduction in Gilts and Sows. Reprod. Suppl. 2006, 62, 205–218. [Google Scholar] [CrossRef]
- Love, R.J.; Klupiec, C.; Thornton, E.J.; Evans, G. An Interaction between Feeding Rate and Season Affects Fertility of Sows. Anim. Reprod. Sci. 1995, 39, 275–284. [Google Scholar] [CrossRef]
- Claus, R.; Schopper, D.; Wagner, H.G. Seasonal Effect on Steroids in Blood Plasma and Seminal Plasma of Boars. J. Steroid Biochem. 1983, 19, 725–729. [Google Scholar] [CrossRef]
- Mauget, R.; Boissin, J. Seasonal Changes in Testis Weight and Testosterone Concentration in the European Wild Boar (Sus scrofa L.). Anim. Reprod. Sci. 1987, 13, 67–74. [Google Scholar] [CrossRef]
- Drimaj, J.; Kamler, J.; Rečková, Z.; Mikulka, O. Seasonal Variation in Testicular Biometry of Wild Boar in the Game Preserve. J. Vertebr. Biol. 2022, 71, 22059. [Google Scholar] [CrossRef]
- Decreto-Lei no. 1/2019, de 10 de Janeiro | DR. Available online: https://diariodarepublica.pt/dr/detalhe/decreto-lei/1-2019-117663336 (accessed on 19 July 2023).
- Diretiva 2010/63/UE Do Parlamento Europeu e Do Conselho de 22 de Setembro de 2010 Relativa à Protecção Dos Animais Utilizados Para Fins Científicos. Off. J. Eur. Union 2010, L276, 33–79.
- European Council Council Directive 93/119/EC on the Protection of Animals at the Time of Slaughter or Killing. Off. J. Eur. Union 1993, L340, 21–34.
- European Commission Council Regulation (EC) 1/2005 of 22 December 2004 on the Protection of Animals during Transport and Related Operations and Amending Directives 64/432/EEC and 93/119/EC and Regulation (EC) No 1255/97. Off. J. Eur. Union 2005, L3, 1–44.
- European Commission Regulation of the European Parliament and of the Council (29 April 2004). Laying down Specific Hygiene Rules for the Hygiene of Foodstuffs, 853/2004/EC. Off. J. Eur. Union 2004, L139, 55–206. [Google Scholar]
- Sakamoto, H.; Saito, K.; Oohta, M.; Inoue, K.; Ogawa, Y.; Yoshida, H. Adult Urology Testicular Volume Measurement: Comparison of Ultrasonography, Orchidometry, and Water Displacement. Urology 2007, 69, 152–157. [Google Scholar] [CrossRef] [PubMed]
- Drimaj, J.; Kamler, J.; Hošek, M.; Zeman, J.; Plhal, R.; Mikulka, O.; Kudláček, T.; Brno, I. Reproductive Characteristics of Wild Boar Males (Sus scrofa) under Different Environmental Conditions. Acta Vet. Brno 2019, 88, 401–412. [Google Scholar] [CrossRef]
- Botelho-Fontela, S.; Paixão, G.; Pereira-Pinto, R.; Vaz-Velho, M.; Dos, M.; Pires, A.; Payan-Carreira, R.; Patarata, L.; Lorenzo, J.; Silva, A.; et al. Effect of Immunocastration on Culled Sows—A Preliminary Study on Reproductive Tract, Carcass Traits, and Meat Quality. Vet. Sci. 2023, 10, 600. [Google Scholar] [CrossRef]
- Commission Internationale de l’Éclairage (CIE). Colorimetry, 2nd ed.; Commission Internationale de l’Éclairage: Vienna, Austria, 1986. [Google Scholar]
- Hornsey, H.C. The Colour of Cooked Cured Pork. II.—Estimation of the Nitric Oxide-Haem Pigments. J. Sci. Food Agric. 1956, 7, 534–540. [Google Scholar] [CrossRef]
- DeVore, D.P.; Solberg, M. Oxygen Uptake in Postrigor Bovine Muscle. J. Food Sci. 1974, 39, 22–28. [Google Scholar] [CrossRef]
- Honikel, K.O. Reference Methods for the Assessment of Physical Characteristics of Meat. Meat Sci. 1998, 49, 447–457. [Google Scholar] [CrossRef]
- Honikel, K.O. How to Measure the Water-Holding Capacity of Meat? Recommendation of Standardized Methods. In Evaluation and Control of Meat Quality in Pigs. Current Topics in Veterinary Medicine and Animal Science; Tarrant, P.V., Eikelenboom, G., Monin, G., Eds.; Springer: Dordrecht, The Netherlands, 1987; pp. 129–142. ISBN 978-94-009-3301-9. [Google Scholar]
- ISO 1442:1197; Determination of Moisture Content. (Reference Method). International Organization for Standardization: Geneva, Switzerland, 2002.
- ISO 937:1978; Determination of Total Nitrogen Content. (Reference Method). International Organization for Standardization: Geneva, Switzerland, 2002.
- ISO 1615/2002; Determination of Total Ashes. Reference Method. International Organization for Standardization: Geneva, Switzerland, 2002.
- Pinto, R.P.; Vaz-velho, M.; Mata, F.; Pires, P. A Modified High-Performance Liquid Chromatographic Method for Simultaneous Quantification of Skatole and Androstenone in Pig’s Backfat. In Proceedings of the XVI Food Chemistry Meeting (Portuguese Chemistry Society), Castelo Branco, Portugal, 23–26 October 2022; pp. 111–112. [Google Scholar]
- Botelho-Fontela, S.; Paixão, G.; Pereira-Pinto, R.; Vaz-Velho, M.; Pires, M.A.; Payan-Carreira, R.; Patarata, L.; Lorenzo, J.M.; Silva, A.; Esteves, A. The Effects of Different Immunocastration Protocols on Meat Quality Traits and Boar Taint Compounds in Male Bísaro Pigs. Theriogenology 2024, 214, 89–97. [Google Scholar] [CrossRef]
- ISO 8586:2012; Sensory Analysis—General Guidelines for the Selection, Training and Monitoring of Selected Assessors and Expert Sensory Assessors. International Organization for Standardization: Geneva, Switzerland, 2003.
- Garrido, M.D.; Egea, M.; Linares, M.B.; Martínez, B.; Viera, C.; Rubio, B.; Borrisser-Pairó, F. A Procedure for Sensory Detection of Androstenone in Meat and Meat Products from Entire Male Pigs: Development of a Panel Training. Meat Sci. 2016, 122, 60–67. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.A.; Cardoso, R.; Vieira, R.; Almeida, J.C.; Gomes, M.J.; Venâncio, C.; Patarata, L. The Effect of Weaning and Slaughter Age on the Physicochemical and Sensory Characteristics of Arouquesa Beef—A PDO Portuguese Meat. Foods 2022, 11, 2505. [Google Scholar] [CrossRef] [PubMed]
- Lunde, K.; Egelandsdal, B.; Choinski, J.; Mielnik, M.; Flåtten, A.; Kubberød, E. Marinating as a Technology to Shift Sensory Thresholds in Ready-to-Eat Entire Male Pork Meat. Meat Sci. 2008, 80, 1264–1272. [Google Scholar] [CrossRef] [PubMed]
- Domínguez, R.; Purriños, L.; Pateiro, M.; Campagnol, P.C.B.; Reyes, J.F.; Munekata, P.E.S.; Lorenzo, J.M. Fatty Acids. In Methods to Assess the Quality of Meat Products. Methods and Protocols in Food Science; Humana: New York, NY, USA, 2022; pp. 41–52. [Google Scholar] [CrossRef]
- Cittadini, A.; Munekata, P.E.S.; Pateiro, M.; Sarriés, M.V.; Domínguez, R.; Lorenzo, J.M. Physicochemical Composition and Nutritional Properties of Foal Burgers Enhanced with Healthy Oil Emulsion Hydrogels. Int. J. Food Sci. Technol. 2021, 56, 6182–6191. [Google Scholar] [CrossRef]
- Borg, K.E.; Lunstra, D.D.; Christenson, R.K. Semen Characteristics, Testicular Size, and Reproductive Hormone Concentrations in Mature Duroc, Meishan, Fengjing, and Minzhu Boars. Biol. Reprod. 1993, 49, 515–521. [Google Scholar] [CrossRef]
- Ausejo, R.; Martínez, J.M.; Soler-llorens, P.; Bolarín, A.; Tejedor, T.; Falceto, M.V. Seasonal Changes of Nuclear DNA Fragmentation in Boar Spermatozoa in Spain. Animals 2021, 11, 465. [Google Scholar] [CrossRef]
- Zhang, X.Z.; Liu, J.H.; Sheng, H.Q.; Wu, H.J.; Wu, Y.; Yao, K.S.; Lu, J.C.; Zhang, F. Bin Seasonal Variation in Semen Quality in China. Andrology 2013, 1, 639–643. [Google Scholar] [CrossRef]
- Petrocelli, H.; Batista, C.; Gosálvez, J. Seasonal Variation in Sperm Characteristics of Boars in Southern Uruguay. Rev. Bras. Zootec. 2015, 44, 1–7. [Google Scholar] [CrossRef]
- Sancho, S.; Pinart, E.; Briz, M.; Garcia-Gil, N.; Badia, E.; Bassols, J.; Kádár, E.; Pruneda, A.; Bussalleu, E.; Yeste, M.; et al. Semen Quality of Postpubertal Boars during Increasing and Decreasing Natural Photoperiods. Theriogenology 2004, 62, 1271–1282. [Google Scholar] [CrossRef]
- Zasiadczyk, L.; Fraser, L.; Kordan, W.; Wasilewska, K. Individual and Seasonal Variations in the Quality of Fractionated Boar Ejaculates. Theriogenology 2015, 83, 1287–1303. [Google Scholar] [CrossRef]
- Knecht, D.; Środoń, S.; Szulc, K.; Duziński, K. The Effect of Photoperiod on Selected Parameters of Boar Semen. Livest. Sci. 2013, 157, 364–371. [Google Scholar] [CrossRef]
- Sancho, S.; Rodríguez-Gil, J.E.; Pinart, E.; Briz, M.; Garcia-Gil, N.; Badia, E.; Bassols, J.; Pruneda, A.; Bussalleu, E.; Yeste, M.; et al. Effects of Exposing Boars to Different Artificial Light Regimens on Semen Plasma Markers and “in Vivo” Fertilizing Capacity. Theriogenology 2006, 65, 317–331. [Google Scholar] [CrossRef] [PubMed]
- Kunavongkrit, A.; Suriyasomboon, A.; Lundeheim, N.; Heard, T.W.; Einarsson, S. Management and Sperm Production of Boars under Differing Environmental Conditions. Theriogenology 2005, 63, 657–667. [Google Scholar] [CrossRef] [PubMed]
- Claus, R.; Weiler, U. Influence of Light and Photoperiodicity on Pig Prolificacy. J. Reprod. Fertil. Suppl. 1985, 33, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.; Robison, O.W.; Whisnant, C.S.; Cassady, J.P. Effect of Divergent Selection for Testosterone Production on Testicular Morphology and Daily Sperm Production in Boars. J. Anim. Sci. 2004, 82, 2259–2263. [Google Scholar] [CrossRef]
- Knecht, D.; Środoń, S.; Duziński, K. The Influence of Boar Breed and Season on Semen Parameters. S. Afr. J. Anim. Sci. 2014, 44, 1–9. [Google Scholar] [CrossRef]
- James, E.R.; Carrell, D.T.; Aston, K.I.; Jenkins, T.G.; Yeste, M.; Salas-Huetos, A. The Role of the Epididymis and the Contribution of Epididymosomes to Mammalian Reproduction. Int. J. Mol. Sci. 2020, 21, 5377. [Google Scholar] [CrossRef]
- Gentry, J.G.; McGlone, J.J.; Miller, M.F.; Blanton, J.R. Environmental Effects on Pig Performance, Meat Quality, and Muscle Characteristics. J. Anim. Sci. 2004, 82, 209–217. [Google Scholar] [CrossRef]
- Lebret, B. Effects of Feeding and Rearing Systems on Growth, Carcass Composition and Meat Quality in Pigs. Animal 2008, 2, 1548–1558. [Google Scholar] [CrossRef]
- Millet, S.; Moons, C.P.H.; Van Oeckel, M.J.; Janssens, G.P.J. Welfare, Performance and Meat Qualify of Fattening Pigs in Alternative Housing and Management Systems: A Review. J. Sci. Food Agric. 2005, 85, 709–719. [Google Scholar] [CrossRef]
- Lebret, B.; Čandek-Potokar, M. Review: Pork Quality Attributes from Farm to Fork. Part I. Carcass and Fresh Meat. Animal 2022, 16, 402. [Google Scholar] [CrossRef] [PubMed]
- Ludwiczak, A.; Kasprowicz-Potocka, M.; Zaworska-Zakrzewska, A.; Składanowska-Baryza, J.; Rodriguez-Estevez, V.; Sanz-Fernandez, S.; Diaz-Gaona, C.; Ferrari, P.; Pedersen, L.J.; Couto, M.Y.R.; et al. Husbandry Practices Associated with Extensification in European Pig Production and Their Effects on Pork Quality. Meat Sci. 2023, 206, 109339. [Google Scholar] [CrossRef] [PubMed]
- Lebret, B.; Meunier-Salaün, M.C.; Foury, A.; Mormède, P.; Dransfield, E.; Dourmad, J.Y. Influence of Rearing Conditions on Performance, Behavioral, and Physiological Responses of Pigs to Preslaughter Handling, Carcass Traits, and Meat Quality. J. Anim. Sci. 2006, 84, 2436–2447. [Google Scholar] [CrossRef] [PubMed]
- Čobanović, N.; Stajković, S.; Blagojević, B.; Betić, N.; Dimitrijević, M.; Vasilev, D.; Karabasil, N. The Effects of Season on Health, Welfare, and Carcass and Meat Quality of Slaughter Pigs. Int. J. Biometeorol. 2020, 64, 1899–1909. [Google Scholar] [CrossRef] [PubMed]
- Bee, G.; Guex, G.; Herzog, W. Free-Range Rearing of Pigs during the Winter: Adaptations in Muscle Fiber Characteristics and Effects on Adipose Tissue Composition and Meat Quality Traits. J. Anim. Sci. 2004, 82, 1206–1218. [Google Scholar] [CrossRef] [PubMed]
- Enfält, A.C.; Lundström, K.; Hansson, I.; Lundeheim, N.; Nyström, P.E. Effects of Outdoor Rearing and Sire Breed (Duroc or Yorkshire) on Carcass Composition and Sensory and Technological Meat Quality. Meat Sci. 1997, 45, 1–15. [Google Scholar] [CrossRef]
- Warriss, P.D. Meat Science: An Introductory Text; CABI Pub: Oxon, UK, 2000; ISBN 0-85199-424-5. [Google Scholar]
- Kyriazakis, I.; Whittemore, C.T. Whittemore’s Science and Practice of Pig Production; Blackwell Publishing: Hoboken, NJ, USA, 2006; ISBN 9781405124485. [Google Scholar]
- Čobanović, N.; Bošković, M.; Vasilev, D.; Dimitrijević, M.; Parunović, N.; Djordjević, J.; Karabasil, N. Effects of Various Pre-Slaughter Conditions on Pig Carcasses and Meat Quality in a Low-Input Slaughter Facility. S. Afr. J. Anim. Sci. 2016, 46, 380–390. [Google Scholar] [CrossRef]
- Correa, J.A.; Gonyou, H.W.; Torrey, S.; Widowski, T.; Bergeron, R.; Crowe, T.G.; Laforest, J.P.; Faucitano, L. Welfare and Carcass and Meat Quality of Pigs Being Transported for Two Hours Using Two Vehicle Types during Two Seasons of the Year. Can. J. Anim. Sci. 2013, 93, 43–55. [Google Scholar] [CrossRef]
- Scheeren, M.B.; Gonyou, H.W.; Brown, J.; Weschenfelder, A.V.; Faucitano, L. Effects of Transport Time and Location within Truck on Skin Bruises and Meat Quality of Market Weight Pigs in Two Seasons. Can. J. Anim. Sci. 2014, 94, 71–78. [Google Scholar] [CrossRef]
- Barlocco, N.; Vadell, A.; Ballesteros, F.; Galietta, G.; Cozzolino, D. Predicting Intramuscular Fat, Moisture and Warner-Bratzler Shear Force in Pork Muscle Using near Infrared Reflectance Spectroscopy. Anim. Sci. 2006, 82, 111–116. [Google Scholar] [CrossRef]
- Gentry, J.G.; McGlone, J.J.; Miller, M.F.; Blanton, J.R. Diverse Birth and Rearing Environment Effects on Pig Growth and Meat Quality. J. Anim. Sci. 2002, 80, 1707–1715. [Google Scholar] [CrossRef] [PubMed]
- Prunier, A.; Brillouët, A.; Merlot, E.; Meunier-Salaün, M.C.; Tallet, C. Influence of Housing and Season on Pubertal Development, Boar Taint Compounds and Skin Lesions of Male Pigs. Animal 2013, 7, 2035–2043. [Google Scholar] [CrossRef] [PubMed]
- Thomsen, R.; Edwards, S.A.; Jensen, B.B.; Rousing, T.; Sorensen, J.T. Weight and Season Affects Androstenone and Skatole Occurrence in Entire Male Pigs in Organic Pig Production. Animal 2015, 9, 1577–1586. [Google Scholar] [CrossRef] [PubMed]
- Walstra, P.; Claudi-Magnussen, C.; Chevillon, P.; Von Seth, G.; Diestre, A.; Matthews, K.R.; Homer, D.B.; Bonneau, M. An International Study on the Importance of Androstenone and Skatole for Boar Taint: Levels of Androstenone and Skatole by Country and Season. Livest. Prod. Sci. 1999, 62, 15–28. [Google Scholar] [CrossRef]
- Hansen, L.L.; Larsen, A.E.; Jensen, B.B.; Hansen-Møller, J.; Barton-Gade, P. Influence of Stocking Rate and Faeces Deposition in the Pen at Different Temperatures on Skatole Concentration (Boar Taint) in Subcutaneous Fat. Anim. Prod. 1994, 59, 99–110. [Google Scholar] [CrossRef]
- Grela, E.R.; Świątkiewicz, M.; Kowalczuk-Vasilev, E.; Florek, M.; Kosior-Korzecka, U.; Skałecki, P. An Attempt of Implementation of Immunocastration in Swine Production—Impact on Meat Physicochemical Quality and Boar Taint Compound Concentration in the Meat of Two Native Pig Breeds. Livest. Sci. 2020, 232, 103905. [Google Scholar] [CrossRef]
- Liu, X.; Trautmann, J.; Wigger, R.; Zhou, G.; Mörlein, D. Fatty Acid Composition and Its Association with Chemical and Sensory Analysis of Boar Taint. Food Chem. 2017, 231, 301–308. [Google Scholar] [CrossRef]
- Aldal, I.; Kristin, A.; Haugen, J.; Gr, A.; Fjetland, O.; Leif, J.; Eikaas, H. Levels of Androstenone and Skatole and the Occurrence of Boar Taint in Fat from Young Boars. Livest. Prod. Sci. 2005, 95, 121–129. [Google Scholar] [CrossRef]
- Aluwé, M.; Heyrman, E.; Theis, S.; Sieland, C.; Thurman, K.; Millet, S. Chicory Fructans in Pig Diet Reduce Skatole in Back Fat of Entire Male Pigs. Res. Vet. Sci. 2017, 115, 340–344. [Google Scholar] [CrossRef]
- Bonneau, M.; Kempster, A.J.; Claus, R.; Claudi-magnussen, C.; Diestre, A.; Tornberg, E.; Walstra, P.; Chevillon, P.; Weiler, U.; Cook, G.L. An International Study on the Importance of Androstenone and Skatole for Boar Taint: I. Presentation of the Programme and Measurement of Boar Taint Compounds with Different Analytical Procedures. Meat Sci. 2000, 54, 251–259. [Google Scholar] [CrossRef]
- Hansen, L.L.; Stolzenbach, S.; Jensen, J.A.; Henckel, P.; Hansen-Møller, J.; Syriopoulos, K.; Byrne, D.V. Effect of Feeding Fermentable Fibre-Rich Feedstuffs on Meat Quality with Emphasis on Chemical and Sensory Boar Taint in Entire Male and Female Pigs. Meat Sci. 2008, 80, 1165–1173. [Google Scholar] [CrossRef] [PubMed]
- Prusa, K.; Nederveld, H.; Runnels, P.L.; Li, R.; King, V.L.; Crane, J.P. Prevalence and Relationships of Sensory Taint, 5α-Androstenone and Skatole in Fat and Lean Tissue from the Loin (Longissimus Dorsi) of Barrows, Gilts, Sows, and Boars from Selected Abattoirs in the United States. Meat Sci. 2011, 88, 96–101. [Google Scholar] [CrossRef] [PubMed]
- Whittington, F.M.; Zammerini, D.; Nute, G.R.; Baker, A.; Hughes, S.I.; Wood, J.D. Comparison of Heating Methods and the Use of Different Tissues for Sensory Assessment of Abnormal Odours (Boar Taint) in Pig Meat. Meat Sci. 2011, 88, 249–255. [Google Scholar] [CrossRef] [PubMed]
- Pinto, R.P.; Mata, F.; Pires, P.; Barros, M.; Araújo, J.P.; Vaz-velho, M. The Use of Sugar Beet Pulp in Pig Diet to Control Skatole Analysed by HPLC The Use of Sugar Beet Pulp in Pig Diet to Control Skatole Analysed by HPLC. Sci. Agric. 2023, 80, 2–6. [Google Scholar] [CrossRef]
- Pereira Pinto, R.; Martin, L.; Ramos, A.; Conceição, M.A.; da Costa, R.P.R.; Vaz-Velho, M. Feeding and Housing Boars after Puberty without Castration allows for Good Performance and Low Boar Taint. J. Anim. Feed Sci. 2022, 31, 123–134. [Google Scholar] [CrossRef]
- James Squires, E.; Bone, C.; Cameron, J. Pork Production with Entire Males: Directions for Control of Boar Taint. Animals 2020, 10, 1665. [Google Scholar] [CrossRef]
- Bee, G.; Quiniou, N.; Maribo, H.; Zamaratskaia, G.; Lawlor, P.G. Strategies to Meet Nutritional Requirements and Reduce Boar Taint in Meat from Entire Male Pigs and Immunocastrates. Animals 2020, 10, 1950. [Google Scholar] [CrossRef]
- De Smet, S.; Vossen, E. Meat: The Balance between Nutrition and Health. A Review. Meat Sci. 2016, 120, 145–156. [Google Scholar] [CrossRef]
- Morales, J.; Baucells, M.D.; Pérez, J.F.; Mourot, J.; Gasa, J. Body Fat Content, Composition and Distribution in Landrace and Iberian Finishing Pigs given Ad Libitum Maize- and Acorn-Sorghum-Maize-Based Diets. Anim. Sci. 2003, 77, 215–224. [Google Scholar] [CrossRef]
- Rossi, R.; Pastorelli, G.; Cannata, S.; Corino, C. Recent Advances in the Use of Fatty Acids as Supplements in Pig Diets: A Review. Anim. Feed. Sci. Technol. 2010, 162, 1–11. [Google Scholar] [CrossRef]
- Belkova, J.; Rozkot, M.; Weisbauerova, E.; Truneckova, J. Influence of Feed Substitution with Acorns on Fattening Performance, Carcass Characteristics, and Meat Composition of Prestice Black-Pied Pigs Finished in a Conventional Indoor System. Cogent Food Agric. 2023, 9, 2220182. [Google Scholar] [CrossRef]
- Kušec, G.; Komlenić, M.; Gvozdanović, K.; Sili, V.; Krvavica, M.; Radišić, Ž.; Kušec, I.D. Carcass Composition and Physicochemical Characteristics of Meat from Pork Chains Based on Native and Hybrid Pigs. Processes 2022, 10, 370. [Google Scholar] [CrossRef]
- Kasprzyk, A.; Tyra, M.; Babicz, M. Fatty Acid Profile of Pork from a Local and a Commercial Breed. Arch. Anim. Breed. 2015, 58, 379–385. [Google Scholar] [CrossRef]
- Wood, J.D.; Richardson, R.I.; Nute, G.R.; Fisher, A.V.; Campo, M.M.; Kasapidou, E.; Sheard, P.R.; Enser, M. Effects of Fatty Acids on Meat Quality: A Review. Meat Sci. 2004, 66, 21–32. [Google Scholar] [CrossRef]
- Dugan, M.E.R.; Vahmani, P.; Turner, T.D.; Mapiye, C.; Juárez, M.; Prieto, N.; Beaulieu, A.D.; Zijlstra, R.T.; Patience, J.F.; Aalhus, J.L. Pork as a Source of Omega-3 (n-3) Fatty Acids. J. Clin. Med. 2015, 4, 1999–2011. [Google Scholar] [CrossRef]
- Pugliese, C.; Sirtori, F. Quality of Meat and Meat Products Produced from Southern European Pig Breeds. Meat Sci. 2012, 90, 511–518. [Google Scholar] [CrossRef]
- Cebulska, A.; Václavková, E.; Bocian, M.; Dybała, J.; Wiśniewska, J.; Kapelański, W. Quality and Dietary Value of Pork Meat of the Puławska and Złotnicka Spotted Breeds, and Commercial Fattening Pigs. Anim. Sci. 2018, 18, 281–291. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Su (n = 11) | Wi (n = 9) | SEM | p-Value |
|---|---|---|---|---|
| Aggregate | ||||
| Weight (g) | 829.27 | 834.00 | 27.47 | 0.824 |
| Length (cm) | 16.77 | 17.50 | 0.25 | 0.145 |
| Testicles | ||||
| Length (cm) | 12.36 | 12.86 | 0.21 | 0.331 |
| Width (cm) | 7.68 | 8.81 | 0.21 | 0.005 |
| Height (cm) | 6.50 | 6.67 | 0.11 | 0.462 |
| Weight (g) | 313.05 | 312.06 | 10.95 | 0.966 |
| Volume (cm3) | 444.07 | 538.90 | 22.35 | 0.031 |
| Epididymis | ||||
| Length (cm) | 20.11 | 21.19 | 0.30 | 0.072 |
| Width of the head (cm) | 4.14 | 5.06 | 0.15 | <0.001 |
| Width of the tail (cm) | 4.66 | 3.42 | 0.19 | <0.001 |
| Weight (g) | 101.59 | 104.94 | 3.66 | 0.661 |
| GSI | 0.48 | 0.54 | 0.02 | 0.152 |
| Parameter | Su (n = 11) | Wi (n = 9) | SEM | p-Value |
|---|---|---|---|---|
| Live weight (kg) | 171.35 | 153.71 | 3.57 | 0.010 |
| Carcass weight (kg) | 136.04 | 118.17 | 3.45 | 0.006 |
| Dressing (%) | 79.33 | 76.85 | 0.68 | 0.067 |
| pH45min | 6.15 | 6.41 | 0.06 | 0.029 |
| pH24h | 5.41 | 5.60 | 0.03 | <0.001 |
| L* | 51.41 | 47.86 | 0.95 | 0.800 |
| a* | 21.94 | 21.91 | 0.26 | 0.968 |
| b* | 6.25 | 6.43 | 0.27 | 0.757 |
| C* | 22.83 | 22.86 | 0.30 | 0.955 |
| h° | 15.89 | 16.22 | 0.55 | 0.772 |
| Heme (mg/g) | 1.60 | 1.71 | 0.05 | 0.241 |
| Drip loss (%) | 4.15 | 3.05 | 0.33 | 0.131 |
| Cooking loss (%) | 27.75 | 28.02 | 0.63 | 0.840 |
| Shear force (N/cm2) | 54.95 | 71.20 | 4.21 | 0.052 |
| Moisture (%) | 73.08 | 72.12 | 0.18 | 0.004 |
| Protein (%) | 22.79 | 23.44 | 0.12 | 0.004 |
| Intramuscular Fat (%) | 1.50 | 1.93 | 0.18 | 0.261 |
| Ashes (%) | 1.14 | 1.13 | 0.01 | 0.635 |
| Parameter | Su (n = 4) | Wi (n = 4) | SEM | p-Value |
|---|---|---|---|---|
| Aroma | ||||
| Pork Intensity | 5.4 | 5.4 | 0.2 | 0.865 |
| Boar taint | 4.9 | 3.3 | 0.3 | 0.029 |
| Urine | 3.4 | 2.6 | 0.3 | 0.091 |
| Feces | 2.8 | 2.2 | 0.3 | 0.343 |
| Flavor | ||||
| Pork Intensity | 5.2 | 5.4 | 0.1 | 0.366 |
| Boar taint | 3.7 | 2.7 | 0.2 | <0.001 |
| Urine | 2.2 | 2.2 | 0.2 | 0.916 |
| Feces | 1.5 | 1.5 | 0.1 | 0.958 |
| Juiciness | 4.8 | 5.2 | 0.2 | 0.449 |
| Tenderness | 5.2 | 5.4 | 0.3 | 0.771 |
| Chewiness | 5.1 | 5.4 | 0.2 | 0.548 |
| Fatty Acid (mg/100 g Pork) | Su (n = 11) | Wi (n = 9) | SEM | p-Value |
|---|---|---|---|---|
| Saturated fatty acid (SFA) | 708.03 | 903.71 | 72.23 | 0.063 |
| Lauric acid (C12:0) | 1.29 | 1.42 | 0.14 | 0.909 |
| Myristic acid (C14:0) | 23.27 | 27.49 | 2.48 | 0.790 |
| Pentadecylic acid (C15:0) | 0.55 | 0.50 | 0.05 | 0.849 |
| Palmitic acid (C16:0) | 439.69 | 582.43 | 46.08 | 0.037 |
| Heptadecanoic acid (C17:0) | 3.50 | 3.74 | 0.30 | 0.732 |
| Stearic acid (C18:0) | 233.02 | 279.79 | 22.76 | 0.239 |
| Arachidic acid (C20:0) | 3.56 | 4.74 | 0.40 | 0.102 |
| Monounsaturated fatty acid (MUFA) | 908.24 | 1232.13 | 102.31 | 0.037 |
| Palmitoleic acid (C16:1 n − 7) | 59.70 | 89.01 | 8.08 | 0.020 |
| Oleic acid (C18:1 n − 9) | 733.02 | 977.13 | 83.39 | 0.074 |
| Eicosenoic acid (C20:1 n − 9) | 12.70 | 15.42 | 1.18 | 0.305 |
| Polyunsaturated fatty acid (PUFA) | 291.85 | 354.23 | 14.00 | 0.022 |
| Linoleic acid (C18:2 n − 6) | 220.29 | 259.35 | 11.19 | 0.082 |
| Alpha-linolenic (C18:3 n − 3) | 5.20 | 6.15 | 0.44 | 0.210 |
| Dihomo-γ-linolenic acid (C20:3 n − 6) | 5.87 | 5.90 | 0.20 | 0.949 |
| Arachidonic acid (C20:4 n − 6) | 47.09 | 64.13 | 2.73 | <0.001 |
| Eicosatrienoic acid (C20:3 n − 3) | 1.14 | 1.27 | 0.07 | 0.342 |
| Eicosapentaenoic acid (C20:5 n − 3) | 0.78 | 1.55 | 0.10 | <0.001 |
| Docosapentaenoic acid (C22:5 n − 3) | 3.39 | 6.32 | 0.42 | <0.001 |
| Docosahexaenoic acid (C22:6 n − 3) | 0.15 | 0.84 | 0.12 | 0.004 |
| PUFA/SFA | 0.44 | 0.42 | 0.02 | 0.729 |
| Total n − 3 | 10.66 | 16.14 | 0.87 | <0.001 |
| Total n − 6 | 281.17 | 338.09 | 13.24 | 0.028 |
| n − 6/n − 3 | 27.26 | 21.03 | 1.09 | <0.001 |
| Index of atherogenicity | 0.44 | 0.43 | 0.01 | 0.458 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Botelho-Fontela, S.; Ferreira, S.; Paixão, G.; Pereira-Pinto, R.; Vaz-Velho, M.; Pires, M.d.A.; Payan-Carreira, R.; Patarata, L.; Lorenzo, J.M.; Silva, J.A.; et al. Seasonal Variations on Testicular Morphology, Boar Taint, and Meat Quality Traits in Traditional Outdoor Pig Farming. Animals 2024, 14, 102. https://doi.org/10.3390/ani14010102
Botelho-Fontela S, Ferreira S, Paixão G, Pereira-Pinto R, Vaz-Velho M, Pires MdA, Payan-Carreira R, Patarata L, Lorenzo JM, Silva JA, et al. Seasonal Variations on Testicular Morphology, Boar Taint, and Meat Quality Traits in Traditional Outdoor Pig Farming. Animals. 2024; 14(1):102. https://doi.org/10.3390/ani14010102
Chicago/Turabian StyleBotelho-Fontela, Sofia, Sílvia Ferreira, Gustavo Paixão, Ricardo Pereira-Pinto, Manuela Vaz-Velho, Maria dos Anjos Pires, Rita Payan-Carreira, Luís Patarata, José M. Lorenzo, José A. Silva, and et al. 2024. "Seasonal Variations on Testicular Morphology, Boar Taint, and Meat Quality Traits in Traditional Outdoor Pig Farming" Animals 14, no. 1: 102. https://doi.org/10.3390/ani14010102
APA StyleBotelho-Fontela, S., Ferreira, S., Paixão, G., Pereira-Pinto, R., Vaz-Velho, M., Pires, M. d. A., Payan-Carreira, R., Patarata, L., Lorenzo, J. M., Silva, J. A., & Esteves, A. (2024). Seasonal Variations on Testicular Morphology, Boar Taint, and Meat Quality Traits in Traditional Outdoor Pig Farming. Animals, 14(1), 102. https://doi.org/10.3390/ani14010102